外源物质处理对大果榉扦插苗抗寒性的影响
2021-08-16白京凡白晋华郭红彦
霍 静,白京凡,乔 虹,白晋华,郭红彦
(山西农业大学 林学院,北方功能油料树种培育与研发山西省重点实验室,山西 太谷 030801)
大果榉(Zelkovasinica)又名小叶榉、抱榉,为榆科榉属的一种落叶乔木[1],是第三世纪温带森林孑遗物种[2],也是山西省珍惜的乡土树种[3]。大果榉较喜光、喜温暖湿润气候,耐干旱、贫瘠,常生长在石灰岩发育的山地钙质土壤上,在我国主要分布于甘肃、陕西、四川北部、湖北西北部、河南、山西南部和河北等地[4-5]。在大果榉的栽培繁育过程中,低温是限制其生长与分布的主要环境因素[3]。
低温作为重要的非生物胁迫因子,对植物生长发育有重要影响,会造成植物新陈代谢等生理过程的紊乱,使生物膜和细胞结构遭受损伤[6-8]。关于林木抗寒性的研究已有诸多报道,在杨树、桢楠、梨、核桃等植物[9-12]抗寒性的研究中发现,植物的抗寒性与相对电导率、MDA和可溶性蛋白含量等指标的变化具有一定相关性。研究表明,喷施外源物质能有效影响和控制植物的生长发育及对低温胁迫的适应性[13],低温来临前施用一定浓度的生长调节剂、CaCl2均可提高植物的抗寒性,缓解植物的低温伤害[14]。PP333是一种高效、低毒的植物生长调节剂,能延缓生长、控制形态建成、提高植物抗性[15-16];Ca2+不仅是植物生长发育所必需的矿质元素,且参与植物生长发育的调节过程,在细胞内逆境信号的转导和植物对逆境的调节适应过程中发挥着重要作用[17]。目前,有关榉属植物抗寒性方面的研究引起人们的重视,经外源物质处理的大果榉幼苗对低温的生理响应尚未见报道。本研究根据大果榉栽培过程中遇到的实际问题,探索不同浓度PP333与CaCl2处理对大果榉抗寒性的影响,以期为提高大果榉幼苗抗寒性提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为山西农业大学林学院苗圃中扦插的当年生大果榉盆栽苗。于2018年3月底,选取生长基本一致、无病虫害、直径8 mm的大果榉枝条剪成10 cm的插穗,以每钵一根的标准,分别插于装有珍珠岩与草炭(比例为1∶1)、规格为10 cm×10 cm的营养钵中,日常管理使温室内保持70%的湿度。待生根后将整株幼苗移栽到21 cm×21 cm的营养钵内,栽培基质为田土和营养土1∶1混合,基质约占营养钵体积的80%,每盆1株,置于室外,进行常规的浇水养护,缓苗至生长健壮后喷施外源物质、进行低温胁迫。
1.2 试验设计
试验选用PP333和CaCl2处理大果榉幼苗,设置对照(清水)、PP333(0.1%、0.15%)和CaCl2(20、25 mmol/L)共5个处理。于2018年8月15日早上8:00,将大果榉盆栽苗分成5组,每个处理20株,进行叶面喷施,以叶面滴水为宜,每隔5 d喷1次,共处理3次。
于2018年10月中旬,随机选取不同处理苗木的1年生枝条,每个处理选6株,每株采3个侧枝,共90个枝条;将采集的枝条用湿报纸包裹带回实验室进行人工模拟低温处理,设置4、-10、-15℃ 3个温度梯度,各温度均放不同处理采自不同植株的枝条6个,每个温度30个枝条,低温处理12 h。在4℃环境下解冻,分别取每个处理不同温度条件下的枝条基部起第4~6位的叶片混匀、用于生理指标的测定,每个指标均重复测定3次。
