三峡库区城市富营养化河流凋落物的混合分解效应
2021-09-03豆鹏鹏林敦梅黄屿玥姚婧梅重庆大学三峡库区生态环境教育部重点实验室重庆400044中国农业大学草业科学与技术学院北京0093
豆鹏鹏 ,林敦梅 ,王 芳 ,黄屿玥 ,高 杰 ,杨 巍 ,韦 冰 ,李 伟 ,姚婧梅 * (.重庆大学,三峡库区生态环境教育部重点实验室,重庆 400044;2.中国农业大学草业科学与技术学院,北京 0093)
河流具有调节气候、改善生态环境、维持生物多样性等多种作用,为人类社会提供了重要的生态系统服务.同时,由于和人类活动的紧密联系,河流成为目前世界上受威胁最严重的生态系统之一.人类的生产生活向河流排放了大量的氮、磷等元素,导致河流富营养化,引起水质的恶化和生物多样性的降低,严重威胁着河流的生态系统功能与服务[1],包括河岸带植物凋落叶的分解功能[2].凋落物的分解作为河流重要的生态过程和功能之一,为河流生态系统提供了重要的物质和能量来源[3],对于维持生态系统碳和养分平衡具有重要意义,常作为评价河流生态系统健康的重要指标[4-6].
河流中的凋落物分解受到水体中养分(如氮、磷)含量、温度及流速等环境因素的影响[7-9],而底栖分解者类群(微生物和底栖动物)在叶片上的定殖和摄食活动等生物因素以及凋落物本身的理化性质更是影响其分解过程的关键因素.凋落物在自然界往往以混合物的形式存在,其混合分解效应已成为凋落物分解研究的热点[10], 相关研究多在陆地森林生态系统展开,在河流生态系统中的研究偏少,且已有研究多局限于源头溪流,鲜有研究关注城市富营养化河流的凋落物分解过程[11].关于底栖动物参与的富营养化河流中凋落物分解的混合效应未见报道,尚不清楚其分解过程是否与源头河流具有相同的格局.混合凋落物的分解可能与单种叶片分解的简单加和不同,产生正或负的非加和效应(协同或者拮抗),即质量损失实际值与以单种叶片推测的预期值不符,这种差异产生的原因可能是:(1)混合凋落物中,具有特殊性状的优势凋落叶种加快或者减慢了其他物种的分解[12].(2)随着混合凋落物间性状差异的增大,分解者可利用的资源类型可能增加(生态位增加),进而加快凋落物分解.(3)水体富营养化影响了水生生物群落组成[13],不同分解者类群(微生物、小型和大型底栖动物)对混合凋落物分解影响不同.
本研究以三峡库区重庆主城典型富营养化河流、嘉陵江的一级支流清水溪为代表,选取河岸 3种常见乔木叶凋落物为对象,通过原位分解实验,分析底栖动物参与叶凋落物在富营养化河流中的混合分解效应.以期更好地阐明富营养化河流凋落叶分解的特征与变化规律,为受损河流的生态环境恢复提供理论依据.
1 材料与方法
1.1 研究区概况
清水溪位于重庆市沙坪坝区(29°27'~29°46'N,106°14'~106°31'E),发源于歌乐山镇山洞村天灯杆,最终在磁器口金沙街汇入嘉陵江,是嘉陵江右岸的一级支流[14],流经生活区、工业区、商业区,受人为影响较大,为典型的富营养化水体,水质总体呈劣V类(表 1)[14].清水溪地处亚热带湿润气候区,气候温和,雨量充沛.区域年均气温为 17.8~18.6℃,年均降水量为 1085.1~1141.8mm,平均水温为 15.6℃(表 1).河岸带植被种类较少,常见的有构树(Broussonetia papyrifera)、刺桐(Erythrina variegata)、榆树(Ulmus pumila)、小叶榕(Ficus microcarpa)等半自然更新的乔木.

表1 凋落物分解实验中溪水的物理化学参数(数据为平均值,n=12)Table 1 Physicochemical parameters of the stream water during the litter decay (mean value, n = 12)
1.2 凋落物样品采集
选择清水溪河岸带3种常见树种(构树、刺桐和小叶榕)的叶片凋落物,于2017年8~9月收集,挑出没有破损和病虫害的叶片,在60℃烘箱中烘干备用.
