不同坛紫菜丝状体生化性状的比较研究
2021-09-08陆彩萍骆其君陈娟娟王铁杆陈海敏
陆彩萍 杨 锐 骆其君 陈娟娟 张 鹏 王铁杆 陈海敏
(1. 宁波大学食品与药学学院, 宁波 315211; 2. 宁波大学海洋学院, 浙江省海洋生物工程重点实验室, 宁波 315211;3. 浙江省海洋水产养殖研究所, 温州 325005)
坛紫菜(Neoporphyra haitanensis)作为我国特有的一种暖温带大型藻类, 主要在浙江和福建等南方沿海地区进行栽培, 其年产量约占全国紫菜总产量的75%, 具有非常重要的经济作用[1]。然而, 与其他人工栽培海藻的情况类似, 目前坛紫菜也面临着抗病性差和种质退化等问题[2]。因此, 研究者致力于通过品种选育, 寻找性状更优良、适应性和抗病能力强的新品种。
坛紫菜生长经历丝状体和叶状体2个世代, 自由丝状体处于营养生长阶段, 在没有环境改变的情况下, 其生长状态和生化参数较为稳定, 并且培养简单, 扩增速度快, 常用作种质保存的材料[2,3]。同时, 丝状体也是重要的种质材料, 其生长特性与抗逆性能决定了紫菜丝状体育苗工作的成败与效益。因此, 在丝状体阶段通过室内条件的控制来开展品种/品系的生化指标的对比研究, 不仅能准确反映出这些参数的差异, 开展性状的关联, 还有利于缩短育种过程, 提高育种效率。
生化参数是体现生物性状的内在指标。光合色素通过影响藻类的光合效率, 影响到藻类的生长[4], 与高产特性密切相关; 红藻糖苷是红藻重要的渗透压调节物和碳同化前体物质[5], 在抗逆性能和同化作用中具有重要意义; 类菌胞素氨基酸(MAAs)在红藻中起到减少紫外辐射伤害的作用[6],与红藻糖苷一起关系到紫菜的耐高温和干出等逆境适应; 而激素则是藻类生长发育和抗病抗逆的关键调节物质[7]。这些物质是否可以作为种质评价的标准运用到育种选育评估当中, 值得深入探讨。
本研究对闵丰1号(MF1)、闵丰2号(MF2)、申福2号(SF2)、浙东1号(ZD1)和浙东2号(ZD2)几个品种/品系的坛紫菜自由丝状体的光合色素、光合生长参数及与抗逆、生长有关的几种关键代谢物进行对比分析, 以评估各品种/品系之间的生产性能与这些参数间的关系, 为紫菜新品种选育的生化评价体系的提出提供依据。
1 材料与方法
1.1 材料与试剂
坛紫菜丝状体ZD1和ZD2来自浙江省海洋生物工程实验室; SF2由上海海洋大学严兴洪教授赠送;MF1和MF2由集美大学谢潮添教授赠送; 激素标准品, 美国Sigma Aldrich公司。
以添加适量的Ⅲ号营养母液(990 nmol/L KNO3,57 nmol/L K2HPO4, 9 nmol/L FeSO4·7H2O, 54 nmol/L Na2EDTA, 1 nmol/L MnSO4·H2O)的灭菌海水作为自由丝状体的培养液, 在(20±0.5) ℃, 20 μmol/(m2·s)的光照强度, 25‰的盐度, 在光照周期12L﹕12D的条件下摇床培养(转速200 r/min)。待生长密度达到0.8 g/L时收集, 进行指标检测。
1.2 仪器与设备