2018年11月至2019年2月,统计自然越冬过程中不同处理大果榉枝条的冻害级别,每月1次,共4次。
1.3 指标测定
光合色素含量采用乙醇浸提比色法[18],可溶性蛋白含量按照G-250考马斯亮蓝法测定[19],相对电导率采用电导法测定[20],丙二醛含量采用硫代巴比妥酸法测定[19]。
参照李瑞雪[21]田间冻害调查法的标准对冬季不同处理大果榉扦插苗的冻害等级进行统计,并在此基础上稍有改动,共分为6级,依次为:0级,枝条无受冻现象;1级,枝条受冻比例<20%;2级,枝条受冻比例<40%;3级,枝条受冻<60%;4级,枝条受冻比例<80%;5级,几乎所有枝条或整株幼苗冻死。
1.4 抗寒性综合评价方法
参照玉苏甫·阿不力提甫等[22]的方法,运用相关性分析和隶属函数法对大果榉的抗寒性进行综合分析。隶属函数值的计算方法如下:
如果指标与抗寒性为正相关,Zij=(Xij-Ximin)/(Ximax-Ximin)
如果指标与抗寒性为负相关,Zij= 1-(Xij-Ximin)/(Ximax-Ximin)
式中,Zij为第i个处理第j个指标的抗寒隶属函数值;Xij为第i个处理第j个指标的测定值;Ximin和Ximax分别为各指标的最小值和最大值;后求出各处理的平均隶属度进行比较。
1.5 数据处理
采用SPSS17.0、Excel2003进行数据处理和作图,结合LSD与Duncan法进行方差分析与多重比较,各指标间采用Spearman系数进行相关性分析。
2 结果与分析
2.1 对低温胁迫下大果榉光合色素的影响
不同处理的大果榉幼苗光合色素含量随温度降低均呈现出逐步下降的趋势;与对照相比,喷施外源物质可不同程度减缓低温胁迫下大果榉光合色素的降解。在相同温度条件下,经P1、P2、C1、C2处理的大果榉叶片中叶绿素a含量、叶绿素b含量、叶绿素总量、类胡萝卜素含量均高于对照;P2处理的大果榉叶绿素a、叶绿素总量与对照相比差异显著;C1、C2处理后的4个色素含量均显著高于对照,且20 mmol/L CaCl2时达到峰值,处理效果最好。-15℃时,20 mmol/L CaCl2处理的大果榉幼苗光合色素含量分别比对照高8.15、1.64、9.79、1.36 mg/g(图1)。
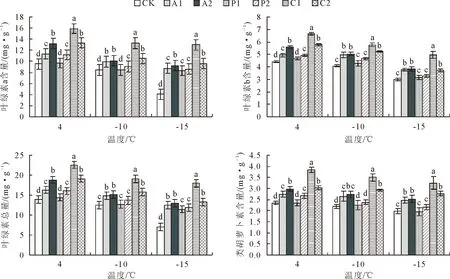
注:不同小写字母表示相同温度不同处理间差异显著(P<0.05),不同大写字母表示相同处理不同温度间差异显著(P<0.05)。CK、P1、P2、C1、C2分别表示对照,0.1% PP333,0.15% PP333,20 mmol/L CaCl2,25 mmol/L CaCl2。下同。
2.2 对低温胁迫下大果榉可溶性蛋白含量的影响