1.3 凋落物原位分解实验
使用3种孔径(50μm、250μm和2mm)的分解袋:小孔径(50μm)只允许微生物进出分解袋,中孔径(250μm)允许微生物和小型底栖动物进出,大孔径(2mm)允许所有分解者(微生物、小型底栖动物和大型底栖动物)进出.称取烘干后的叶凋落物 4.2g (精确到0.1g)装入分解袋内(15×10cm)中,共计126个分解袋(7种凋落物处理×3种孔径×6个重复).7个处理中包含 3个单一物种凋落叶,即构树(B)、刺桐(E)、小叶榕(F)各4.2g,和4个混合物种凋落叶,即构树和刺桐各 2.1g(BE,4.2g)、构树和小叶榕各 2.1g(BF,4.2g)、刺桐和小叶榕各2.1g(EF,4.2g),3个物种各1.4g(BEF,4.2g).将分解袋绑在尼龙网上,并用石块将尼龙网随机固定在河床上.实验从2018年11月17日开始,于2019年 1月12日回收分解袋,将每一个分解袋装入自封袋中,低温保存运回实验室.取样同时测量了溪水的理化指标,并采集了水样和底栖动物样品.
1.4 水体理化性质与凋落物初始性状测试
测定的水体理化参数包括水温、pH值、电导率、溶解氧(DO)、总氮(TN)、总磷(TP)、氨氮、硝酸盐及磷酸盐等.其中,使用多参数传感器(YSI Professional plus,Yellow Spring,美国)现场测定溪水的温度、pH值、电导率和溶解氧.同时,收集水样,冷藏运回实验室.总氮采用过硫酸钾消解紫外分光光度法测定;总磷采用钼酸铵分光光度法测定;水样过0.45μm滤膜,采用纳氏试剂比色法测定氨氮含量;采用紫外分光光度法测定硝酸盐及磷酸盐含量[15].
测定的凋落叶初始理化性质包括纤维素、木质素、碳、氮、磷含量和叶片韧性.其中,使用范氏洗涤法测定木质素、纤维素含量[16];使用拉力计(ZP-50,艾固,中国香港)测定韧性,采用撕裂单位宽度叶片所用的力作为韧性值;使用 TOC-TN 分析仪(岛津,日本)高温灼烧测定有机碳含量;TN、TP先用浓硫酸-过氧化氢消解粉碎的样品,然后用TOC-TN测定氮含量,使用钼锑抗比色法测定磷含量.根据测定结果计算C/N、C/P、N/P与木质素/N.
1.5 底栖动物的分离与鉴定
将250μm孔径的索伯网套在50μm孔径的索伯网(30×30cm)中,采集了 3份小型底栖动物和大型底栖动物的平行样品[17].将样品存放于 75%的酒精中,运回实验室进行鉴定和计数.对大型底栖动物和小型底栖动物进行分类鉴定到尽可能低的分类水平.根据采样面积,换算出每种底栖动物的密度(ind/m2)并根据 Tachet等[18]的分类方法将大型底栖动物划分为不同的摄食功能类群:食碎屑者、撕食者、刮食者、滤食者、钻食者、捕食者和寄生者.
1.6 数据分析
根据凋落物分解前后的质量差异,计算凋落物的质量损失:
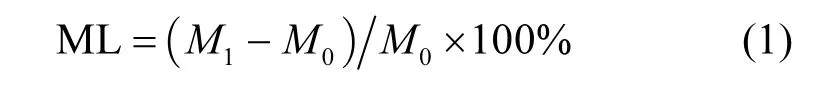
式中:ML为质量损失;M0和M1分别为凋落物分解前后的重量.同时根据单个物种的平均质量损失,计算混合凋落物的期望质量损失,并结合其实际质量损失,计算凋落物分解的混合效应以探究混合凋落物分解的质量损失是否可由其组成物种来预测[19].
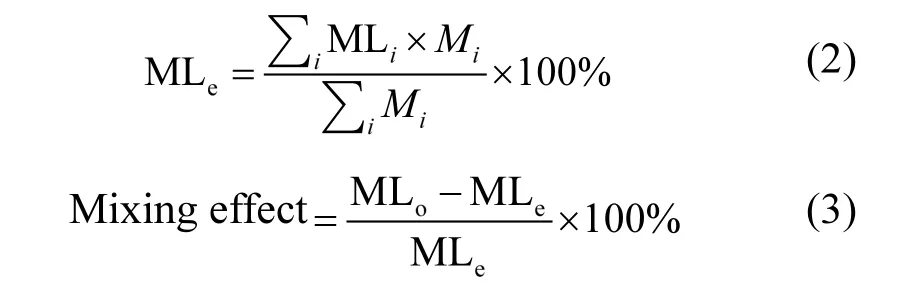
式中:MLe为混合凋落物质量损失期望值;MLi为物种i的质量损失;Mi为组合中物种i的重量;MLo为混合凋落物实际的质量损失.
