2,4-表油菜素内酯对烟苗幼茎生长及相关基因表达的影响
2021-08-09张娟吴晓颖许娜杜沙沙张彦马兴华
张娟 吴晓颖 许娜 杜沙沙 张彦 马兴华
摘 要:为研究油菜素内酯(Brassinolide, BR)对烟草主茎生长的影响,利用外源2, 4-表油菜素内酯(2, 4-Epibrassinolide, EBR),设置0.5×10-7 mol/L(T1)、0.5×10-5mol/L(T2)两个浓度,以蒸馏水(CK)为对照对烟草幼苗进行喷施。分析了不同处理的烟草幼苗株高,节距及解剖结构特征,检测了茎中与细胞分裂和细胞大小调控相关基因以及与油菜素内酯(BR)、生长素(IAA)和赤霉素(GA)合成相关基因的表达量。结果表明,喷施EBR对烟草幼苗节距和株高有显著促进作用,并随处理浓度升高,促进作用增强。观察石蜡切片发现EBR处理后茎皮层薄壁细胞数目增多,单细胞面积减小。EBR处理后,细胞周期调控基因NtCYCD3表达上调,细胞大小调控基因NtARF6、NtARF16表达下调;BR信号受体基因NtBRI1,NtBIN2表达上调,BR转录因子NtBES1T表达下调;BR、IAA和GA的关键生物合成基因NtDWF4、NtYUCCA8、NtGA3ox-2表达量均上调。说明喷施EBR可促进内源BR、IAA和GA的合成基因表达,通过促进节间细胞分裂、抑制细胞大小促进烟草幼苗茎的生长。
关键词:2, 4-表油菜素内酯;烟草;茎伸长生长;解剖结构;基因表达量
Abstract: To study the effect of brassinolide (BR) on tobacco stem growth and elongation, the tobacco seedlings were treated with 2,4-epibrassinolide (EBR) of two different concentrations 0.5×10-7 mol/L (T1), 0.5×10-5 mol/L (T2) and distilled water was used as control (CK). Plant height, internode distance, and stem anatomical structure were analyzed. Genes related to cell division and cell size regulation, and genes related to biosynthesis of BR, auxin (IAA), and gibberellin (GA) were also analyzed. The results showed that the application of EBR increased the seedling pitch and plant height, the promotion effect on stem growth was enhanced with increment of EBR concentrations. Stem anatomical study revealed an increase in the number of parenchyma cells and a decrease in its area after EBR treatment. EBR upregulated cell cycle-related gene NtCYCD3 while downregulated NtARF6 and NtARF16 genes that are related to cell size. Similarly, BR signaling receptor genes NtBRI1 and NtBIN2 were upregulated while the BR transcription factor NtBES1T was downregulated in response to EBR application. The expression of key biosynthesis genes NtDWF4, NtYUCCA8, and NtGA3ox-2 of BR, IAA, and GA, respectively, were all upregulated. The results showed that the application of EBR promoted the expression of endogenous BR, IAA and GA biosynthesis genes, and helped in the growth and elongation of tobacco seedling stem by promoting the division of internode cells and inhibiting cell expansion.