相同低温条件下,与对照相比,不同外源物质处理均可促进大果榉叶片中可溶性蛋白含量的积累,延缓低温胁迫下可溶性蛋白含量的减少。PP333处理后的可溶性蛋白含量随浓度的增加而增加,CaCl2处理的大果榉叶片中可溶性蛋白含量随浓度的升高而降低,且CaCl2处理后的可溶性蛋白含量均高于PP333;其中,C1、C2在各温度下的可溶性蛋白含量均显著高于对照。不同低温环境下,C1处理后的大果榉幼苗可溶性蛋白含量比对照分别显著增加7.34、8.59、6.19 mg/g,C2处理后的分别显著提高5.26、5.19、5.07 mg/g,以20 mmol/L CaCl2促进效果最好(图2)。

图2 不同处理对低温胁迫下大果榉可溶性蛋白含量的影响Fig.2 Effects of different treatments on soluble protein content of Z.sinica
2.3 对低温胁迫下大果榉相对电导率的影响
研究结果表明(图3),各处理对不同低温下大果榉叶片的相对电导率均有显著影响,与 CK相比,喷施外源物质不同程度降低了大果榉叶片的相对电导率、减小了随温度降低各处理大果榉幼苗相对电导率的上升幅度。在相同温度条件下,P1、P2处理的大果榉相对电导率均低于对照、但差异不显著,C1、C2处理后大果榉的相对电导率均显著低于对照。2种外源物质分别以P2、C1处理效果较好;C1处理可显著减缓相对电导率的增加,且对-15℃胁迫后大果榉幼苗相对电导率的影响最大,与CK相比显著降低35.75%。
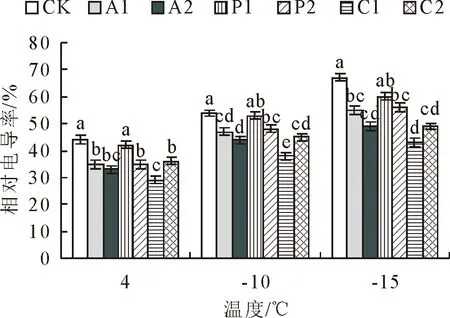
图3 不同处理对低温胁迫下大果榉相对电导率的影响Fig.3 Effects of different treatments on relative conductivity of Z.sinica
2.4 对低温胁迫下大果榉丙二醛含量的影响
不同低温环境下,经外源物质处理的大果榉幼苗MDA含量均低于对照,各处理在-10℃时的MDA含量与对照相比均差异显著,分别降低30.69%,34.38%,60.09%,36.30%。PP333处理的大果榉MDA含量随浓度增加而降低,P2处理的MDA含量在-10℃、-15℃时与CK相比差异显著;经C1、C2处理后的大果榉MDA含量在不同温度下均显著低于对照。不同处理大果榉的MDA含量均随温度降低而不断上升,从-10℃~-15℃,PP333处理与对照组的大果榉幼苗MDA含量均显著升高,C1、C2处理的大果榉MDA含量升高不显著,且C1处理后的大果榉MDA含量在不同温度下均最低、随温度降低其上升幅度最小(图4)。

图4 不同处理对低温胁迫下大果榉丙二醛含量的影响Fig.4 Effects of different treatments on MDA content of Z.sinica
2.5 不同处理对大果榉幼苗越冬过程中冻害程度的影响
在自然越冬过程中,大果榉幼苗有不同程度的冻害现象。与对照相比(表1),喷施外源物质均可不同程度减缓大果榉幼苗的受冻害情况。2018年11月,不同处理大果榉幼苗均没有发生冻害现象。2018年12月,部分处理的大果榉幼苗枝条开始出现冻害,且随时间推移,受冻害程度不断加深。到2019年2月,对照组大果榉幼苗受冻害最严重、冻害级别达5级;经P1和P2处理的大果榉幼苗冻害级别分别为4级、3级;经CaCl2处理的大果榉幼苗耐寒力较好,喷施20 mmol/L CaCl2的大果榉受冻害程度最低,冻害级别为1级。