根据Seastedt等[20]的研究量化了3种分解者(微生物、小型和大型底栖动物)对叶片重量损失的平均贡献:

式中:Efauna(%)是某种分解者对叶凋落物重量损失的影响;Lfauna是某种分解者造成的重量损失百分比,是由不同孔径分解袋中凋落物重量损失的差异得出的;Ltotal是大孔分解袋中重量损失百分比,是由微生物、底栖动物和其他效应导致的叶片分解.
选用混合凋落物性状的群落加权平均值(CWM)作为相应指数[21]:
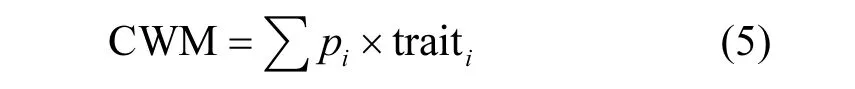
式中:pi是物种 i在混合凋落物中所占的比例;traiti是物种i的性状值.
混合凋落物的性状多样性由功能分散指数(FDis)表示,可描述多维特征空间中组成物种到混合物种质心的平均距离的功能多样性分量[22](R软件,“FD”包).
采用非参数组间差异的Kruskal-Wallis检验混合凋落物物种丰富度对质量损失的影响;配对t检验探究混合处理中的混合效应是否显著;双因素方差分析探究处理与分解袋孔径及其交互作用对质量损失和混合效应的影响,并对质量损失进行 Tukey HSD检验.根据Loreau等[23]的公式计算了混合凋落物的选择效应和互补效应.采用“lme4”包中的线性混合效应模型分析加权性状、性状多样性等解释变量对质量损失、混合效应等响应变量的影响[24].所有分析均在R软件(R version 3.4.2)中完成,P < 0.05定为统计上差异显著.
2 结果与分析
2.1 凋落叶的初始理化性质
3种叶凋落物的初始理化性质存在显著差异(表2).构树叶的N、P含量最高,而C和木质素含量、韧性及C/N、C/P和N/P均最低,因此其可降解性是3种叶片中最好的.刺桐叶的C、N、P、木质素含量与韧性,以及C/N、C/P介于其它物种之间,叶片可降解性居中.小叶榕的C和木质素含量、韧性及木质素/N、C/N和C/P也显著高于另外两个物种,其叶片可降解性最差.

表2 三种凋落物的初始理化性质(平均值±标准误,n=5)Table 2 Initial physical and chemical properties of three leaf species (mean ± SE, n=5)
对4个混合处理的各初始加权性状进行主成分分析,前两轴分别解释了总体变异的 95.1%和 4.9%.第一轴与凋落叶N、P含量负相关,与C含量、N/P、韧性、木质素含量、木质素/N、C/N、纤维素含量和 C/P正相关(图 1).因此,选用第一轴(CWM1)作为混合处理的初始性状.

图1 四个混合处理初始加权性状的主成分分析Fig.1 Principal component analysis of initial community trait of four mixture treatments
2.2 凋落叶的分解
凋落物分解的质量损失受凋落叶物种组成(F6,105=61.33,P<0.001)和分解者类群(F2,105=10.85,P<0.001)及其交互作用(F12,105=1.89,P<0.05)的显著影响,其中物种组成的解释度最大(图2,表3).同时,物种组成及其与分解者类群的交互作用也显著影响凋落物的混合效应(表 4).物种丰富度与凋落叶的质量损失无关(图3a-c).

表3 物种组成和孔径对质量损失影响的双因素方差分析Table 3 Two-way ANOVA of species combination and mesh size on mass loss

表4 物种组成和孔径对混合效应影响的双因素方差分析Table 4 Two-way ANOVA of species combination and aperture on mixing effect
2.2.1 凋落叶分解的质量损失 对于单个物种而言,无论何种孔径,构树在 3个物种中的质量损失均最多,且在大孔径(2mm)中达到83.97%(图2).刺桐和小叶榕的质量损失在 3个孔径下均相近,无显著差异.其中,与小孔径(50μm)相比,刺桐的质量损失在大孔径中由63.63%显著降低至49.62%.