Keywords: 2, 4-epibrassinolide; Nicotiana tabacum; stem elongation; anatomical structure; gene expression;
莖是植物的三大营养器官之一,具有支持、运输、储存、繁殖和光合的功能,在植物的生长发育过程中起着不可替代的作用[1]。株高是指植株主茎基部到顶部之间的距离,由植株的茎节数目和节间长度决定,是农作物重要的生物学性状之一,与作物经济产量密切相关。适当的株高能抗倒伏,还有利于CO2的分布,提高光能利用率[2]。
植物株高的增长包括主茎节数的增多以及各节间的伸长[3]。细胞的分裂和伸长是植物茎伸长生长的主要原因,这一过程受多种内外因素的调节和控制。油菜素内酯(BR)是一种甾醇类植物激素,具有促进生长、增加植物抗逆性、延缓衰老、促进细胞再分化等功能,但最突出的生理作用就是促进植物的生长[4]。BR对植物生长的促进是通过促进细胞分裂和伸长的作用实现的,并且这一促进作用具有浓度效应,多表现为低浓度促进高浓度抑制[5-6]。EBR是广泛使用的一种人工合成油菜素内酯,具有提高烟草抗旱性[7],促进十字花科植物幼苗和豌豆上胚轴细胞伸长[8],诱导水稻节间伸长[9]的功能。
株高是与烤烟产值关系最密切的因素之一,烟草株高通过影响烟株有效叶数、叶长、叶宽而间接影响烟叶产量和品质[10-11]。而目前关于施用植物生长调节剂对烟草茎伸长生长的研究报道尚少;施用BR如何影响烟草主茎伸长,是否会影响GA、IAA含量及其合成基因的表达等尚未见报道。因此本研究通过喷施不同浓度EBR,从生物学性状,茎细胞解剖学特征,控制细胞分裂和大小的关键基因、BR信号通路基因和内源激素合成相关基因表达量等方面分析其对烟草茎伸长生长的影响,进一步阐述外源植物生长调节剂在调控烟草茎生长中的作用。
1 材料与方法
1.1 供试材料
供试品种为普通烟草K326,由国家农作物种质资源平台烟草种质资源子平台提供,2,4-表油菜素内酯购于索莱宝生物有限公司。
1.2 试验设计
在中国农业科学院烟草研究所即墨试验基地进行。共设置0.5×10-7 mol/L(T1)、0.5×10-5 mol/L(T2)两个浓度,以蒸馏水(CK)为对照。育苗基质(泥炭和蛭石比例为1∶1)经120 ℃灭菌20 min后装盆,种子直接播种于规格为5 cm×5 cm×7 cm的黑色塑料花盆中并于种子萌发一周后间苗,每盆保留一棵幼苗,每个处理30盆。当烟苗生长至4叶1心时,选出长势均匀一致的烟苗,每天于15:00喷施1次,叶片正面均匀喷施,喷至叶片上形成均勻的液滴为止,连续喷施5 d。
1.3 测定方法
1.3.1 烟草主茎生物学性状调查 喷施EBR处理5 d后,每个处理选长势均匀一致的植株10株,去除叶片和根系后留主茎拍照,用Image J软件测量株高(根茎结合处到心叶叶柄着生处的距离)和节距(自下而上第5节节距)。
1.3.2 茎细胞解剖学特征观察 取第5片真叶叶柄着生处1 cm长的茎组织,用FAA固定液(含5%醋酸,70%乙醇和5%甲醛,体积分数)固定,然后经乙醇逐级脱水,石蜡包埋,切割成5 μm薄片,甲苯胺蓝染色,明胶封片后于光学显微镜(Leica DMC 2900,上海徕卡仪器有限公司)下观察皮层薄壁细胞形态和细胞数目,用Image J软件计算细胞面积。
