基于MaxEnt模型的菜豆象全球潜在适生区预测
2021-09-06徐养诚刘孝贤李东育高桂珍
徐养诚,刘孝贤,王 婷,李东育,高桂珍
1通标标准技术服务(青岛)有限公司,山东 青岛 266101; 2中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011; 3伊犁师范大学生物与地理科学学院, 新疆 伊犁 835000; 4新疆农业大学林学与园艺学院,新疆 乌鲁木齐 830052;
菜豆象Acanthoscelidesobtectus(Say)又名大豆象,属于鞘翅目豆象科三齿豆象属,以幼虫钻蛀籽粒的方式危害菜豆Phaseolusvulgaris(L.)、豇豆Vignaunguiculate(Linn.)Walp.、绿豆Vignaradiata(L.)Wilczek.、小豆Vignaumbellata(Thunb.)Ohwi et Ohashi、鹰嘴豆CicerarietinumL.、豌豆PisumsativumLinn.等40多种豆科作物,使其丧失商品价值,影响全球豆类贸易(刘永平和张生芳,1995;张润志,2017;Ursulaetal.,2009)。菜豆象具一定的飞翔能力,年自然扩散的平均距离可达25~30 km,具有中等扩散能力,主要以引种和运输被侵染的籽粒等方式传播,生态适应性强(莫仁浩和吴佳教,2006;吴长松等,2019;张润志,2017)。菜豆象原产中美洲和南美洲(张润志,2017),现已在美国、澳大利亚、南非、俄罗斯、日本、阿富汗、中国等几十个国家及地区传播(丁毅弘等,1993;刘永平和张生芳,1995;姚洁等,2017)。自1991年起,我国贵州、吉林、陕西、宁夏、云南等省区都有菜豆象疫情的报道,以2013年贵州省疫情最为严重,主要分布在7个地(市、州)的40个区、县(江兆春等,2018;齐贵林,1991;任轲亮等,2018;申智慧等,2014;赵龙等,2015)。经调查,菜豆象在贵州每年发生2~3代,世代重叠现象明显,仅2017年就有8个县(区)130多个乡镇的3.14万户农户和180多个经销商储存的豆类作物受害,一般受害率为2%~34%,最高受害率为100%(吴长松等,2019)。
生态位模型(ecological niche models, ENMs)是基于物种已知的地理分布数据和相关环境因素数据,采用一定算法推算物种的生态需求,并将结果投射至不同时间和空间来预测物种的实际与潜在分布(朱耿平等,2013)。常用的生态位模型中,最大熵模型MaxEnt具有操作简单、运行时间短、结果稳定和预测结果精度高等特点(李国庆等,2013;张路,2015),已被用于多种入侵性害虫的适生区预测,如马铃薯甲虫LeptinotarsadecemlineataSay(侯沁文等,2020)、稻水象甲LissorhoptrusoryzophilusKuschel(丁新华等,2019;齐国军等,2012)、草地贪夜蛾SpodopterafrugiperdaJ.E.Smith(秦誉嘉等,2019)、橙带蓝尺蛾MilioniabasalisPryeri(罗集丰等,2020)、红脂大小蠹DendroctonusvalensLeConte(崔骁芃等,2019)、蔗扁蛾OpogonasacchariBojer(王梦琳等,2017)、茶翅蝽HalyomorphapicusFabricius、美国白蛾HyphantriacuneaDrury(朱耿平等,2017,2018)等。