全波长扫描多功能酶标仪, 上海ThermoFisher公司; 液相氧电极, 英国Hansatech公司; 冷冻干燥机(美国LABCONCO公司); TSQ Quantum Access液相色谱-三重四极杆质谱联用分析系统, 美国Thermo Fisher Scientific公司; Hypersil Gold C18色谱柱(100 mm×2.1 mm, 1.7 μm, 美国Thermo Fisher Scientific公司)。
1.3 不同品种/品系坛紫菜丝状体光合色素的检测
参照Wellburn等[8]的方法提取并测到不同品种/品系坛紫菜自由丝状体中的叶绿素a(Chl.a)和类胡萝卜素(Car)含量。以酶标仪测定样品浸提液在750、665、652、510和480 nm处的吸光值, 以100%甲醇作为空白对照。根据公式计算Chl.a和Car的含量:
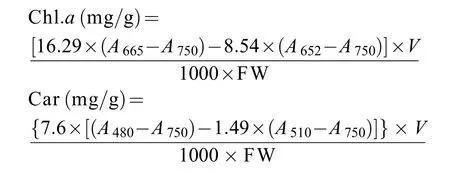
式中,A750、A665、A652、A510和A480表示样品在各波长下的吸光值,V为提取液的体积(mL), FW为丝状体鲜重(g)。
参照Hemlata等[9]的方法测定不同品种/品系坛紫菜自由丝状体中藻红蛋白(PE)和藻蓝蛋白(PC)的含量。测定样品上清液在455、564、592、618和645 nm处吸光值, 以磷酸缓冲液作为空白对照。PE和PC含量的计算公式如下:

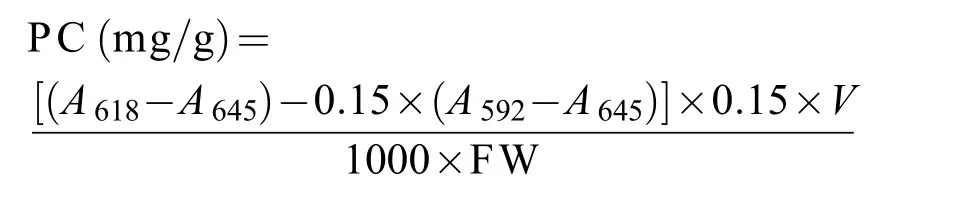
式中,A455、A564、A592、A618和A645表示样品在各波长下的吸光值。
1.4 坛紫菜丝状体净光合放氧速率检测
利用液相氧电极对样品进行净光合放氧速率的测定, LED灯提供光源, 光照强度为20 μmol/(m2·s)。称取丝状体鲜重约10 mg, 放入2 mL的新鲜培养液中, 培养温度20℃。方法见参考文献[10], 通过计算获得丝状体的净光合放氧速率。
1.5 坛紫菜丝状体生长曲线检测
将不同品种/品系丝状体放在100 mL锥形瓶中培养, 每隔5天将丝状体滤出, 称取丝状体的湿重, 记为Wt, 计算不同品种/品系坛紫菜丝状体的相对生长速率(Relative growth rate,RGR, %/d)。RGR的计算公式为:
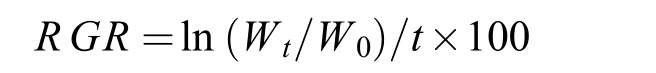
式中,W0表示初始重量(mg),t表示培养时间(d)。
1.6 坛紫菜丝状体红藻糖苷含量检测
取各品种/品系丝状体鲜重100 mg, 具体操作参照赖晓娟[11]的方法。根据不同浓度的红藻糖苷和异红藻糖苷绘制标准曲线, 计算含量。
1.7 坛紫菜丝状体MAAs含量检测
取各品种/品系丝状体冻干样品200 mg, 具体操作方法参照文献[12]。以MAAs标准品绘制标准曲线, 并计算各样品中MAAs含量。
1.8 坛紫菜丝状体激素含量检测
取各品种/品系丝状体冻干样品1 g, 液氮研磨,以10 mL甲醇﹕水﹕甲酸(V﹕V﹕V=15﹕4﹕1)溶液超声浸提10 min, 再于-20℃避光提取16h。上清液旋蒸并冻干, 以甲醇复溶, LC-MS检测参照宋悦等[10]的方法。根据不同浓度激素标准品绘制标准曲线, 并计算样品中植物激素含量。