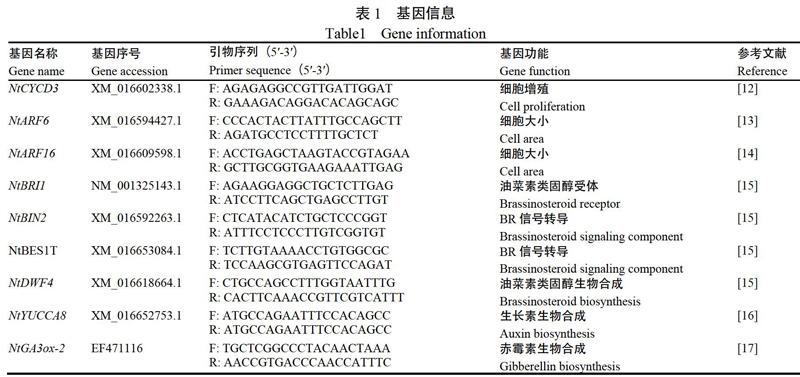
1.3.3 RNA提取和qRT-PCR 参考已发表的拟南芥的基因序列,在NCBI网站(https://www.ncbi.nlm. nih.gov/)中搜索获得BR、IAA、GA合成与转导相关的基因以及与细胞大小、细胞分裂相关基因的序列(表1)。
2 结 果
2.1 喷施EBR对烟草幼苗生物学性状的影响
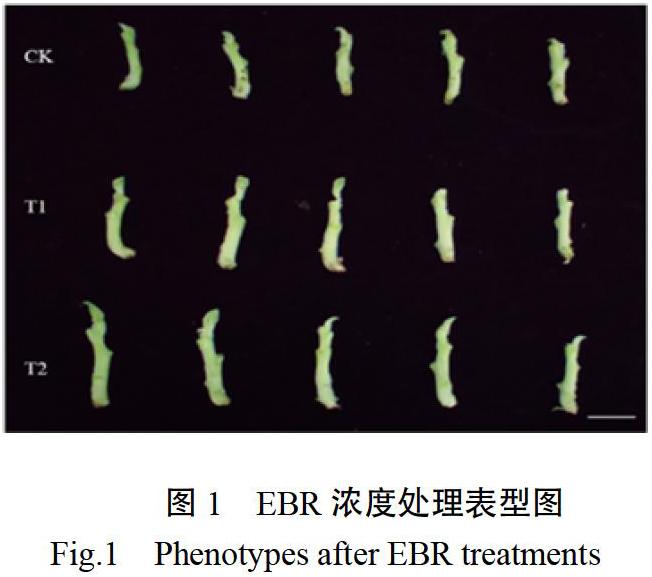
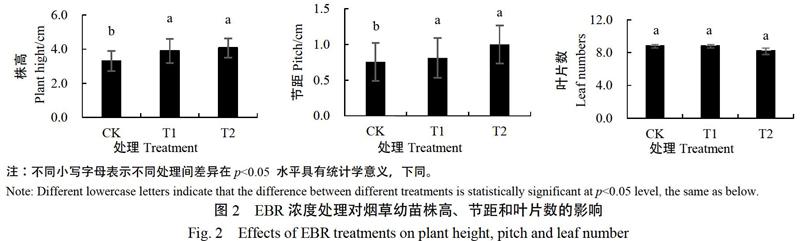
不同浓度EBR处理下烟苗主茎见图1,EBR叶面喷施5 d后观察茎的生物学性状发现(图2),EBR浓度越高,烟草幼苗株高越高,T1、T2较CK分别增加17.8%和23.13%。同时测量了自下而上第五节节距,发现节距也随着EBR浓度升高而增大,T1、T2较CK分别增加7.35%和32.35%。不同浓度EBR处理下叶片数没有显著差异。结果表明在本试验浓度范围内,EBR具有促进主茎伸长的作用。
2.2 喷施EBR对烟草皮层薄壁细胞的影响
EBR处理5 d后观察第5片真叶叶柄着生处茎的纵切面(图3),用Image J软件测量皮层薄壁细胞面积发现,EBR处理后皮层薄壁细胞面积较CK显著减小,而T1和T2处理间细胞面积无显著性差异。其中T1处理下皮层薄壁细胞面积为36.46 μm2,比CK(51.19 μm2)减小28.39%,T2处理下皮层薄壁细胞面积为35.16 μm2,比CK减小31.53%。虽然细胞面积减小,但是T1、T2处理下单位面积内细胞数目显著增加,其中T1处理下外皮层单位面积内细胞数目为1 325.67个/mm2,比CK(734.37 个/mm2)增加80.52%;T2处理下外皮层单位面积内细胞数目为1 267.27个/mm2,比CK增加73.02%。说明EBR处理使细胞数目增多,细胞面积减小。
2.3 喷施EBR对基因表达量的影响
不同浓度EBR处理24 h后,BR、IAA、GA相关基因和细胞分裂、细胞大小调控基因表达量如图4-6所示。由图4可见,T2处理细胞周期蛋白基因NtCYCD3表达量较CK增加105.2%。T1处理细胞大小调控基因NtARF6表达量较CK下降75.9%,T2处理较CK下降60.9%。T1处理细胞大小调控基因NtARF16表达量较CK下降39.4%,T2处理较CK下降23.9%。可见EBR处理促进了细胞分裂基因的表达,抑制了细胞大小调控基因的表达。
由图5可见,T2处理BR受体基因NtBR11表达量较CK增加234.1%,NtBIN2表达量较CK增加71.22%。T1处理BR信号转录因子NtBES1T的表达量较CK下降71.5%,T2处理较CK下降95.9%。由图6可见,T2处理BR合成基因NtDWF4表达量较CK提高105.5%,IAA生物合成基因NtYUCCA8表达量较CK增加36.3%。T1处理GA生物合成基因NtGA3ox-2表达量较CK增加116.3%,T2处理增加162.4%。EBR处理后BR、IAA、GA合成基因表达量均增加,对应受体基因表达量增加,转录因子表达量下降。说明EBR处理促进细胞分裂和BR、IAA、GA的合成。
3 讨 论
