巽他陆架末次冰期植被之谜
2021-08-17郁金勇李丽贺娟贾国东
郁金勇,李丽,贺娟,贾国东
同济大学海洋与地球科学学院,海洋地质国家重点实验室,上海200092
巽他陆架位于南海以南的东南亚赤道区域,被中南半岛、马来半岛、婆罗洲以及部分印尼群岛包围[1]。末次冰期(110~11.7 kaBP)时海平面下降,巽他陆架广泛暴露。特别是末次盛冰期(LGM,23~19 kaBP),巽他陆架海平面与现代海平面相比下降约123 m,陆地面积几乎扩大一倍[2]。这种海陆面积的显著变化对当时的全球古气候、古生态、碳循环甚至古人类迁徙等方面产生了重要影响[3-4]。
在冰期-间冰期旋回中,陆地植被对于陆地碳储库有着重要影响。冰期陆地碳储库的数值估算最大的不确定来源就是出露的陆架[5],特别是在东南亚热带地区。亚马逊和刚果河流域的热带雨林分布由于受冰期的气候影响其面积会出现明显收缩[6],而海平面变化对这两个地区的陆地区域面积影响不大[7]。相反,冰期巽他陆架出露土地上的植被状况对陆地生物圈的碳储量估算会产生重要的影响。若陆架上覆盖的是热带森林,在冰期东南亚热带森林区的碳储量会与南美的亚马逊流域十分接近;若出露陆架上呈现的是草原景观,由于其碳密度仅为森林的1/10,则其储碳的全球意义就会大打折扣[3]。
冰期时巽他陆架上的植被状况仍存在争议,主要有两种观点:第一种是由Heaney提出的“热带稀树草原走廊”(图1)[8]。该走廊由泰国南部开始沿马来西亚半岛延伸,并穿越了婆罗洲和爪哇之间的暴露陆架。热带雨林主要集中在婆罗洲东部和北部、苏门答腊岛西部和北部[9]。在这种情况下,巴拉望岛和菲律宾西部也被认为是热带稀树草原[4]。第二种可能性是,在冰期雨林植被仍主导巽他陆架,并不存在“草原走廊”。
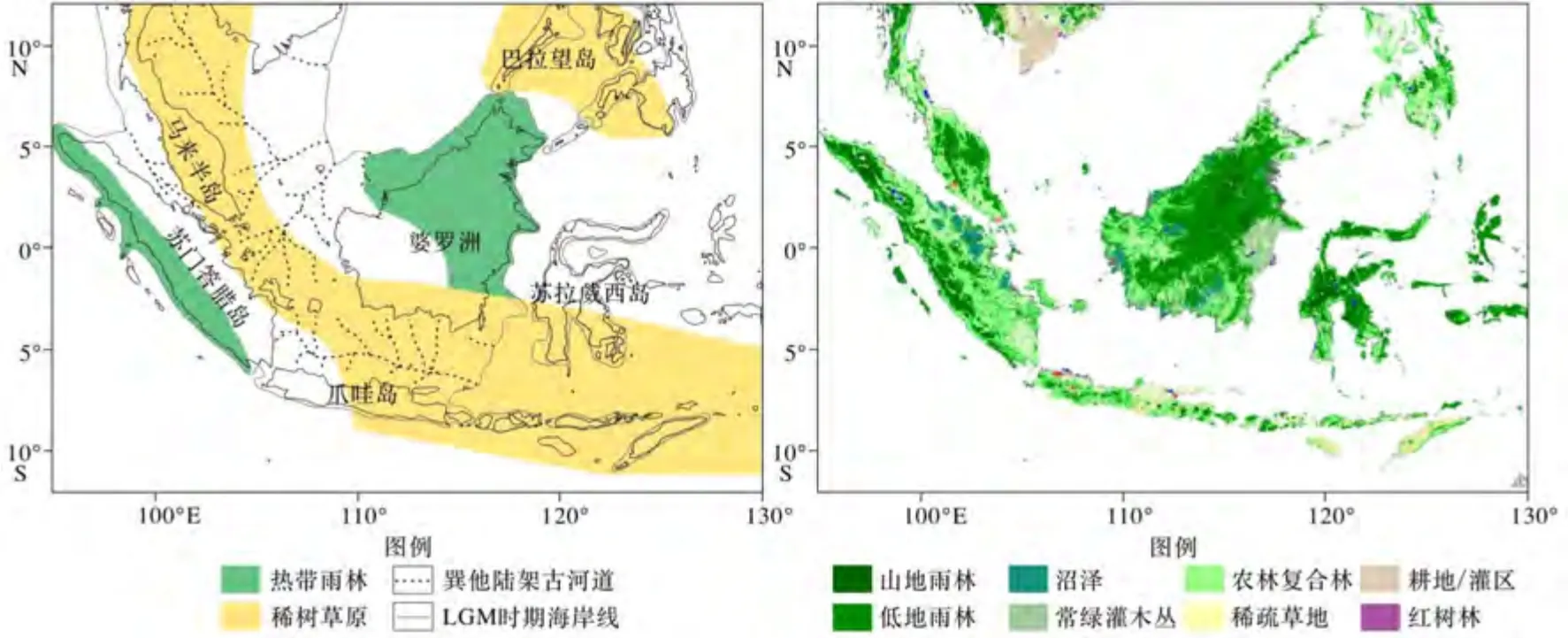
图1 巽他陆架“稀树草原廊道”假说(左)及现代植被分布(右)植被数据来自:https://ec.europa.eu/jrc/en/scientific-tool/global-land-coverFig.1 Hypothesis of "Savanna Corridor"on Sunda Shelf (left)and modern vegetation distribution (right)Vegetation data comes from: https://ec.europa.eu/jrc/en/scientific-tool/global-land-cover