图2 凋落物在不同处理和分解袋孔径下的质量损失Fig.2 Mass loss of litter under different treatments and mesh sizes
在混合凋落物中,只有微生物参与分解(50μm)时,质量损失为 53.90%(刺桐和小叶榕,EF)~72.37%(构树和刺桐,BE);小型底栖动物加入后(中孔径,250μm),质量损失为 49.10%(刺桐+小叶榕,EF)~63.55%(构树+刺桐+小叶榕,BEF);大型底栖动物加入后(大孔径, 2mm),质量损失为 51.03%(刺桐和小叶榕,EF)~67.78%(构树和刺桐,BE).
2.2.2 凋落叶分解的混合效应 4种凋落物组合在3种不同分解袋孔径下共计 12种混合处理,其中 6个均表现为显著负的非加和效应,即实际质量损失显著低于预期质量损失[图 3(d)-(f)].仅微生物参与分解时,构树+小叶榕(BF, P<0.01)、刺桐+小叶榕(EF,P<0.01)、构树+刺桐+小叶榕(BEF, P<0.05)的混合分解实际质量损失均显著低于其期望值.小型底栖动物加入分解后,上述组合中除了 3物种混合的实际质量损失与其期望值无显著差异(P=0.98),其他有显著差异.大型底栖动物进入分解袋后,所有组合中只有 BF(P<0.05)的实际质量损失仍显著低于各组成物种质量损失之和(图 3).其中产生显著负的非加和效应的组合中均包含小叶榕.
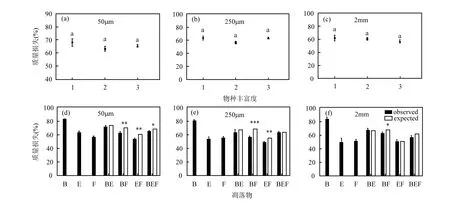
图3 不同物种丰富度和凋落物处理分别在3种分解袋孔径中(50μm、250μm和2mm)的质量损失Fig.3 Mass loss of litter in three mesh size (i.e., 50μm, 250μm, and 2mm) with species richness and treatment
2.3 分解者类群及其对凋落叶分解的影响
在清水溪中发现 8个大型底栖动物类群,平均密度为 4408个/m2,分属 4个纲(蛭纲、寡毛纲、昆虫纲和腹足纲),苏氏尾鳃蚓(食碎屑者)和静泽蛭(捕食者)为优势种,分别占比70.06%和24.45%;4个小型底栖动物类群(线虫、桡足类、寡毛纲和昆虫纲),平均密度为14287个/m2,猛水蚤和线虫为优势种,分别占比60.86%和30.54%(表5).
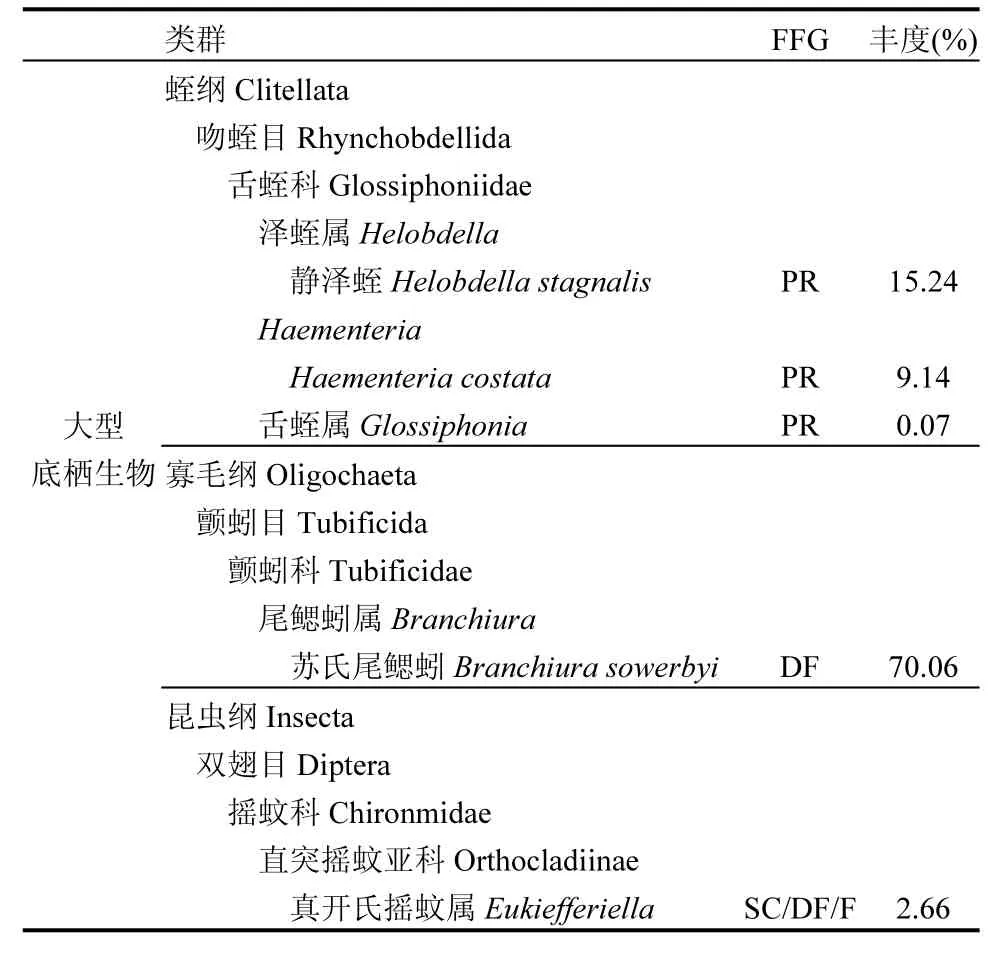