BR通过激活细胞分裂、促进细胞伸长而促进节间伸长[4-5]。在本试验中,不同浓度EBR处理后烟草幼苗主茎均显著伸长,叶片数无显著差异,节距显著增大,细胞数目增多(图1),因此EBR可能是通过促进节距增加而促进主茎伸长。细胞周期蛋白基因CYCD3具有通过调节细胞增殖影响植株叶形和细胞数目的作用[18],研究证明EBR促进AtCYCD3的转录促进细胞分裂[19]。本研究检测了茎中NtCYCD3的表达量,结果显示在T2处理下其表达量显著增加,说明NtCYCD3参与了EBR调控主茎伸长的过程。BR可使细胞壁松弛,促使细胞摄入水分而产生机械膨大[8]。本试验中细胞并未膨大而是出现了面积减小的现象,这一现象出现的原因有待进一步研究。NtARF6和NtARF16能调控细胞分裂和细胞大小,NtARF16基因过表达拟南芥叶片的细胞面积显著增大[14]。本研究发现EBR处理后NtARF6和NtARF16在茎中的表达量下降,细胞面积减小。以上结果从转录水平解释了BR通过促进细胞分裂,增加细胞数量进而促进茎伸长生长。
在BR信号转导途径中,细胞膜外受体激酶BRI1与BR结合后,使BKI1磷酸化,同时BIN2被磷酸酶BRI1-SUPPRESSO1(BSU1)脱磷酸化并被降解,抑制了BES1/BZR1的表达[20-21]。之后,BR通过调节编码XET基因的表达来促进细胞壁的松弛,减小壁压,降低水势,使水分和养分进入细胞,促使细胞扩大[8]。在本试验中,T2处理中BR促进了NtBRI1和NtBIN2在茎上部的表达,在T1和T2处理中BR抑制了NtBES1T的表达。而T1浓度处理下BR的促进效应并不显著,可能是因为烟草主茎对低浓度BR并不敏感。这些结论说明EBR处理可以提高BR信号转导途径中基因的表达,进而促进BR信号转导激活BR反应。
BRs合成酶DWF4是BR合成关键酶[22],DWF4的转录表达量与有活性的BR含量呈正相关[23]。拟南芥中,YUCCA能催化吲哚-3-丙酮酸(IPyA)氧化脱羧过程中的不可逆限速步骤,形成IAA[24]。研究表明,BR调节GA生物合成以调节细胞伸长[25]。通过分析BR突变体,发现GA含量以及GA代谢基因(包括GA20ox-2,GA3ox-2和GA2ox-3)的表达与BR含量相关[26]。本研究通过对BR、IAA和GA的合成基因进行检测,发现EBR处理除了诱导BR合成关键基因NtDWF4表达上调外,还促进了植物IAA生物合成基因NtYUCCA8和GA生物合成基因NtGA3ox-2的表达(图5),而且IAA的合成和应答反应对高浓度的EBR响应敏感。相较于IAA,GA对BR的反应似乎更为敏感,NtGA3ox-2的表达量在T1浓度处理下就已显著升高,在T2浓度处理下表达量变化较T1更加显著。这也充分说明BR可影响内源激素的合成。
4 结 论
试验结果表明,喷施0.5×10-7 和0.5×10-5 mol/L EBR使烟草幼苗节距和株高增加,并且随浓度升高增加效应增大。EBR通过促进细胞分裂进而促进茎的伸长。EBR处理后茎皮层薄壁细胞数目增多,细胞分裂基因表达量增加;细胞面积减小,细胞大小调控基因表达量下降。同时,喷施EBR会促进内源BR信号转导相关基因和BR、IAA、GA合成基因的表达。
参考文献
[1]贺学礼. 植物学[M]. 北京:高等教育出版社,2010.