菜豆象是重要的入侵害虫,已有学者运用有害生物风险分析(pest risk analysis, PRA)程序对菜豆象在印度和中国的入侵风险进行了评估,结果表明,菜豆象的入侵风险高(江兆春等,2018;刘永平和张生芳,1995;任轲亮等,2018;汪成平等,2010;王菊平等,2006;Shashietal.,2016)。但目前鲜见采用生态位模型结合ArcGIS软件对其潜在分布区进行预测的相关报道,故本研究利用MaxEnt模型对菜豆象在全球的适生区进行预测,以期为农业生产部门和检疫部门对菜豆象的综合防控提供参考。
1 材料与方法
1.1 菜豆象的地理分布数据与处理
参考全球生物多样性信息网(http:∥www.gbif.org)、中国国家有害生物检疫信息系统(http:∥www.pestchina.com/pest/SitePages/PestInfoSearch.aspx)、中华人民共和国农业农村部2021年印发的《全国农业植物检疫性有害生物分布行政区名录》,结合相关参考文献(黄信飞等,1993;刘永平和张生芳,1995; 王成炬等,1993; 赵龙等,2015),全面收集菜豆象的地理分布数据,逐一核对、比较并删除重复发生点信息后得到415个发生点的经、纬度数据。按照物种名称、经度和纬度的次序输入Microsoft office excel 2010转换为CSV格式备用。
1.2 全球生物气候因素数据来源
生物气候因素数据是以全球的气象记录信息为基础数据,整合插值生成的全球气候栅格数据(http:∥www.worldclim.org),包含19个生物气候因素(11个温度相关因素和8个降雨相关因素)(表1)。数据的时间跨度为1970—2020年,分辨率为10 ARC-minutes,版本为WorldClim Version 2。

表1 生物气候因素信息表
1.3 地图数据来源
本研究使用的世界矢量地图从标准地图服务网站(http:∥bzdt.ch.mnr.gov.cn/index.html)获取。
1.4 MaxEnt模型软件及使用
MaxEnt软件从MaxEnt 主页(http:∥www.cs.princeton.edu/~schapire/MaxEnt/)下载,版本为MaxEnt Version 3.4.1。模拟过程中软件参数重复次数设置为10次,其余保持默认。
将菜豆象的地理分布数据和19个生物气候因素数据输入MaxEnt,筛选出影响菜豆象潜在分布的主要生物气候因素(雷军成等,2015;王茹琳等,2017;殷晓洁等,2013)。采用SPSS软件的Spearman相关系数分析模块计算其相关性,剔除相关性较强的生物气象因素,消除共线性对模型建模和结果的影响(王茹琳等,2017; Lemkeetal.,2011; Yangetal.,2013)。将剩余的生物气候因素输入MaxEnt模型并运行,输出连续格栅数据。将其导入ArcGIS转换为栅格图层,利用ArcGIS分析工具模块analysis tools再分类功能中的自然间断法选定合适的阈值进行分级,分为非适生区、低度适生区、中度适生区和高度适生区。
采用刀切法对生物气候因素贡献率进行分析,采用受试者工作特征曲线(receiver operating characteristic, ROC)线下面积值(area under curve, AUC)对模型的模拟结果进行评价。AUC值的取值范围为[0, 1],越接近1表明预测模型与生物气候因素之间的相关性越高,预测结果准确度就越高(Phillipsetal.,2006; Swets,1988)。
2 结果与分析
2.1 影响菜豆象分布的主要生物气候因素的贡献率及其相关性分析