1.9 数据分析
采用Pearson相关性分析各品种/品系丝状体间相对生长速率与光合色素、净光合放氧速率、红藻糖苷及激素的相关性。
每个实验设3个平行(n=3), 采用SPSS 23.0软件的最小显著性差异法进行单因素方差分析, 差异显著水平设置为P<0.05, 数据以平均值±标准差表示。
2 结果
2.1 不同品种/品系坛紫菜丝状体的光合色素
在不同品种/品系的坛紫菜丝状体中, 光合色素的含量皆以藻红蛋白和藻蓝蛋白为主(表 1)。对比5种丝状体发现, ZD2及MF1的藻红蛋白含量较为丰富, 均高于6 mg/g, 而ZD1和MF2中的含量则均低于4 mg/g。对比藻蓝蛋白含量, MF2中的含量仅为0.63 mg/g, 而其余4种丝状体中的藻蓝蛋白含量均高于1 mg/g, 且以MF1含量最高。在5种丝状体中, ZD2的叶绿素a含量最高, 但类胡萝卜素含量最低; ZD1和MF2中的叶绿素a含量较低但类胡萝卜素较高。

表1 不同品种/品系坛紫菜丝状体光合色素含量Tab. 1 The photosynthetic pigments contents of different varieties/strains of conchocelis of N. haitanensis (n=3)
2.2 不同品种/品系坛紫菜丝状体的净光合放氧速率
对坛紫菜丝状体中的净光合放氧速率进行检测, 发现不同品种/品系丝状体之间差异较大(图 1)。其中ZD2的净光合放氧速率相对其他品种/品系显著较高(P<0.05), 达到30.96 μmol O2/(g·h), ZD1和SF2相比ZD2显著较低(P<0.05), MF1和SF2之间差异不显著(P>0.05), 但MF2的净光合放氧速率和其余4种丝状体中相比, 显著较低(P<0.05), 只有23.07 μmol O2/(g·h)。由此可知, ZD1和ZD2尤其是ZD2的净光合放氧能力较强, MF2最弱。

图1 不同品种/品系坛紫菜丝状体净光合放氧速率测定Fig. 1 The net photosynthetic oxygen release rate in conchocelis of different varieties/strains of N. haitanensis
2.3 不同品种/品系坛紫菜丝状体的生长曲线
在最适生长条件下对5种坛紫菜丝状体进行培养, 不同时间取样检测其生长情况。图 2显示, ZD2和SF2的增长较为迅速, 相对生长速率分别达到0.93和0.81%/d, 最终重量是初始值的1.37倍和1.42倍。ZD2的相对生长速率显著高于其他几种(P<0.05),SF2、MF1及MF2丝状体之间差异不显著(P>0.05),ZD1相对生长速率为0.69%/d, 重量只是初始值的1.3倍, 为5种丝状体中最低。由此可知, 生长方面ZD2和SF2较占优势, ZD1的生长最为缓慢。

图2 不同品种/品系坛紫菜丝状体生长曲线及相对生长速率Fig. 2 Growth curves and relative growth rates of conchocelis of different varieties/strains of N. haitanensis