目前研究古植被类型的方法主要有两种:一种方法是根据不同植被类型(C3和C4植物)之间的碳同位素(δ13C)分馏差异[10],根据所获得的δ13C值判断植被类型;另一种是直接通过沉积物中的植物化石(如孢粉、植硅体)鉴定,定性或半定量地分析源区不同属种植物的比例。本文汇总了现已发表的巽他陆架区域范围内末次冰期以来相关碳同位素、植物孢粉的研究成果,以期对末次冰期时巽他陆架植被状况有一个全面的认识。
1 现代气候与植被
巽他陆架位于印度-太平洋暖池区(Indo-Pacific Warm Pool,IPWP),在季节尺度上,气候受到赤道辐合带(Intertropical Convergence Zone,ITCZ)迁移的控制[11]。在IPWP和海洋大陆(Maritime Continent,MC)上,ITCZ由于温暖的地表水、相关的深层大气对流、陆地-海洋温度差异和季风的影响而增强[12]。MC降水的年变化明显表现为旱季和雨季,现代观测显示:从11月到4月强降雨主导了MC,5月开始MC上的降雨减少,在随后的几个月中ITCZ向北移动,在北半球秋季,降雨带逐渐返回MC地区[13]。东亚季风和澳大利亚-印尼季风在巽他陆架中部和北部形成了强降雨区[11]。这种气候导致今天婆罗洲大部分地区普遍湿润,低地雨林大量发育且该岛绝大部分地区的热带草相对丰度不足0.1[14]。而南部的苏拉威西岛、弗洛勒斯岛等地区的降水受澳大利亚-印尼夏季风控制明显,表现出明显的季节性变化[15-16]。在年际尺度上,ENSO可导致快速强烈的水文变化。MC降水的年际变化及其与ENSO的密切关系已得到充分认识,区域降雨在厄尔尼诺年期间减少,而在拉尼娜年期间则增加[17-18]。现代观测数据显示,在厄尔尼诺时期降水受到抑制,月平均降水量比正常平均降水量少32 mm;而在拉尼娜时期,平均月降水量比正常月份多39 mm[19]。在更长的时间尺度上(103~104年),日射周期变化、海平面变化、大西洋径向翻转流的强度以及陆-海、纬向和径向的温度梯度都会影响IPWP区域的降雨[20-24]。
东南亚现代植被主体为热带雨林,但也受到降水季节性变化(或干旱程度)的影响而有所变化(图1)。在赤道附近的核心区域,全年降水充足,以常绿雨林为主要植被类型如马来半岛、婆罗洲、苏门答腊岛和爪哇岛西部;而外围的植被主要为含一定落叶成分的季雨林,如中南半岛最南端、巴拉望岛以及爪哇岛中部和东部[25]。东南亚地区山地植被类型丰富多样,以婆罗洲为例,位于海拔1 200 m以下的热带低地雨林以龙脑香科植物为主,并伴生其他种类丰富的热带植物;在海拔1 200~1 500 m,植被主要为热带低山雨林;热带高山雨林则位于海拔1 500~3 000 m或3 500 m的 地 带;当 海 拔 超 过3 000 m或3 500 m时,热带亚高山森林开始出现[19]。