HE X L. Botany[M]. Beijing: Higher Education Press, 2010.
[2]邹江石,姚克敏,吕川根,等. 水稻两优培九株型特征研究[J]. 作物学报,2003,29(5):652-657.
ZHOU J S, YAO K M, LV C G, et al. Study on individual plant type character of liangyoupeijiu rice[J]. Acta Agronomica Sinica, 2003, 29(5): 652-657.
[3]王晓萌. 水分胁迫对烟苗主茎生长发育与结构功能的影响[D]. 北京:中国农业科学院,2019.
WANG X M. Effects of water stress on growth, development and structure function of main stem of tobacco seedlings[D]. Beijing: Chinese Academy of Agricultural Sciences, 2019.
[4]何少勇. 玉米矮稈突变体的等位性鉴定及对外源激素的敏感性研究[D]. 雅安:四川农业大学,2017.
HE S Y. Allelic identification of maize dwarf mutants and sensitivity to exogenous hormones[D]. Yaan: Sichuan Agricultural University, 2017.
[5]TONG H N, CHU C C. Functional specificities of brassinosteroid and potential utilization for crop improvement[J]. Trends in Plant Science, 2018, 23: 13-27.
[6]叶梅荣,朱昌华,甘立军,等. 激素间相互作用对植物茎伸长生长的调控综述[J]. 中国农学通报,2007,23(4):238-241.
YE M R, ZHU C H, GAN L J, et al. Review on the regulation of plant stem elongation and growth by hormone interactions[J]. Chinese Agricultural Science Bulletin, 2007, 23(4): 238-241.
[7]丁丹阳,张璐翔,朱智威,等. 叶面喷施2,4-表油菜素内酯对烟草抗旱性的影响[J]. 中国烟草科学,2018,39(4):50-57.
DING D Y, ZHANG L X, ZHU Z W, et al. Effect of leaf spray 2,4- epibrassinolide on drought resistance of tobacco[J]. China Tobacco Science, 2018, 39(4): 50-57.
[8]鲁旭东,刘华英. 油菜素内酯促进植物伸长生长机理研究进展[J]. 生物学教学,2005,30(6):3-5.
LU X D, LIU H Y. Research progress on the mechanism of brassinolide promoting plant elongation and growth[J]. Biology Teaching, 2005, 30(6): 3-5.
[9]MAEDA N, KUNO N, YOKOTA T, et al. Effects of brassinolide on cell division and elongation in epicotyl growth of pea seedlings[J]. Plant and Cell Physiology, 2001, 42(1): 80-83.
[10]徐興阳,欧阳进,张俊文,等. 烤烟品种数量性状与烟叶产量和产值灰色关联度分析[J]. 中国烟草科学,2008,29(2):23-26.
XU X Y, OUYANG J, ZHANG J W, et al. Greycorrelation analysis of quantitative characters of flue-cured tobacco varieties and tobacco leaf yield and output value[J]. China Tobacco Science, 2008, 29(2): 23-26.
