植株结构对不同虫态蠋蝽搜索和捕食斜纹夜蛾幼虫行为的影响
2021-08-11李发倩李保平
李发倩,李保平,孟 玲
(南京农业大学植物保护学院/农作物生物灾害综合治理教育部重点实验室,南京 210095)
自Price等[1]的经典论述后,植物与更高营养层级的捕食性天敌昆虫的化学生态学研究得到了广泛而深刻的关注。植物化学特征可影响捕食性和寄生性天敌的搜寻和捕食(寄生)行为,进而影响天敌的种群数量动态和群落食物网结构与功能[2]。与此相比,对植物物理特性如何影响天敌昆虫行为的认识,尚缺乏足够的实据。Andow[3]曾提出影响节肢动物搜寻行为的三个植株物理特征:(1)植株大小或体表面积;(2)植株结构的异质性,如种穗、花、蜜腺和叶片具有不同的表皮(如被毛情况);(3)植株结构复杂性,又称“连通性”(connectedness),代表单位投影面积下枝叶数量。理论模型预测植株结构可影响捕食性昆虫和螨类的捕食效率[4-5],该模型预测得到了若干实证研究的支持,叶面的被毛、蜡质层和坑窝、枝叶的密度、方向和重叠度、植株体形结构的复杂性等植株微生境特征,可影响捕食性节肢动物的行为和生态学特征[6]。例如,对番茄Solanumlycopersicum与天敌互作的研究发现,番茄叶面被毛的物理结构可影响寄生性和捕食性天敌昆虫的生态学特征[7];大豆Glycinemax植株结构复杂性可影响异色瓢虫Harmoniaaxyridis与方斑瓢虫Propyleaquatuordecimpunctata之间的集团内捕食行为的发生[8];菜豆Phaseolusvulgaris植株的连通性可影响智利小智绥满Phytoseiuluspersimilis对二斑叶螨Tetranychusurticae的捕食效率[9];普通草蛉Chrysoperlacarnea在叶片结构不同的沙生冰草Agropyrondesertorum和长毛落芒草Oryzopsishymenoides上对麦双尾蚜Diuraphisnoxia的捕食量不同[10]。迄今,对活动性较强的捕食性节肢动物对防卫能力较弱的猎物的捕食关注较多,例如,食蚜天敌(瓢虫、草蛉、小花蝽等)对小型害虫蚜虫、粉虱和蓟马的捕食,这类天敌的处理猎物时间很短,可在短时间内捕食较多的猎物;而对那些活动性较弱的捕食性蝽如何搜寻和捕食防卫能力较强的大型蛾类幼虫的关注,明显较少,例如,某些行动迟缓的蝽科Pentatomidae捕食者对大体型鳞翅目幼虫的捕食,这类捕食者处理猎物时间很长,长时间内只捕食1头猎物。
捕食性蝽种类繁多,攻击性强、捕食量大,捕食效率高,成虫和若虫均可捕食多种植食性昆虫,在控制植食性昆虫种群数量中发挥着重要作用[11-13]。近年来,蠋蝽Armachinensis在害虫生物防治中的作用日益受到重视,对多种害虫具有较强的捕食作用,包括:榆紫叶甲Ambrostomaquadriimopressum[14,15]、黄刺蛾Cnidocampaflavescens[15]、美国白蛾Hyphantriacunea[16]、草地贪夜蛾Spodopterafrugiperda[17]和斜纹夜蛾S.litura[18]等多种农林作物上的重要的本土和外来入侵害虫[19]。蠋蝽活动性较弱[17],蛾类幼虫尤其高龄幼虫,对天敌昆虫的攻击往往表现出较强的形态、生理或行为防卫反应[20-22]。迄今,对植株结构复杂性如何影响捕食性昆虫的知识,多来自于对活跃的捕食者与防卫能力低的猎物的研究[4,5],而对植物结构如何影响不活跃的捕食性天敌与大体型猎物的互作行为,了解很少。
本研究以蠋蝽和斜纹夜蛾为捕食者-猎物模式生物,旨在明确植株结构复杂性如何影响蠋蝽对斜纹夜蛾幼虫的搜寻和捕食行为,以及该影响是否随蠋蝽虫龄(态)不同而变化。本研究旨在揭示不活泼的捕食者对防卫性较强的猎物的捕食行为是否受植物结构的影响。