2 末次冰期的植被记录
2.1 碳同位素记录
外形丰富、多姿多彩的植物群落在植物死亡后,很快会腐烂消亡而难以追溯,文物古迹和历史记载等只可了解较短历史的区域植被历史。更长时间的地质记录还需借助易保存且不会发生地质转化的指标来实现。
在自然生态系统中,植物在酶参与下通过光合作用吸收二氧化碳,将光能转变为化学能,储存在有机物中,除提供植物本身能量外,还是其他动物、微生物以及人类活动的能量来源。在此过程中由于不同类型植物的碳同化途径的差异(Calvin循环和Hatch-Slack循环),产生了显著的碳同位素分馏效应,而关键步骤的有机物分别是3-磷酸甘油酸(含3个碳原子)和草酰乙酸(含4个碳原子),因此具有该同位素分馏特征的植被分别形象地称为“C3植被”,“C4植被”,其碳同位素分馏范围分别为-22‰~-35‰和-9‰~-19‰[10,26]。此外在陆地植被中还存在景天酸代谢循环(CAM循环),其碳同位素变化范围介于C3、C4植被之间,而其主要植被为如景天属、仙人掌属等肉质植被,多分布于干旱的沙漠环境,不是常规环境分布植被类型,因此不包含在一般的植被类型讨论中。植物光合作用产生的碳同位素分馏可以很好地保存在合成的碳物质中,即使在后期的沉积和埋藏过程中,也几乎不会发生进一步的碳同位素分馏,且C3和C4植物的碳同位素组成具有明显不同的分布范围,因此其是重建植被变化的理想代用指标,被广泛应用于土壤、湖泊、海洋沉积的古植被重建[27-28]。
目前巽他陆架区域内可供古植被重建的碳同位素记录来自于洞穴粪便中的昆虫壳体、海洋或湖泊沉积物中的叶蜡脂类生物标志物(表1,图2)。
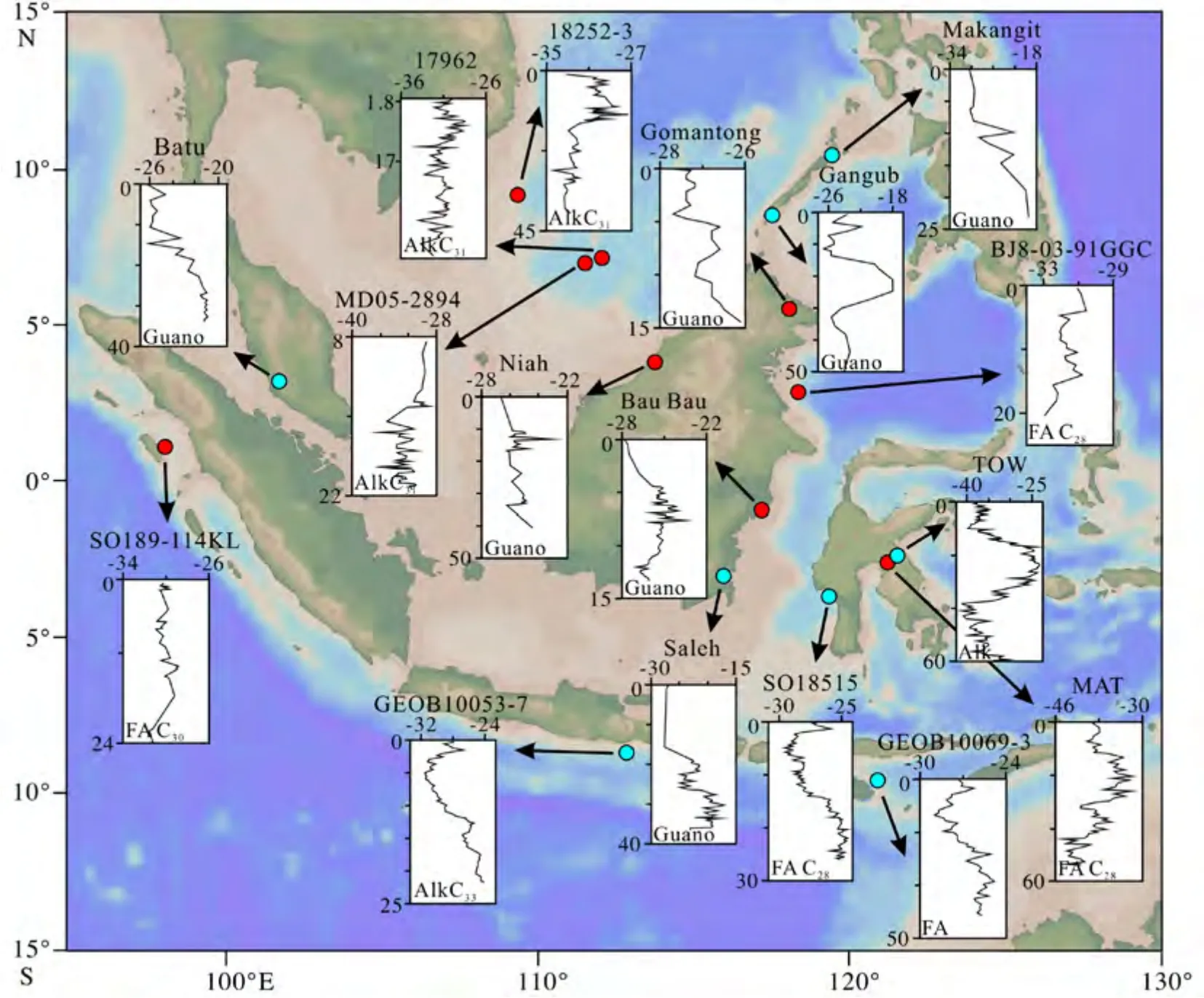
图2 巽他陆架及其周边地区已发表碳同位素记录汇总碳同位素图上部的数字为碳同位素值,左边的数字为年代,单位:ka,左下角为所用的生物标志物,岩芯位置信息见表1。Fig.2 Summary of published carbon isotope records in the Sunda shelf and surrounding areasThe upper number of the carbon isotope map is the carbon isotope value,the number on the left is the age,unit:kaBP,the lower left corner is the biomarker used,and the core location information is shown in Table 1.