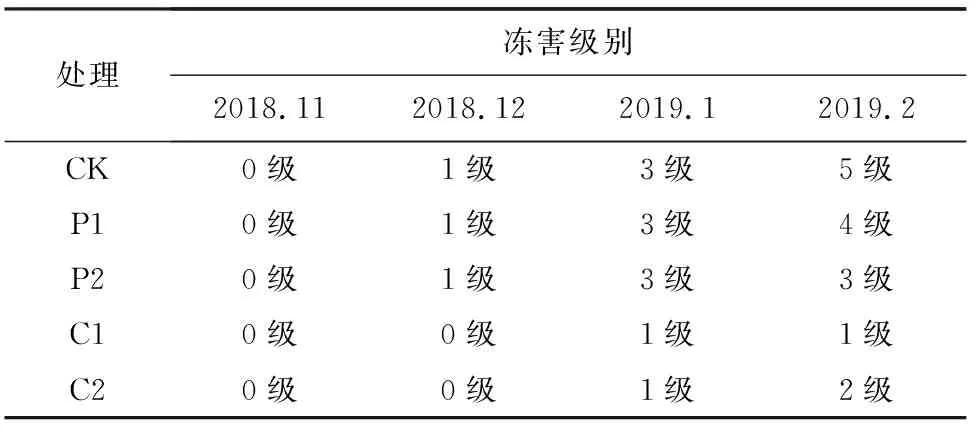
表1 自然越冬过程中各处理大果榉的冻害级别Table 1 Freezing damage levels of Z.sinica during the natural over wintering process
2.6 大果榉各抗寒指标之间的相关性
对不同处理大果榉幼苗各生理指标与冻害级别之间的相关性分析结果(表2)表明,光合色素、可溶性蛋白含量与冻害级别呈极显著负相关,其值越高,冻害程度越低,抗寒性越强;相对电导率、MDA含量与冻害级别呈极显著正相关,其值越高,冻害程度越严重,抗寒性越弱。

表2 大果榉各抗寒指标间的相关系数Table 2 Correlation coefficients among cold resistance indexes of Z.sinica
2.7 不同处理对大果榉幼苗抗寒性影响的综合评价
对不同浓度PP333、CaCl2处理大果榉幼苗的各项生理指标进行隶属函数分析,综合评价各处理对低温胁迫下大果榉抗寒性的影响。外源物质对大果榉抗寒性影响的综合评价值均高于对照(表3),低温下各处理的隶属函数值排序依次为C1>C2>P2>P1>CK,与自然越冬后不同处理大果榉冻害级别所反映出的结果相一致,以20 mmol/L CaCl2缓解大果榉低温伤害的效果最好。

表3 不同处理对大果榉抗寒性影响的综合评价Table 3 Comprehensive evaluation of the effects of different treatments on cold resistance of Z.sinica
3 讨论
3.1 对低温胁迫下大果榉幼苗光合色素的保护作用
光合作用为植物生长、发育提供所需的物质和能量,是植物生长发育的基础。低温对植物光合色素产生、光系统活性及光合能量代谢等一系列过程都有不同程度的影响[23]。具体表现为,低温胁迫会抑制光合作用反应中酶活性的水平,破坏叶绿体双层膜结构,引起光合色素降解[24],导致光合能力下降,进而影响植物的其他生理代谢过程[13-14,25]。本研究发现,在低温胁迫下,经PP333、CaCl2处理后的大果榉幼苗叶片叶绿素和类胡萝卜素含量有所提高,说明适宜浓度的外源PP333和CaCl2对大果榉叶绿素的结构和功能具有一定的保护作用,在遭遇低温胁迫时仍能够维持其结构与功能、发挥正常的生理作用,这与于永畅等[26]对国兰、孙兵等[27]对桢楠和闽楠抗寒性的研究结果相一致。可能原因是外源物质起到调节植物细胞的作用,最终减少低温对叶绿体的破坏,在促进光合色素合成的同时,保护光合色素的结构,使细胞内光合色素的含量维持在一个较高水平。
3.2 对低温胁迫下大果榉幼苗的渗透调节作用