1 材料与方法
1.1 供试材料
蠋蝽由中国农科院植物保护研究所生物防治研究室提供,以斜纹夜蛾幼虫为猎物在养虫室(26 ℃±1 ℃,RH 60%±10%,光周期16L∶8D)建立饲养种群。蠋蝽若虫饲养于圆形PP材质塑料盒(上口径15 cm、底径11 cm、高7 cm,盒盖有用纱网封上透气用的开口)中,提供用湿棉球缠绕叶柄的新鲜榔榆Ulmusparvifolia树叶作为辅助食物供蠋蝽吸食(1龄若虫主要吸食榆树汁液,2龄若虫开始取食斜纹夜蛾幼虫)。选取蠋蝽蜕皮后24 h内的3龄、5龄若虫以及羽化24 h内的雌、雄成虫作为试虫。
斜纹夜蛾幼虫用人工饲料[23]饲养于养虫室中,在圆形食品塑料盒(直径20 cm,高10 cm)中群体饲喂;化蛹后收集蛹;羽化的成虫置于方形纱笼(40 cm×40 cm×40 cm)中产卵。选取蜕皮后24 h内的4龄幼虫供试。
供试大豆品种为“南农1138-2”,由南京农业大学国家大豆改良研究中心提供。用营养土(镇江兴农有机肥有限公司生产)装盆(上口径12 cm、底径8 cm、高10 cm)栽种大豆,在室外生长约65 d (株高约50 cm)待用。
1.2 试验设计
参考大田蚕豆种植中的株距范围,设置大豆植株结构复杂性低、高两个水平,分别简称为“简单植株”和“复杂植株”。简单植株在1个花盆(直径12 cm)中栽种1株大豆,高约50 cm,生有8个枝条(每枝生3片叶);复杂植株在1个花盆(直径12 cm)中栽种2株大豆,每株50 cm高并生有8个枝条,2株紧靠使枝叶部分重叠。设置蠋蝽虫龄(态)4个水平:3龄若虫、5龄若虫、雌成虫和雄成虫,均选蜕皮24 h内的若虫和羽化24 h内的成虫,释放前饥饿48 h(只提供保鲜榆树叶)。分别在大豆植株顶部4个枝条(按顺序编号为1、2、3、4)的叶片上分别接1头斜纹夜蛾幼虫(放在3片叶中间),待幼虫稳定(约10 min)后释放1头供试蠋蝽,放在距离最下部有虫枝条分叉处约5 cm主茎干上。蠋蝽沿主茎向上搜索,连续观察记录其2 h内的行为(表1),因为蠋蝽在2 h内往往可完成一次攻击猎物(麻痹猎物)行动[24]。每盆植物、每头蠋蝽只用1次;重复30次。

表1 观测的蠋蝽搜寻及捕食行为特征描述Table 1 A description of recorded searching and predation behaviors of A.chinensis
1.3 数据统计与分析
用多项logit模型(multinomial logit model)拟合蠋蝽对4个有虫枝条的首次选择,运行ordinal功能包分析[25];用广义线性模型(误差为Gamma分布,连接函数为倒数)拟合行为的时间特征变量(分析静伏时间时设置误差分布为Poisson分布型,连接函数为自然对数)。用Cox比例风险回归模型拟合蠋蝽潜伏期可估计捕食者对猎物的捕食风险(瞬时概率),该模型公式如下[26]:
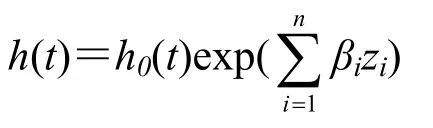
h(t) 为时间t时的概率,此处代表猎物被发现的风险大小;h0(t)为基准概率,类似多重回归模型中的截距(不同之处在于它在该模型随时间而变,而在多重回归模型中为常量),此处代表在对照下对猎物的捕食风险大小;时间t为从某一时刻开始直到事件发生的间隔期,此处代表蠋蝽的潜伏期;β为系数,z为协变量(影响因素),此处代表所检验的影响因素——植株复杂性和蠋蝽虫龄(态)对猎物被发现的风险大小的影响。检验协变量影响系数的指数函数为风险比率(odds ratio),风险比率>1说明与对照相比风险增大;风险比率<1,说明风险降低;运行survival功能包[27]拟合Cox模型。在所有模型中,用对数似然比测验确定植株结构复杂性和蠋蝽虫龄(态)的影响,运行car功能包分析[28];在比较蠋蝽虫龄(态)处理间差异时用Tukey法矫正差异显著性概率(α=0.05),运行emmeans功能包分析[29]。数据分析用R(3.5.0版)统计软件[30]。