2.1.1 洞穴粪便碳同位素
东南亚地区的洞穴中生存着大量以昆虫为食的金丝燕和/或食虫蝙蝠,它们的粪便堆积可形成几米厚的沉积物[9]。昆虫的δ13C值由饮食决定,昆虫表皮的δ13C值可反映其进食的植被类型[29]。食虫动物粪便主要由昆虫表皮组成,因此,其δ13C可以反映区域的植被类型[30]。金丝燕和食虫蝙蝠通常在栖息地15 km范围内觅食[31],可认为其反映了局部区域的植被信息。此外,洞穴粪便沉积物适合进行放射性碳年代测定[32]。东南亚地区洞穴中的食虫动物粪便已被证明可以通过碳同位素记录提供周围区域的植被变化记录[9,14,33]。
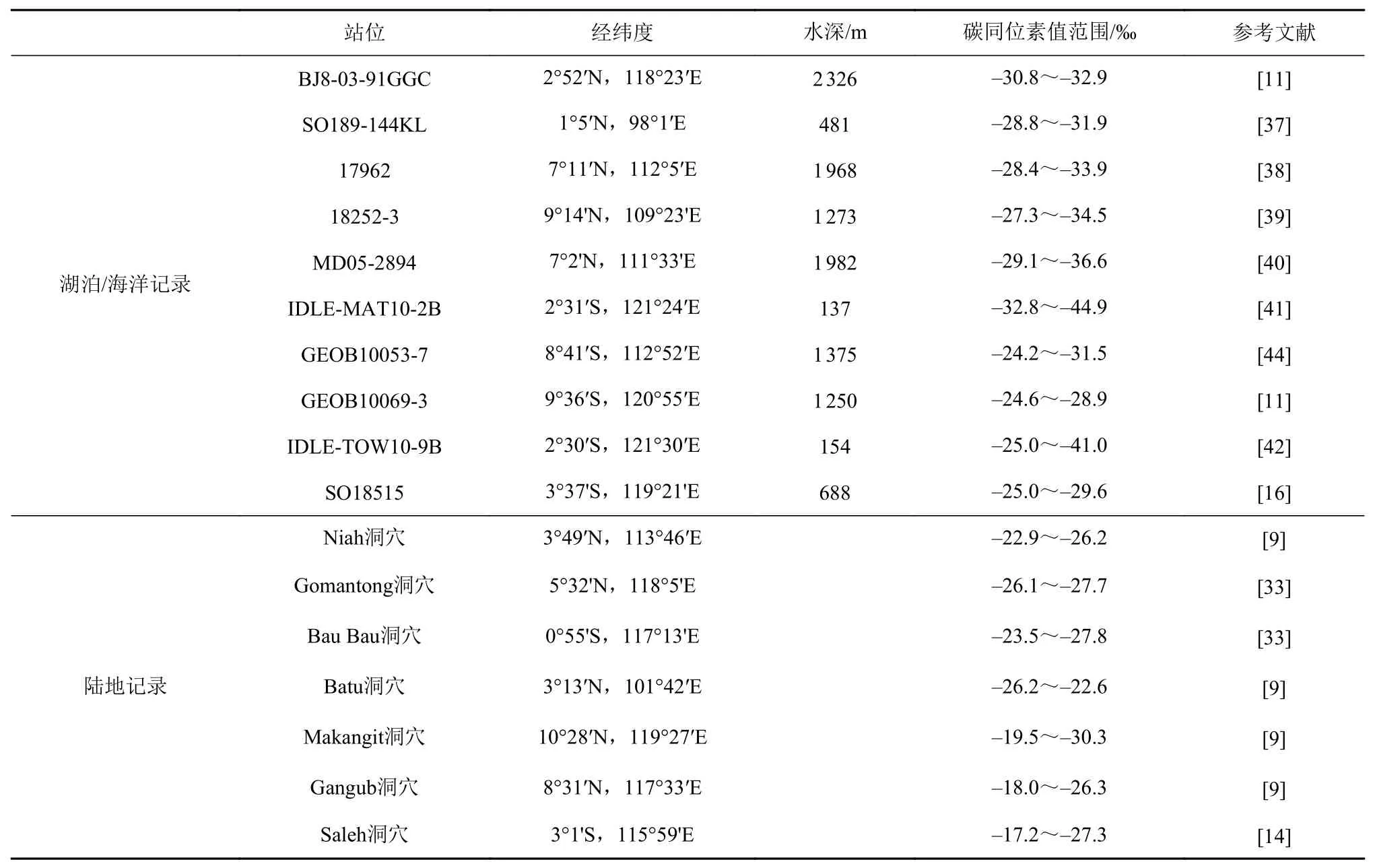
表 1 巽他陆架及周边地区末次冰期碳同位素变化记录Table 1 Carbon isotope records in the Sunda shelf and surrounding areas during the last glacial period
婆罗洲北部Niah洞穴的δ13Cguano值除了在13.4 kaBP短暂上升至-22.9‰外,在冰期的时间段一直处于-24.7‰至-26.2‰之间,C3植物在冰期持续存在于该地区[9]。婆罗洲东北部的Gomantong洞穴和中部的Bau Bau洞穴碳同位素记录也显示这两个洞穴周边地区在冰期被C3植物覆盖[33]。
婆罗洲南部的Saleh洞穴的δ13Cguano在冰期达到了-17.2‰的高值,显示了C4植物在冰期的大规模扩张,全新世之后,热带雨林重新出现在这一地区[14]。此外,Saleh洞穴的记录也显示出植被变化与IPWP的日照和海面温度有很强的对应关系,在冰期-间冰期旋回中,植被对热带气候变化具有很高的敏感性[14]。马来半岛南部的Butu洞穴的δ13Cguano在35~16 kaBP一直保持在-22.6‰之上,C4植物是该地区冰期植被的主要成分[9]。巴拉望岛南部的Gangub洞穴在冰期发生了明显的C4植物扩张,δ13Cguano由-26.3‰上升至-18.0‰[9]。
2.1.2 叶蜡脂类生物标志物碳同位素
长链正构烷基化合物是陆地维管植物叶片表皮蜡的主要成分,包括正构烷烃、正构烷醇、正构烷酸和蜡脂[34]。这些叶蜡脂类可以附着在黏土或细小颗粒物上,通过风或河流的搬运沉积在湖泊和海洋中,成为记录源区植被信息的生物标志物[28]。受合成途径影响,相比植物的总碳同位素,脂类标志物的碳同位素相对更偏负,一般C3植物叶蜡正构烷烃δ13C的范围为-29‰~-39‰,C4植物叶蜡正构烷烃δ13C的范围为-14‰~-26‰[35]。其他叶蜡脂类如正构烷醇和脂肪酸也有着类似的碳同位素分馏现象,C3植物叶蜡脂肪酸δ13C的范围为-30.8‰~-41.8‰,C4植物叶蜡脂肪酸δ13C的范围为-19.3‰~-21.6‰[11,36]。因此,叶蜡脂类δ13C变化可以反映源区植被类型的变化,重建陆地古植被。