表5 清水溪中底栖动物的相对丰度(%)和功能摄食群Table 5 Relative abundance (%) and functional feeding group of invertebrates in Qingshuixi stream

续表5
微生物对富营养化溪流凋落叶分解的贡献最大,只有微生物参与分解时(50μm),混合凋落叶的质量损失最多(图4).整体来看,大型和小型底栖动物均抑制了凋落物的分解,且小型底栖动物的抑制作用更大(图4,a).

图4 不同分解者类群对质量损失的影响及其贡献度Fig.4 Impact and contribution of different decomposers on mass loss
2.4 凋落叶混合对其质量损失和混合效应的 影响
混合凋落物初始加权性状(CWM1)显著影响其质量损失(图 5a).随着加权性状 N、P含量的增加,混合凋落物质量损失增加,互补效应增强,选择效应减弱;随着C含量、韧性、C/N、木质素、纤维素含量的增加,混合凋落物质量损失减少,互补效应减弱,选择效应增强(图 5a,图 6).同时,随着混合凋落物的功能差异的增大,混合效应显著减小(图 5b),即混合分解的实际质量损失减少.

图5 混合凋落物初始性状及其功能差异对凋落物分解的影响Fig.5 Effect of initial traits and functional divergence of litter mixture on litter decomposition CWM,性状群落加权均值;FDis,功能离散度.C,碳含量;N,氮含量;P,磷含量
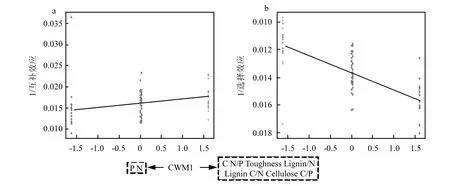
图6 混合凋落物初始性状对互补和选择效应的影响Fig.6 Impact of initial traits of litter mixture on complementary and selectivity effects
3 讨论
3.1 凋落叶的质量损失
富营养化河流(清水溪)中凋落叶性状是其质量损失的重要影响因素,与其他陆地和溪流中的研究结果一致[25-26].全球范围溪流生态系统中凋落物分解的meta分析表明,木质素含量和C/N是最重要的凋落叶性状[26].同时,可降解性较高的凋落物(高N、P含量和低C/N、C/P)更有利于微生物定殖[27-28],比可降解性较低的凋落物(低N、P含量,高C/N、高木质素含量)更快分解[29-30].具有高N和低C/N的构树的分解显著快于其他单个物种(图 1).而凋落物的木质素含量越高、韧性越大(如小叶榕),越不利于微生物定殖和底栖动物对凋落叶的破碎[31-32].