2.4 不同品种/品系坛紫菜丝状体中红藻糖苷含量差异
5个品种/品系丝状体的红藻糖苷含量差异较为明显(图 3)。在5个品种/品系中, ZD1和ZD2的总红藻糖苷含量较高, 分别为5.23 mg/g FW和5.82 mg/g。ZD1、ZD2和SF2中以异红藻糖苷含量为主, 且ZD2中的异红藻糖苷含量显著较高(P<0.05), 为5.38 mg/g含量极低。MF1和MF2中以红藻糖苷为主, 明显高于另外3种丝状体(P<0.05), 但异红藻糖苷含量极低。其中MF2的红藻糖苷显著高于MF1, 达到2.43 mg/g(P<0.05)。SF2丝状体红藻糖苷含量仅0.31 mg/g。

图3 不同品种/品系坛紫菜丝状体中红藻糖苷含量Fig. 3 The content of floridosides in conchocelis of different varieties/strains of N. haitanensis
2.5 不同品种/品系坛紫菜丝状体中MAAs含量差异
从图 4可知, MF2丝状体中Shinorine和Porphyra-334的含量显著高于另外4个品种/品系, 分别为4.27 μg/g DW(干重)和18.67 μg/g(P<0.05)。对比Shinorine含量, 可以看出ZD1和ZD2中含量显著低于其余3种(P<0.05), ZD2和SF2的差异不显著(P>0.05), MF1含量相对其余3种丝状体略高(P<0.05)。对比Porphyra-334含量, 只有MF2中的含量显著较高(P<0.05), 其余4种则无显著差异(P>0.05)。

图4 不同品种/品系坛紫菜自由丝状体MAAs含量Fig. 4 The content of MAAs of different varieties/strains of conchocelis of N. haitanensis
2.6 不同品种/品系坛紫菜丝状体的激素含量差异
在5种丝状体中只检测到异戊烯腺嘌呤、异戊烯腺苷、反式玉米素核苷和芸苔素内酯四种激素,且以反式玉米素核苷及芸苔素内酯含量最为丰富,其余2种激素含量均小于1 ng/g DW(表 2)。MF1和MF2中的反式玉米素核苷含量显著低于其余3种丝状体(P<0.05), 而ZD2中含量最丰富, 为662.52 ng/g。在5种丝状体中只有SF2的芸苔素内酯含量低于100 ng/g, 其次是ZD2和ZD1, 而MF1和MF2中的含量显著高于其余3种(P<0.05), 分别达到2325.09和6927.66 ng/g。可以看出, ZD和SF的坛紫菜丝状体含有的反式玉米素核苷较多, 芸苔素内酯较少, 并且ZD的异戊烯腺嘌呤含量相对较少, 而MF含有的主要是芸苔素内酯。
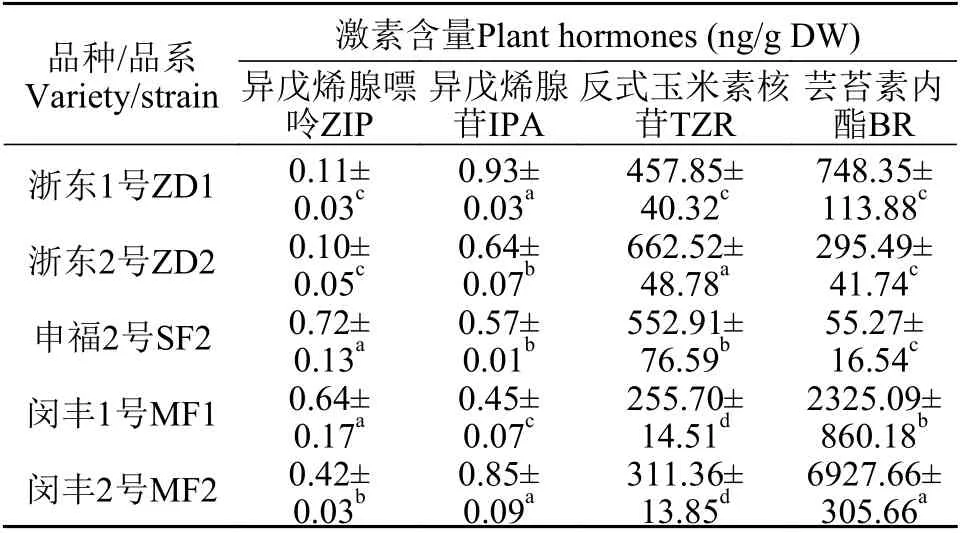
表2 不同品种/品系坛紫菜自由丝状体中植物激素含量Tab. 2 The content of plant hormones of different varieties/strains of conchocelis of N. haitanensis