可溶性蛋白是广泛存在于植物细胞液中能够调控渗透压的物质之一[28],在植物细胞中含量的高低与植物抗寒性之间有着密切的关系[14]。研究表明,喷施适当浓度的外源物质可诱导植物体内产生并积累一定量的可溶性蛋白,进而增加植物细胞渗透压、减缓细胞内水分损失,以增强植株对低温的抵抗力[14,29]。可溶性蛋白含量的上升还能降低植物细胞液的冰点,从而提高植物对低温的耐受力[27]。本研究发现,低温胁迫前用PP333、CaCl2处理后的大果榉幼苗,其可溶性蛋白含量均显著升高,说明经外源物质处理的大果榉幼苗在低温胁迫下可通过自身的调节能力而自发产生某种对低温胁迫的适应机制,积累一定的蛋白含量,缓解低温胁迫带来的伤害,这与梁永富等[30]关于生长调节剂对低温胁迫下酸橙影响的研究结果相一致,且可溶性蛋白含量与大果榉抗寒性呈正相关,其含量越高,抗寒性越强。
3.3 对低温胁迫下大果榉幼苗细胞膜的保护作用
在低温环境下,植物敏感和最易伤害的部位是细胞膜[31]。植物处于逆境时,其体内活性氧代谢失调致使细胞膜被破坏、透性增加,细胞内溶质向胞间渗漏,从而导致植物的电导率增加[13],严重时还会发生膜脂过氧化作用。相对电导率是反映细胞膜透性大小的指标,其值越大,说明受胁迫后细胞膜的透性越大[14,32];MDA是反映生物膜过氧化程度的指标,其含量多少可以体现细胞膜受伤害的程度[6,32],MDA含量积累越多,膜系统受伤害越严重,MDA含量与植物抗寒性呈负相关性[22,33],本研究中MDA和相对电导率与大果榉幼苗冻害级别呈极显著正相关的结果与此相一致。喷施PP333、CaCl2可显著减缓大果榉幼苗在低温胁迫下叶片中MDA含量和相对电导率的上升,且随温度降低CaCl2处理的大果榉幼苗MDA含量升高不显著,说明经CaCl2处理后大果榉幼苗的细胞膜系统在低温胁迫下受害程度较低。总之,叶面喷施外源物质可以减少活性氧的产生,降低膜脂过氧化的程度,有效抵御低温对大果榉幼苗的伤害,从而提高了大果榉幼苗的抗寒性,这与王丽特等[24]关于4种化学调控剂对烟草幼苗耐冷性、周伟权等[34]对库尔勒香梨抗寒性的研究结果相类似。
3.4 大果榉幼苗抗寒性的综合评价
植物各抗寒性指标间有一定的相关性[22,33,35]。本研究结果表明,低温胁迫下大果榉幼苗的冻害级别与其相对电导率、MDA含量呈极显著正相关,与光合色素、可溶性蛋白含量呈极显著负相关;在低温胁迫下,大果榉幼苗相对电导率、MDA含量越低,叶绿素、可溶性蛋白含量越高,其受冻害程度越低,抗寒性越强。采用单一指标评价植物的抗寒性不具有代表性,应采用多个指标对其抗寒能力进行综合评价[22,34-35]。本研究运用模糊数学的隶属函数法能较全面地反映不同处理大果榉幼苗的抗寒性强弱,平均隶属度越大,表明抗寒能力越强,反之抗寒能力越弱[21-22]。结合自然越冬后各处理冻害级别和抗寒性综合评价结果可知,经PP333、CaCl2处理后的大果榉幼苗在低温胁迫条件下其抗寒能力均有所提高,以20 mmol/L CaCl2处理的抗寒性效果最好,可为今后大果榉生产提供可靠的依据。
4 结论
低温是影响大果榉生长的重要环境因子之一,在大果榉栽培过程中应采取相应抗寒措施降低大果榉树苗的低温伤害,以确保其正常越冬。本研究发现,在低温来临之前,叶面喷施PP333、CaCl2均能够减缓低温胁迫下大果榉幼苗光合色素的下降趋势,促进可溶性蛋白的积累,降低细胞膜的相对电导率和MDA含量,并最终提高低温胁迫下大果榉幼苗的抗寒性。不同处理对大果榉抗寒性影响的隶属函数综合评价结果与自然越冬后冻害级别的统计结果相一致,以20 mmol/L CaCl2处理的大果榉幼苗抗寒效果最好。