3.2 凋落物混合对其分解的影响
凋落物的混合分解在质量变化上的非加和效应在陆地[19,33-35]和非富营养化的水生生态系统[36-37]中广泛存在.随着混合凋落物功能离散度的增加,尤其组合中木质素等难分解物质的含量较高时(含小叶榕),混合凋落物分解的质量损失低于预期[图3(d)~(f)].其他研究也发现了凋落物混合后总体失重率的降低,可能是由于其中某种特殊凋落物的他感作用生成抑制微生物活性的次级代谢产物(如酚类等)干扰了凋落物的分解[38].也有研究发现混合凋落物的功能多样性显著促进了水生态系统凋落叶的分解[36].如 Zhang等[36]在林中水塘的研究发现了由含氮较高物种引起的功能多样性的增加对凋落物分解的促进作用,他们所选森林水塘生态系统贫营养化(缺乏N, P),因此微生物分解含氮较高的物种可以获得生命活动所需的氮, 凸显了含氮较高物种所带来的凋落物功能多样性增加的重要性;但富营养化河流为微生物提供了丰富的氮磷,氮磷元素不是微生物进行分解活动的限制因子,因此本研究中功能多样性的增加并没有促进混合凋落物的分解.
3.3 分解者类群对凋落物分解的影响
与微生物相比,底栖动物对凋落物分解的贡献较低,甚至为负(图4),这与非富营养化河流中各分解者类群对凋落物分解的影响显著不同[3,17,39],这可能主要是由于富营养化胁迫下底栖动物群落结构发生变化引起的.清水溪的小型底栖动物群主要是由猛水蚤和线虫两类组成,大型底栖动物主要以苏氏尾鳃蚓和静泽蛭为优势类群(表 5),主要的摄食群功能是食碎屑者(70.8%)和捕食者(24.4%),缺乏源头溪流中常见的直接参与落叶分解的撕食者(表 5)[40].底栖动物中主要通过直接取食叶片(撕食者)和通过改变叶片表面的微环境(生物膜结构、氧化作用、养分供应等)[41-42],或者通过对微生物的捕食作用间接影响凋落物的分解[42].本研究发现大孔径和中孔径的叶片质量损失均小于小孔径的质量损失,因此小型和大型底栖动物的加入显著抑制了微生物的落叶分解(图 4),推断底栖动物可能通过捕食作用对微生物分解凋落物起了负面效应.富营养化河流中底栖动物对微生物群落结构的影响可在今后进一步分析.此外,本研究主要通过不同孔径分解袋中剩余凋落物质量间的差异来衡量不同分解者类群的作用,而在凋落物分解过程中,不同分解者间可能存在协同、竞争等,在今后的研究中还需通过更精细的控制实验来量化这些复杂关系.
依据广泛应用的 Gessner等[4]提出的由落叶分解评价溪流健康的模式,健康溪流中大、小孔径分解袋凋落物分解速率的比值应为 1.2~1.5,而清水溪中的比值均小于 1(大孔径的质量损失低于小孔径),表明清水溪的生态功能已经受到严重影响,这可能主要是富营养化河流中底栖动物多样性严重下降引起的:清水溪的小型和大型底栖动物的香浓威纳多样性指数仅为 0.93和 0.97,远低于其他非富营养化河流(2.5~3.0)[17,43].因此,应该恢复底栖动物多样性,并检验其大、小孔径分解袋凋落物分解速率的比值是否恢复到1.2~1.5的范围中.
综合来看,由河岸带植物凋落物所带来的外源有机物对河流生态系统的营养输入十分重要,不同河岸带植被的组成会影响水体的营养盐负荷[44]以及水生食物网中的凋落物分解过程[45].依据植物的群落加权性状进行配置可能比单独增加或者减少河岸带植物的物种数量更能为富营养化水体的修复实践提供有益的理论参考.
4 结论
通过3个物种的不同组合以及3种孔径的分解袋设计,探明了三峡库区城市富营养化河流凋落物的混合分解效应.所有混合处理中,产生加和效应与显著负的非加和效应的处理各占一半,这主要是受凋落物性状的显著影响:混合凋落物的质量损失随群落加权性状N、P含量的增加而增加,随C含量、韧性、木质素含量、C/N、纤维素含量的增加而减少;随着混合凋落物功能离散度的增加,如组合中含高木质素等难分解物质的凋落物时,分解受到抑制.微生物在富营养化溪流凋落物分解中起重要作用,底栖动物的重要性降低,可能与其物种多样性下降有关,如缺乏在非富营养化溪流中普遍存在的撕食者类群.在富营养化河流的修复中,需同时考虑河岸带植物的配置与恢复水生生物的多样性.