2 结果与分析
2.1 蠋蝽对有猎物枝条的选择性
蠋蝽虫态(龄)(χ2=5.60,P=0.78)、植株复杂性(χ2=1.06,P=0.79)以及两因素互作(χ2=14.19,P=0.12)均不影响蠋蝽对有虫枝条的首次选择偏好;蠋蝽对有猎物枝条的选择比例在虫龄(态)之间、植物结构之间存在很大差异(5%~30%),且没有一致的格局(图1)。

图1 不同虫龄(态)蠋蝽在植株结构简单和复杂的大豆植株上对4个有猎物枝条的首选比例Fig.1 Proportion of choices for the four different branches with prey on simple and complex soybean plants by A.chinensis at different life stages
2.2 猎物被捕食风险
用Cox比例风险模型拟合潜伏期估计猎物被捕食风险(瞬时概率)的结果表明,植株复杂性(χ2=30.61,P<0.001)和蠋蝽虫态(龄)(χ2=8.69,P=0.03)显著影响斜纹夜蛾幼虫被蠋蝽捕食的风险,但两因素之间无互作影响(χ2=1.41,P=0.70)。在简单植株上的被捕食风险是在复杂植株上的2.4倍(Z=5.51,P<0.001)(图2A);被雌躅蝽与雄躅蝽成虫捕食的风险没有差异(Z=0.60,P=0.55);与雌成虫相比,被3龄若虫(Z=2.51,P=0.012)和5龄若虫(Z=2.84,P=0.004)捕食的风险分别增大1.7和1.9倍;与雄成虫相比,被3龄(Z=1.83,P=0.26)或5龄(Z=2.24,P=0.11)若虫捕食的风险相似;被3龄与5龄若虫捕食的风险相似(Z=0.18,P=0.93)(图2B)。

图2 斜纹夜蛾幼虫在结构复杂性不同的植株上(A)被不同虫龄(态)蠋蝽(B)捕食的累积风险Fig.2 Cumulative predation risk to S.littura larvae as a function of soybean plant complexity (A) and stinkbug life stage (B) by A.chinensis
2.3 蠋蝽搜索用时
植株复杂性(χ2=51.06,P<0.001)和蠋蝽虫龄(态)(χ2=10.00,P=0.02)分别显著影响搜索用时,但两因素互作无显著影响(χ2=2.17,P=0.54)(图3)。在复杂植株上的搜索用时是在简单植株上的1.5倍(Z=6.93,P<0.001);3龄若虫的搜索用时是5龄若虫的1.3倍(Z=3.12,P=0.01),而两者与雌、雄成虫的搜索用时均无差异。

图3 不同虫龄(态)蠋蝽在简单和复杂大豆植株上的搜索用时(均值±标准误)Fig.3 Searching time on structurally simple and complex soybean plants by A.chinensis at different stages (mean±SE)
2.4 蠋蝽的静伏用时
蠋蝽虫龄(态)(χ2=52.31,P<0.001)显著影响静伏用时,植株复杂性(χ2=0.31,P=0.58)没有显著影响,两因素互作(χ2=3.25,P=0.36)也无显著影响(图4)。雌成虫的静伏用时是3龄若虫的1.8倍(Z=6.42,P<0.001),是5龄若虫的1.6倍(Z=5.31,P<0.001), 是雄成虫的1.3倍(Z=2.66,P=0.04);雄成虫的静伏用时是3龄若虫的1.7倍(Z=3.88,P=0.001),是5龄若虫的1.3倍(Z=2.72,P=0.03);3龄若虫与5龄若虫的静伏用时无显著差异(Z=1.19,P=0.63)。