采用MaxEnt模型筛选得出最冷月份最低温度、年平均温度、年气温变化范围、年降雨量、最干月份降雨量、最热月份最高温度、最暖季度平均温度、温度季节变化方差、昼夜温差月均值是影响菜豆象分布的主要生物气候因素,累计贡献率为96.3%(表2)。运用Spearman相关性分析后,剔除最热月份最高温度和最暖季度平均温度2个与年平均温度相关性较高的生物气象因素(表3)

表2 各生物气象因素贡献率和累计贡献率表

表3 生物气象因素之间的Spearman相关系数
2.2 模型预测精度评价
本研究基于菜豆象的415个全球分布点数据和7个生物气象数据的潜在适生区预测,应用ROC曲线分析法对MaxEnt模型预测结果可靠性进行检验。重复10次的AUC平均值为0.926(图1),大于0.9,表明此模型预测菜豆象的结果可靠。
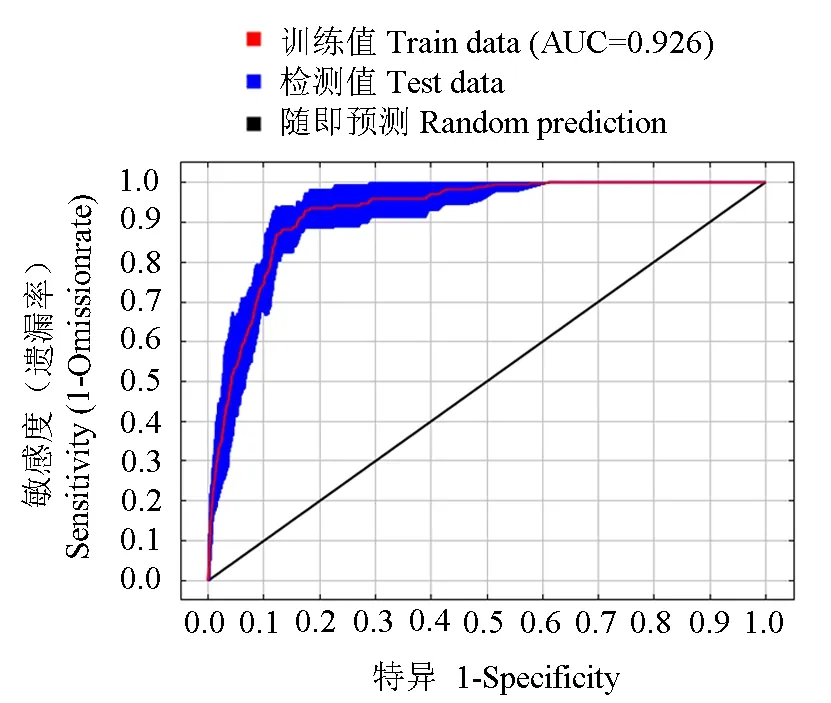
图1 菜豆象的潜在适生区模型的ROC曲线
2.3 菜豆象的潜在适生区
结果(图2)表明,菜豆象高度适生区占陆地面积的4.95%,主要分布在欧洲的德国、法国、荷兰、比利时、波兰西部地区、丹麦、瑞典南部地区,英国和爱尔兰和意大利大部分地区;南美洲的阿根廷东部、乌拉圭和巴西的东南部地区;非洲埃塞俄比亚中部、肯尼亚西南部、乌干达西南部、刚果东部、卢旺达和布隆迪、坦桑尼亚东北部、津巴布韦中部部分地区、南非的东部和南部沿海地区、马达加斯加岛中东部;大洋洲的澳大利亚东南部和西南部沿海地区、新西兰东部及南部地区;亚洲的中国云贵高原的云南北部、广西,云南和贵州交界地区。中度适生区占陆地面积的6.73%,主要分布在欧洲西部的西班牙北部、葡萄牙大部分地区、意大利北部、塞尔维亚、匈牙利、奥地利、波兰东部地区、罗马尼亚、保加利亚、希腊、立陶宛、拉脱维亚大部分地区、瑞典南部和白俄罗斯西部地区;南美洲的阿根廷东部、巴西东南部、巴拉圭南部、安迪斯山西麓;北美洲墨西哥北部和苏必利尔湖沿岸部分地区;亚洲中部的云贵高原大部分地区、四川盆地的部分地区和伊朗北部里海沿岸地区;非洲的南非东部部分地区、肯尼亚西南部地区、津巴布韦中东部地区、乌干达南部地区、埃塞俄比亚中西部地区、坦桑尼亚北部部分地区和马达加斯加中部部分地区;在大洋洲则以带状形式分布在澳大利亚高度适生区边缘。
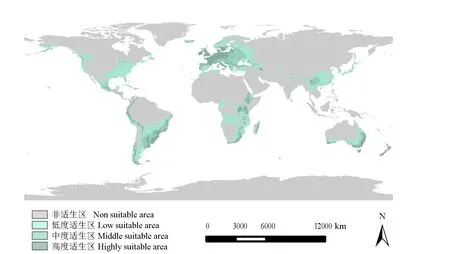
图2 菜豆象在全球潜在适生区的预测结果[审图号:GS(2021)3250号]
低度适生区占陆地面积的13.70%,主要分布在欧洲的西班牙大部分地区、乌克兰、俄罗斯西部地区、挪威、芬兰、瑞典中北部地区;南美洲的阿根廷东部地区、玻利维亚南部、巴拉圭大部分地区和巴西中部地区;北美洲主要分布在墨西哥中北部地区、美国中东部地区和西部沿海地区、加拿大西南部沿海地区;非洲的南非东部地区、马达加斯加南部、安哥拉大部分地区、喀麦隆大部分地区、坦桑尼亚和乌干达大部分地区、赞比亚、津巴布韦和莫桑比克大部分地区、刚果南部;在亚洲则主要分布在中国的华中和华南大部分地区和台湾岛、土耳其西部地区、里海沿岸地区、日本大部分地区、越南和老挝北部、缅甸东部地区、尼泊尔大部分地区、印度北部和南部的部分地区。
2.4 不同生物气候因素的贡献率
刀切法检验中,仅此变量得分越高,说明该因素的预测能力越高,其对物种分布的贡献就越大。由图3可知,本研究筛选的生物气候因素对菜豆象分布的贡献依次为年平均温度、年均温变化范围、最湿季度平均温度、年降雨量、温度季节性变化方差、最干月份降雨量、平均日较差。除此变量得分越低,说明该因素的独特性越高,对物种分布的重要性也越高。本文筛选的生物气候因素因素中,年平均温度、年降雨量和最干月份降雨量较为重要。