2.7 不同品种/品系坛紫菜丝状体生理指标的相关性
对5个品种/品系丝状体部分生理指标进行相关性分析, 由图 5可见, 色素中的藻胆蛋白和叶绿素a与净光合放氧速率呈低至中度正相关(相关系数r为0.44—0.61,P<0.01), 藻红蛋白与相对生长速率呈中度正相关(r=0.66,P<0.01), 叶绿素a与相对生长速率高度正相关(r=0.98,P<0.01), 然而, 类胡萝卜素与光合放氧速率及相对生长速率不呈正相关关系(r=-0.41和-0.97,P<0.01); 此外, 坛紫菜丝状体的净光合放氧速率与相对生长速率也呈中度正相关(r=0.54,P<0.01)。红藻糖苷和异红藻糖苷与相对生长速率均无明显相关性(r=-0.16和0.26,P>0.05), 4种检出的激素中只有反式玉米素核苷与相对生长速率呈中度相关(r=0.55,P<0.01), 其余3种激素均无显著的正相关性。综上, 除了类胡萝卜素外的3种色素, 尤其是叶绿素a, 是影响相对生长速率的关键, 个别激素则存在一定的相关性, 而红藻糖苷与生长速率基本不存在相关性。
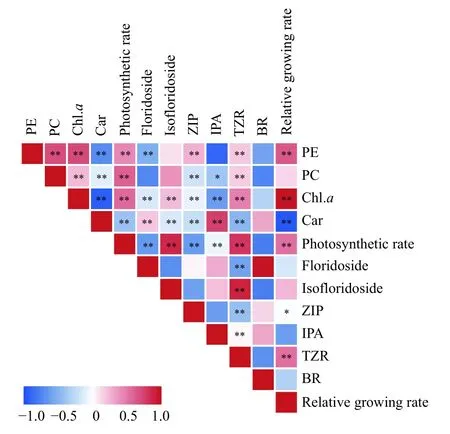
图5 坛紫菜丝状体不同品种/品系间生理指标相关性分析Fig. 5 Correlation analysis of physiological indexes among conchocelis of different varieties/strains of N. haitanensis
3 讨论
坛紫菜的种质评价多以产量、抗逆、表型及营养成分指标为主。比如SF2表现出明显的耐高温、生长快和成熟晚等特性; MF1和MF2具有耐高温和高产特性; 而ZD1则有叶片厚、产量高和壳孢子放散量高的特性等[13—15]。但这些指标偏重于生产性状的描述, 影响它们内在关联的生化指标并不明确。有文献指出, 光合色素与藻类的生长密切相关[16]; 红藻糖苷和MAAs是红藻耐高温和抗紫外的重要物质[17,18]; 而激素对植物的生长发育和抗逆关系密切[19,20]。因此, 这些指标是否可以用于表征品种/品系间的差异, 需要通过对性状明确的品种/品系进行比较来获取信息。
3.1 色素含量对坛紫菜生长的影响
坛紫菜生长及高产的特性与色素的含量和组成有关。藻胆蛋白是红藻中的重要捕光色素, 对短波长的蓝、绿光有较强吸收[21]。在本研究中, 坛紫菜丝状体中藻胆蛋白含量高于叶绿素a, 相关性分析显示出藻胆蛋白和叶绿素a与净光合放氧速率和相对生长速率显著相关, 而净光合速率也与相对生长速率正相关, 证实藻胆蛋白和叶绿素a的含量与丝状体的生长密切相关。这与钟晨辉等[4]和李映霞等[22]研究发现的丝状体光合效率受叶绿素a决定,并受藻胆蛋白影响的结论相符合。对比5个品种/品系丝状体中, ZD2、SF2和MF1的藻红蛋白含量均高于另外2个品种, 且叶绿素a在这3种丝状体中的含量也较高, 这有利于它们捕获光能用于光合作用。而ZD2、SF2和MF1相对其他2个品种的相对生长速率也更高。这表明, 同SF2和MF1这2个高产品种[1,14,15,23], 坛紫菜ZD2在丝状体阶段具有明显的生长优势。