婆罗洲东北部的BJ8-03-91GGC孔的脂肪酸δ13C值在冰期介于-30.8‰至-32.9‰之间,显示冰期C3植物在婆罗洲东北部地区持续存在[11],这与附近的Gomantong洞穴的碳同位素记录相一致。苏门答腊岛西北部的SO189-144KL孔脂肪酸δ13C值在冰期维持在-30‰左右,变化幅度不超过2‰,苏门答腊岛西北部在冰期始终保持着C3植物优势,同一钻孔的δD数据也显示冰期和全新世相比降水量变化不大,该地区在冰期气候湿润[37]。南海南部17 962孔 正 构 烷 烃C31的δ13C值 范 围 为-33.9‰~-28.4‰[38]。在冰期暴露的巽他陆架北部在冰期被C3植物覆盖,并没有出现C4植物扩张的现象,同样的结论也出现在附近的18 252-3孔[39]和MD05-2894孔[40]中。苏拉威西岛中部IDLE-MAT10-2B孔的碳同位素记录显示在冰期该地区周围由C3植被主导[41],但这与附近其他的植被记录如IDLE-TOW10-9B孔的碳同位素记录[42]和Wanda剖面的孢粉记录[43]相矛盾。分析认为该现象与海拔高度和地貌差异有关,该钻孔周围为陡峭的高山,这些高山在末次冰期保留着热带山地雨林,抵消了冰期C4植物扩张造成的δ13C值升高的影响[16]。
东爪哇岛南部沿海的GEOB10053-7孔主要反映低地植被的正构烷烃C31和C33的δ13C记录显示在冰期东爪哇低地C4植物占优势,在LGM时期C4植物比例可达50%[44]。萨武海的GEOB10069-3孔脂肪酸的δ13C在LGM时期达到了-24.6‰的高值,C4植物在此时发生了大规模扩张[11]。爪哇岛和萨武海附近C4植被的优势表明干旱季节的强度和持续时间增加,冰期暴露的萨胡尔陆架也可能减少了秋冬季节来自太平洋的水分通量[11]。苏拉威西岛中部IDLE-TOW10-9B孔[42]和曼达尔湾SO18515孔[16]也表现出冰期δ13C值升高、C4植物扩张的现象。巴拉望岛北部Makangit洞穴沉积物中正构烷烃C29和C31的δ13C值在LGM时期达到了-19.5‰,表明洞穴周围地区在冰期时被C4植物覆盖[9]。
2.2 植物孢粉记录
碳同位素研究植被变迁是近50年随着同位素技术发展而衍生的新技术,更早的研究主要是借助于植物生殖细胞:由被子植物和裸子植物所产生的花粉以及由其他蕨类植物和苔藓植物所产生的孢子,通常称为“孢粉”。孢粉不仅形态多变,可用于不同种属间的辨识,而且其外壁质密而坚固,可形成化石,保存在不同时代的地层中,记录了过去植物群落的组合特征,被广泛应用于植被演化重建[45-46]。
戴璐等曾对东南亚—澳大利亚北部地区的孢粉记录有过统计,探讨该地区末次冰期植被的演变特征[47]。本文在此基础上增补了巽他陆架地区更多的孢粉记录(表2,图3),以期对该区域冰期旋回中的植被变化有更全面的了解。

图3 巽他陆架及其周边地区已发表孢粉记录汇总孢粉图左侧的数字为年代(不均匀变化),单位:ka,岩芯位置信息见表2。Fig.3 Summary of published pollen records in the Sunda shelf and surrounding areasThe number on the left side of the pollen map is the age(uneven change),unit:kaBP,the core position information is shown in Table 2.