图4 不同虫龄(态)蠋蝽在简单和复杂大豆植株上的静伏用时(均值±标准误)Fig.4 Resting time on structurally simple and complex soybean plants by A.chinensis at different stages (mean±SE)
2.5 蠋蝽的麻痹猎物用时
蠋蝽虫龄(态)(χ2=266.26,P<0.001)显著影响麻痹猎物用时,但植株复杂性(χ2=0.02,P=0.90)以及两因素互作(χ2=3.48,P=0.32)没有显著影响(图5)。3龄若虫麻痹猎物用时是雌成虫的2.2倍(Z=12.45,P<0.001),是雄成虫的2.1倍(Z=11.93,P<0.001),是5龄若虫的1.7倍(Z=9.26,P<0.001);5龄若虫麻痹猎物用时是雌成虫的1.3倍(Z=4.04,P<0.001),是雄成虫的1.2倍(Z=3.11,P=0.005);雌、雄成虫之间的麻痹猎物用时无显著差异(Z=0.72,P=0.89)。

图5 蠋蝽不同虫龄(态)在简单和复杂大豆植株上的麻痹猎物用时(均值±标准误)Fig.5 Prey-paralyzing time on structurally simple and complex soybean plants by A.chinensis at different stages (mean±SE)
3 讨论
本研究结果说明,植株结构复杂可降低斜纹夜蛾高龄幼虫被蠋蝽不同虫龄(态)的捕食风险,因为蠋蝽在较复杂的植株上花费的搜寻猎物用时更长;而处理猎物用时不受植株结构的影响,只随虫龄(态)而变化。
对许多活跃的捕食性昆虫与防卫能力较弱的猎物的研究说明,植物密度可以影响植物小生境的气候(如温、湿度等),从而影响昆虫的生境选择[31]。同时,植物密度、多样性可干扰昆虫的视觉反应[32],寄主植物叶片和叶毛不仅能为植食性害虫提供藏身之所,还能刺激或抑制植食性昆虫的取食,同时亦能阻碍昆虫天敌的搜寻行为,降低昆虫天敌的寻找效率[33,34]。植物结构影响捕食性昆虫的搜寻和捕食,已在其他捕食性昆虫类群中观察到[6],本研究提供了新的捕食性蝽证据,说明植物结构复杂性也可影响不活跃的捕食性昆虫对防卫能力较强的大型蛾类幼虫的搜寻行为。
但植株结构对蠋蝽搜寻和捕食行为的影响也有其特征:蠋蝽在复杂植株上的搜索用时延长,而静伏用时和麻痹猎物用时不受植株结构的影响,仅随蠋蝽虫龄(态)不同而变化。这说明植株结构复杂可降低蠋蝽的搜寻效率,其原因可能是猎物在复杂植株上更易于躲避蠋蝽搜索。但蠋蝽攻击猎物行为的时间特征参数不受植株复杂性的影响,而比较活跃的瓢虫[8]、捕食螨[9]、草蛉[10]以及寄生蜂[30]等的捕食(寄生)行为往往受植株复杂性的影响。这一差异的原因可能在于,活跃的天敌昆虫对植株的连通性比不活跃的昆虫更敏感,因为前者仅需很短的时间就完成一次捕食,然后立刻转入对下一猎物的搜索;而后者需要相对较长的时间方完成一次捕食,攻击不同猎物的时间间隔也很长[18]。如果植株结构的连通性影响捕食者高频次反复搜索猎物,则植株结构复杂性对活跃性较强的捕食者的影响必然大于对惰性较强的捕食者的影响。
斜纹夜蛾幼虫被蠋蝽若虫捕食的风险大于被蠋蝽成虫,说明蠋蝽若虫搜寻猎物的积极性高于成虫。此前的功能反应研究也发现,5龄蠋蝽若虫对草地贪夜蛾幼虫的攻击率大于成虫[17,35]。据此推测,若虫捕食控害作用可能大于成虫。
致谢:感谢中国农科院植保所生物防治研究室陈红印先生提供蠋蝽虫源。