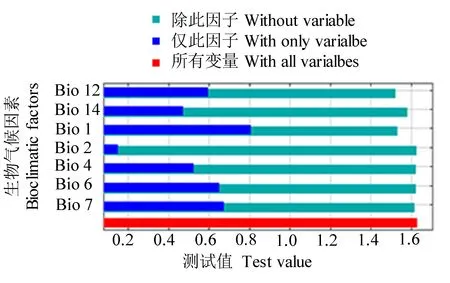
图3 主要生物气候因素正规则化训练增益的刀切法测试
3 讨论
本研究预测结果包含已报道存在菜豆象分布的国家或地区,如:欧洲中西部地区的英国、德国、法国、荷兰、西班牙、葡萄牙、意大利、波兰、乌克兰,美洲的墨西哥、智利、古巴、阿根廷、秘鲁,非洲的乌干达、肯尼亚、埃及、埃塞俄比亚、马达加斯加、南非,大洋洲的澳大利亚、新西兰和斐济,亚洲的伊朗、伊拉克、土耳其、印度、缅甸、日本等(黄信飞等,1993;莫仁浩等,2006;张生芳和刘永平,1992;刘永平和张生芳,1993,1995;王成炬等,1993;汪成平等,2010)。这些国家或地区的大部分区域均呈现出年平均温度较高、年降雨量大、湿度大,极端温度发生较少的气候特征。研究表明,菜豆象在相对湿度85%,温度22 ℃时卵的孵化率与幼虫的蛀入率较高,分别可达91%和95.6 ℃(李南植等,1995)。在相同湿度(湿度70%)条件下,当温度为19 ℃左右时,菜豆象成虫的平均寿命为30 d左右,最高可达43 d;而当温度为26 ℃左右时,成虫的平均寿命为13 d左右,最高18 d。相同温度(26.25 ℃)条件下,当湿度为70%时,菜豆象成虫平均产卵量为46粒,最高88粒;当湿度为80%时,成虫平均产卵量为78粒,最高可达121粒。在温度为22 ℃、湿度为85%时,菜豆象卵的孵化率为91%,而在温度为32 ℃、湿度为62%时,卵的孵化率仅为26.3%(李南植等,1995;番启山等,1994;Howe & Currie,1964)。因此,菜豆象适宜在如上区域建立种群并大规模扩散,本预测结果准确度较高。
生态位模型通常用来描述物种的基本生态位,而非实际生态位。实际生态位通常小于基本生态位。使用生态位模型进行物种潜在分布预测时,多种影响物种分布的生物与非生物因素被忽视。本研究通过筛选仅选取了7个影响其潜在分布的主要生物气象数据,并未考虑寄主分布、天敌捕食等生物因素和土壤类型、地理特征、自然地理屏障、人类行为等非生物因素(Broennimannetal.,2007;Giovanellietal.,2008; Guisan & Zimmermann,2000)。菜豆象的几次大规模入侵和扩散都与菜豆的大量进口或引种种植有关。如1914—1918年的第一次世界大战等事件导致欧洲肉类奇缺,大量进口菜豆以代替动物蛋白。菜豆象在这一时期从欧洲南部地区扩散到了北部奥地利的维也纳地区,法国的塞纳河地区(刘永平和张生芳,1995)。菜豆等寄主植物的分布及扩散可能对菜豆象的入侵扩散尤为重要。
随着我国人民生活水平的提高,豆类的需求日益增高,种植面积与进口豆类数量的持续增长为菜豆象入侵提供了便利。云贵高原、四川盆地、广东、浙江和福建沿海区域作物结构复杂,菜豆象寄主广泛,防治工作形势严峻。依据联合国粮农组织(Food and Agriculture Organization of the United Nations, FAO)国际植物检疫措施(international standards for phytosanitary measures, SPM)标准中PRA原则与外来有害生物风险评估多指标综合评价体系的基本要求,需要提高对菜豆象的诊断能力、检验检疫水平,控制菜豆象的入侵和扩散。建议目前已经发生菜豆象疫情的地区在生产、运输、仓储环节方面加强检疫及防治工作,积极开展菜豆象生物学、生态学、预测和防控方法等方面的研究。