表2 巽他陆架及周边地区末次冰期孢粉记录Table 2 Pollen records in the Sunda shelf and surrounding areas during the last glacial period
外巽他陆架上18300柱记录了巽他陆架北部4万年来的植被信息:在冰期大部分时间段,以热带低地雨林为主的乔木花粉含量为70%左右,以禾本科和莎草科为主的草本植物为30%左右;在LGM时期草本植物含量增长到50%,但反映干旱与湿润气候的植硅石类型含量大体相等,说明此时湿度略微降低,但总体并不干旱[48]。类似的结论也出现在附近多个深海钻孔如17962[49]、17964[50]、18287[51]、18302[48]、18323[48]和CG2孔[19]。此外在多个钻孔记录中,LGM时期桫椤等蕨类植物的含量剧增到LGM前的几十甚至上百倍,Wang等据此推测LGM时期巽他陆架北部的植被并不同于先前的认识:既不是稠密的热带区,也不是热带稀树草原,而是在沿古巽他河流域的周围地区发育沼泽或湿地植被,即以桫椤为主的蕨类植物,以及以沼生禾本科和莎草科为主的草本植物;而在广阔的巽他陆架平原上覆盖有大量的雨林植被,如热带低地雨林和低山雨林[48]。巽他陆坡处的NS07-25和CB-19孔靠近婆罗洲北部,在冰期记录了巽他陆架和婆罗洲北部的植被信息。NS07-25孔在冰期的孢粉以热带低山雨林为主(40%~80%),LGM时期草本植物含量略有增加(不超过20%)[52]。附近的CB-19孔则以蕨类孢子为主,其次是高山雨林,草本植物的含量很少(最多不超过15%)[53]。值得注意的是,在这两个钻孔的孢粉记录中,LGM时期热带高山雨林的成分显著增加而低山和低地雨林成分减少,婆罗洲北部的Niah[54]和Kelabit[55]的孢粉记录中也出现了这一现象。这一现象反映出LGM时期温度降低,热带高山雨林向下扩张,分布面积增大,压缩了低山和低地雨林的生存空间。总的来说,巽他陆架北部及婆罗洲北部在冰期较湿润,在LGM时期降水略微减少但不足以使植被类型发生大规模变化,冰期这一地区热带雨林占主导。
婆罗洲中部的Sentarum湖的孢粉记录显示该地周边2.6万年来的植被组成非常稳定,低地雨林含量维持在80%左右[56]。苏门答腊岛中部高原的Di-Atas孢粉记录显示该地区自末次冰期以来始终被热带山地雨林覆盖[57],北部的Sim Sim钻孔也显示该地区森林在冰期占主导,草本植物孢粉比例不超过20%[58]。马来半岛的Nee Soon[59]和Nong Thale Song Hong(下称NTSH)[60]的孢粉记录在冰期也以热带雨林成分为主。菲律宾南部深海钻孔MD06-3075的孢粉记录中草本植物含量很少(不超过20%),主要是热带低地雨林,且在LGM时期该地区也发生了热带高山雨林向低海拔地区下移的现象[61]。
婆罗洲西部的Misedor孢粉记录在LGM时期出现了明显的草本植物孢粉增多的现象,变化幅度可达40%[62]。类似的现象也发生在苏拉威西岛的Tondano湖[63]和哈马黑拉岛Halmahera[62]的孢粉记录中。在苏拉威西岛中部的Wanda孢粉记录中,热带低地雨林和山地雨林成分仍然占主导,以禾本科为主的草本植物在冰期某些时刻也有增多,可能代表略有干旱[43]。结合区域内其他植被记录如碳同位素记录,该地区在冰期可能呈现山地雨林植被扩张、低地草本植物扩张的状态[16]。苏门答腊岛西南岸外的BAR94-42钻孔在冰期草本植物花粉明显增多,平均含量(30%~40%)远高于全新世[64],爪哇岛西部的Rawa[65]和Bandung[62,66]以及G6-4[67]钻孔也有类似的现象。而在澳大利亚北部的SHI-9 014钻孔中,冰期以草本植物占主导,草本植物孢粉含量可达50%以上[68]。
3 巽他陆架末次冰期植被分布格局
3.1 热带雨林带状分布
综合巽他陆架区域内的碳同位素和植物孢粉记录,冰期植被分布受地理位置显著影响,可大致分为冰期暴露的巽他陆架北部地区、靠近赤道区域以及相对远离赤道的区域。南海南部众多的深海钻孔记录均显示在冰期暴露的巽他陆架北部地区被雨林植被覆盖,冰期时叶蜡烷烃碳同位素值为-27‰~-37‰,是典型的C3植被占主导的特征,植物孢粉记录以热带低地雨林为主(主要为乔木花粉),未发生草本植物大幅扩张的现象[38-40,48-51]。靠近赤道区域包括婆罗洲中部及北部、苏门答腊岛中部等区域。婆罗洲北部Niah和Gomantong洞穴粪便碳同位素及东北部沿岸的BJ8-03-91GGC钻孔脂肪酸碳同位素均显示冰期C3植被在该区域持续存在,中部Sentarum湖2.6万年来的孢粉记录中低地雨林含量维持在80%左右[9,11,33,56]。苏门答腊岛中部的Sim Sim及Di-Atas孢粉记录及沿岸的SO189-144KL钻孔脂肪酸碳同位素都显示该地区在冰期被热带雨林覆盖[37,57-58]。相对远离赤道的区域包括马来半岛、巴拉望岛、婆罗洲南部、苏拉威西岛及爪哇岛等区域。马来半岛Batu洞穴δ13Cguano在35~16 kaBP保持在-22.6‰以上,表明C4植被在这一时间段主导了该地区的植被覆盖[9]。