3.2 红藻糖苷和MAAs在坛紫菜抵御逆境中的影响
红藻糖苷是大部分红藻的碳同化前体物质, 主要调节机体的渗透压、抗氧化及作为细胞壁多糖的合成前体起作用, 并且可以修复因高温受损的细胞壁, 红藻糖苷的合成和积累对坛紫菜应对高温胁迫产生的氧化损伤具有重要意义[5]。在紫菜中, 红藻糖苷由同分异构体红藻糖苷和异红藻糖苷组成,ZD1、ZD2和SF2坛紫菜丝状体中以异红藻糖苷居多, 而在MF1和MF2中以红藻糖苷含量居多。已有研究表明, 红藻糖苷及异红藻糖苷均能有效清除自由基, 且红藻糖苷的清除效率更高, 并且在遭受胁迫时红藻糖苷在代谢上比异红藻糖苷更为活跃[24],这使得坛紫菜在应对热胁迫时能更快作出响应来适应高温, MF品种含有更丰富的红藻糖苷, 说明其对热环境的耐受力更强。MAAs在红藻中起到吸收紫外线, 减少紫外辐射的作用[6]。在坛紫菜中以Porphyra-334和Shinorine两种形式为主。研究发现, 除了MF1是以Shinorine为主, 其余几种都以Porphyra-334为主, 而MF2的MAAs含量高于其他品种/品系。结合红藻糖苷的含量差异, 推测MAAS与红藻糖苷可能共同参与了坛紫菜的干出和高温的应激及耐受反应。而MF作为耐高温和抗紫外辐射的福建品种, 可能与这2个指标的高水平有关。
3.3 激素在坛紫菜生长发育中的影响
在坛紫菜丝状体中只检测到异戊烯腺嘌呤、异戊烯腺苷、反式玉米素核苷和芸苔素内酯4种植物激素, 而在叶状体中可测到吲哚乙酸、水杨酸、茉莉酸、茉莉酸甲酯、反式玉米素核苷、异戊烯腺苷和异戊烯腺苷7种植物激素[10]。异戊烯腺嘌呤、异戊烯腺苷和反式玉米素核苷都属于细胞分裂素, 在植物中是促进细胞分裂的激素, 参与植物的生长发育[25]。芸苔素内酯是植物内源生长激素,具有促进细胞的分裂和伸长, 提高植物的生长、产量等作用[26]。丝状体中缺乏水杨酸、茉莉酸等与抗逆、抗病害相关的激素, 说明丝状体的生长环境相对稳定, 主要需要调节生长和发育相关的激素。5个坛紫菜品种/品系的丝状体中4种激素含量各有差异, 反式玉米素核苷与生长有一定相关性, 在ZD2和SF2中含量较高, 而MF1和MF2的芸苔素内酯显著高于其他品种/品系, 由于几个品种/品系的选育都有高产的特性, 所以激素与生长的关系尚不明确。
4 总结
通过对比5个品种/品系的坛紫菜自由丝状体的光合色素、净光合放氧速率和相对生长速率, 以及几种关键的抗逆相关化合物, 发现光合色素中的藻红蛋白和叶绿素a与生长性状呈正相关性, 可以作为高产性状的评价指标, 其中含藻红蛋白丰富的品种/品系能更有效地捕捉深水中的蓝、绿光, 更适合深池培养; 而叶绿素a含量高的品种可以有效地捕捉红光, 适合浅池培养。红藻糖苷和MAAs与坛紫菜的耐高温和干出时的抗紫外能力有关, 因此这2种物质含量较高的品种/品系适合于较高温的南方水域或浅池培养。芸苔素内酯和反式玉米素核苷可能都具有调节生长的作用, 需进一步研究。总之, 本研究为新品种丝状体的优选和人工培育提出了选育生化指标的可行性。