巴拉望岛南北两端两处洞穴粪便碳同位素记录也显示该岛在冰期发生了大规模的C4植被扩张[9]。婆罗洲南部Saleh洞穴粪便碳同位素以及Misedor孢粉记录显示该地区在冰期发生了大规模的草本植物扩张,草本植物孢粉比例增加了40%[14,62]。苏拉威西岛中部IDLEMAT10-2B孔植被记录看似与周边基础记录发生了矛盾,但这是由于该孔周围为山地,冰期山地雨林向下扩张抵消了低地草本植被扩张造成的碳同位素值升高。结合苏拉威西岛其他植被记录来看,该岛在冰期也发生了草本植被扩张的现象[41]。爪哇岛上Rawa和Bandung孢粉记录及沿岸的GEOB10053-7孔烷烃碳同位素均显示该地区在冰期草本植被扩张,Rawa剖面草本植被花粉在冰期占比达到60%,GEOB10053-7孔碳同位素值反映了东爪哇冰期低地C4植被优势,在LGM时期C4植被比例可达50%[44,62,65-66]。
如上所述,冰期时巽他陆架相对远离赤道的区域,如马来半岛、巴拉望岛、婆罗洲南部及爪哇岛等区域草本植物扩张,这些区域被开放植被覆盖。然而,开放植被并不能广泛替代赤道区域的热带雨林,冰期时热带雨林主要分布在冰期暴露的巽他陆架北部地区、婆罗洲中部及北部和苏门答腊岛中部(图4)。龙脑香科是东南亚热带雨林的主要成分,可以作为热带雨林的代理,Raes等使用基于317个龙脑香种(Dipterocarp)的物种分布模型来估算巽他陆架热带雨林在LGM时期的地理范围[69]。该模型认为,在LGM时期巽他陆架中部地区气候可以维持热带雨林生存,并且冰期暴露的巽他陆架北部地区是龙脑香林的分布中心,不太可能出现所谓的“热带稀树草原廊道”[69]。此外,基于现代植被和环境因子的植被分布区模拟显示,即使在现代非洲气候条件下,巽他陆架的稀树草原群落可能主要分布在中南半岛、爪哇岛等远离赤道的外围地区[70]。基于冰期巽他陆架区域内碳同位素和植物孢粉记录综合分析并结合相关的模拟结果,本文认为,冰期巽他陆架不太可能存在跨越巽他陆架的“稀树草原走廊”,热带雨林仍主导着巽他陆架。与全新世相比,冰期热带雨林分布范围向赤道方向收缩,呈现不规则的带状分布,婆罗洲北部、苏门答腊岛中部及冰期暴露的巽他陆架北部地区可能是当时热带雨林分布的中心(图4)。

图4 末次冰期巽他陆架植被分布两条黑色实线区间为现代热带雨林分布范围,黑色虚线区间为本文推测的收缩的冰期热带雨林分布范围。Fig.4 Vegetation distribution on the Sunda shelf during the last glacial periodThe two black solid line intervals are the distribution ranges of modern tropical rainforests,and the black dashed lines are the shrinking ice age tropical rainforest distribution ranges estimated in this article.
造成这一植被分布的机制,可能与区域的大尺度环流有关。末次冰期时,北半球高纬地区变冷,同时北极扩张的冰盖减弱了海洋的热量释放,进一步加剧了北半球高纬度区的降温[71]。而南半球夏季辐射量增强,温度有所升高,南北半球间的温度梯度增加[72]。这种增加的温度梯度增强了东亚冬季风和澳大利亚-印尼夏季风[19,22,73],并推动ITCZ的向南移动[23]。热带降雨带的推移过程响应ITCZ平均位置的向南移动,使得巽他陆架区域降水减少。ENSO是影响东南亚地区水文气候的另一个重要因素,婆罗洲地区的观测显示该地区的降水量受到ENSO强弱的显著影响[74]。赤道东西太平洋的纬向温度梯度显示在末次冰期时,热带太平洋呈现El Niño-like状态[75]。末次冰期时ITCZ的向南移动和热带太平洋的El Niño-like状态导致了巽他陆架区域的整体降水减少。但相对来说,靠近赤道的区域由于降水季节性较弱受到的影响较小,植被类型没有发生明显变化,仍以雨林植被为主;而相对远离赤道的地区由于季节性较强降水减少,植被类型由雨林向开放植被转变。在冰期暴露的巽他陆架北部虽然可能降水量减少[23,76],但因气团途径南海增加了水汽来源等因素,整体相对湿润[38,77]。降水量减少的幅度也不足以使植被类型发生变化[52],因此植被类型以雨林植被为主。
3.2 热带雨林垂直分布结构演化
山地在巽他陆架末次冰期植被垂直分布结构的演化中起着重要作用。高山雨林植被分布范围是1 500~3 000 m的高山地带,通常指示较为寒冷的环境条件[51,60]。并且可以在气候变冷的情况下,向低海拔迁移来适应气温的下降[54,68]。
在湿而冷的地区,高山雨林的分布范围向下扩展,压缩了低山和低地雨林的生存空间。南海南部CB-19孔的孢粉记录显示,在冰期随着温度的降低,低山和低地雨林分布范围逐渐减小,在LGM时期,高山雨林孢粉达到高峰,表明温度下降幅度扩大,高山雨林的分布范围也向下扩展[53]。附近的NS07-25孔也出现了同样的现象[52]。在婆罗洲北部Niah剖面,低山和低地雨林孢粉在冰期显著减少而高山雨林成分显著增加[54]。这是由于婆罗洲北部陆架较窄,不足以为植被提供伴随温度降低而向山下发展的空间,因此低山和低地雨林的生存空间被极大压缩。
在干而冷的地区,低地雨林面积收缩草本植物扩张,而高海拔地区相对湿润,仍保留着雨林植被。山地扮演着“雨林避难所”的角色,并且这些高山雨林在一定条件下可以向下扩张。苏拉威西岛中部托武帝湖IDLE-TOW10-9B孔的δ13Cwax值在冰期明显升高,显示了周围地区草本植物的大规模扩张[42],而不远处的马塔诺湖IDLE-MAT10-2B孔的δ13Cwax值在冰期并没有发生明显变化[41]。海拔高度和地貌差异导致了这种情形的产生:托武帝湖周围为平坦的河谷,而马塔诺湖周围环绕着高且陡峭的山脉。在冰期,草本植物在托武帝湖周围的河谷大规模扩张造成了δ13Cwax的显著升高,而马塔诺湖周围的高山雨林在冰期向下扩张,可能在一定程度上抵消了草本植物扩张造成的δ13Cwax升高[16]。同一地区的Wanda剖面的孢粉记录也证实了这一点[43]。该处的孢粉记录显示,在冰期,低地和低山雨林面积收缩而高山雨林分布范围扩大。同时,草本植物成分在冰期也出现了明显增加。在东爪哇,Ruan等利用正构烷烃C29、C31和C33单体碳同位素研究冰期植被变化。δ13C31/33记录反映了冰期东爪哇低地的草本植被扩张和降雨季节性增加;δ13C29反映了山地雨林的冠层结构保持封闭,山地降雨季节性相对不变[44]。苏门答腊岛西南部的BAR94-42孔的孢粉记录显示,与全新世相比,冰期草本植物成分显著增加,高山雨林孢粉比例在冰期保持相对恒定[64]。
4 巽他陆架古植被重建工作难点
冰期海平面下降,巽他陆架广泛暴露。出露的巽他大陆架地势较为平坦,完全不同于现代东南亚地区以山地为主的地形。在地势平坦的区域和河口处,可能广泛分布淡水沼泽林、泥炭沼泽林和红树林,这些植被主要受制于局部环境,主要组成成分不同于典型的热带低地雨林[47]。此外,在暴露的巽他陆架中部地区部分覆盖有砂质土壤,可能会形成荒地森林、泥炭沼泽林,并且这些森林即使在湿润的条件下也具有开放森林和封闭森林的特征[69]。在冰期暴露的巽他陆架上,这些可能的隐域性植被会影响我们对典型热带低地雨林的理解。
目前巽他陆架上已发表的大多数叶蜡同位素记录都基于单一化合物。例如在叶蜡脂的正构烷烃同系物同位素研究中通常使用n-C29、n-C31或n-C33,因为这些同系物通常在植物和沉积记录中都是最丰富和普遍存在的[28]。如果观察到几种同系物的同位素值变化之间存在很强的相关性,那么同位素研究也会将重点放在一种同系物上[41]。但是,这种单一化合物的方法往往会忽略通过比较更多同系物的同位素组成而提供的其他信息。而区域内碳同位素记录的另一来源——洞穴粪便也被质疑其代表性,因为大多数蝙蝠和金丝雀更倾向于在森林中觅食[54]。此外,利用碳同位素值计算C3/C4植被比例的二元模型在C3/C4植被δ13C端元值的选择上也没有达成共识[9,14,44]。
在南海南部,低地雨林是最为广泛的森林类型,主要由龙脑香科植物组成,然而此类植物的花粉产量极少,在龙脑香林的表土孢粉组合中其百分比也不足5%。此外,热带雨林中仅有很少的花粉能够传播至雨林之外,这使得从孢粉化石组合中正确识别热带雨林植被变得困难[47]。区域内已有的孢粉记录也存在着记录不连续、沉积或采样分辨率不足而难以识别快速气候事件,仅能提供植被变化的一般性概述[54]。
在降水丰富的热带地区,河流是花粉向海洋传播的主导途径[47]。冰期暴露的巽他陆架上发育着古巽他河、古湄南河、古湄公河等大型河流,在上述古河流河口处的孢粉沉积可能反映上游较为广阔区域的植被面貌,然而,当前的深海钻孔并没有覆盖这些古河口区。此外,苏门答腊岛和婆罗洲之间的区域缺乏相关数据,仍属空白,对这一区域的植被类型无法作出判断。植被记录的空间分布制约着我们对巽他陆架整体植被状况的认识。构的演化中起着重要作用。在湿而冷的地区,高山雨林的分布范围向下扩展,压缩了低山和低地雨林的生存空间。在干而冷的地区,低地雨林面积收缩草本植物扩张,山地扮演着“雨林避难所”的角色。
巽他陆架古植被重建工作仍面临着隐域性植被、植被代用指标局限性及植被记录空间分布不合理等难点,需要更多深入工作,以期获得更丰富、更准确的信息以及更细致的植物类型划分,为定量分析和数值模拟提供可靠的数据资料,综合评判冰期低海平面时期出露陆架植被变化对全球碳循环和气候环境格局变化的影响和响应。
5 总结
末次冰期巽他陆架上的植被状况对于陆地碳储库有着重要影响,但目前还存在争论。本文对巽他陆架区域末次冰期以来已发表的碳同位素和植物孢粉记录进行了收集和整理,分析认为末次冰期巽他陆架上的热带雨林具有带状分布和垂直结构演化两个特征。
(1)与全新世相比,冰期热带雨林分布范围向赤道方向收缩,呈现不规则的带状分布。远离赤道的外围区域草本植物扩张,但不足以替代赤道区域的热带雨林,不太可能出现“热带稀树草原廊道”。ITCZ的向南移动和热带太平洋的El Niño-like状态导致的巽他陆架区域整体降水减少是造成这种现象的重要原因。
(2)山地在巽他陆架末次冰期植被垂直分布结