[11]殷英,张玉,余祥文,等. 烤烟主要农艺性状与产量产值的关系研究[J]. 中国烟草科学,2012,33(6):18-22.
YIN Y, ZHANG Y, YU X W, et al. Relationships among main agronomic attributes and yields and output value of flue-cured tobacco[J]. China Tobacco Science, 2012, 33(6): 18-22.
[12]乐丽娜,黄敏仁,陈英. 植物叶形态建成的分子机理研究进展[J]. 分子植物育种,2016,14(11):3205-3213.
LE L N, HUANG M R, CHEN Y. Research progress on the molecular mechanism of plant leaf morphogenesis[J]. Molecular Plant Breeding, 2016, 14(11): 3205-3213.
[13]LIU K, LI Y H, CHEN X N, et al. ERF72 interacts with ARF6 and BZR1 to regulate hypocotyl elongation in Arabidopsis[J]. Journal of Experimental Botany, 2018, 69(16): 13-27.
[14]周蕾. 烟草叶片发育相关ARF和GRF家族基因的表达分析与功能验证[D]. 北京:中国农业科学院,2020.
ZHOU L. Expression analysis and functional verification of ARF and GRF family genes related to tobacco leaf development[D]. Beijing: Chinese Academy of Agricultural Sciences, 2020.
[15]WANG S S, SUN H J, TANG M Y, et al. Reactive oxygen species-mediated BIN2 activity revealed by single-molecule analysis[J]. New Phytologist, 2019, 223(2): 692-704.
[16]ZHAO Y. The Roles of YUCCA Genes in local auxin biosynthesis and plant development[J]. Plant Developmental Biology, 2010(2): 227-235.
[17]CURABA J. AtGA3ox-2, a key gene responsible for bioactive gibberellins biosynthesis is regulated during embryogenesis by LEAFY COTYLEDON 2 and FUSCA 3 in Arabidopsis[J]. Plant physiology, 2004, 136(3): 3660-3669.
[18]DECHKRONG P, YOSHIKAWA T, ITOH J I. Morphological and molecular dissection of leaf development in wild-type and various morphogenetic mutants in rice[J]. American Journal of Plant Sciences, 2015, 6(8): 1215-1232.
[19]HU Y, BAO F, LI J. Promotive effect of brassinosteroids on cell division involves a distinct CycD3-induction pathway in Arabidopsis[J]. The Plant Journal, 2000, 24(5): 693-701.
[20]GUO H Q, LI L, ALURU M, et al. Mechanisms and networks for brassinosteroid regulated gene expression[J]. Current Opinion in Plant Biology, 2013, 16(5): 545-553.
[21]PENG P, YAN Z Y, ZHU Y Y, et al. Regulation of the Arabidopsis GSK3-like kinase brassinosteroid-insensitive 2 through proteasome-mediated protein degradation[J]. Molecular Plant, 2008, 1(2): 338-346.
[22]CHOE S, DILKES BP, FUJIOKA S, et al. The DWF4 gene of Arabidopsis encodes a cytochrome P450 that mediates multiple 22α-Hydroxylation steps in brassinosteroid biosynthesis[J]. The Plant Cell, 1998, 10(2): 231-243.
[23]張东芝. 拟南芥油菜素内酯合成关键酶DWF4的功能研究[D]. 兰州:兰州大学,2018.
ZHANG D Z. Study on the function of the key enzyme DWF4 in brassinolide synthesis in arabidopsis [D]. Lanzhou: Lanzhou University, 2018.
[24]CAO C, YANG H L, SHANG C Q, et al. The roles of auxin biosynthesis YUCCA gene family in plants[J]. Molecular Science, 2019, 20(24): 6343-6352.
[25]TONG H, XIAO Y, LIU D, et al. Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in rice[J]. Plant Cell, 2014, 26(11): 4376-4393.
[26]MAO J, ZHANG D, LI K, et al. Effect of exogenous Brassinolide (BR) application on the morphology, hormone status, and gene expression of developing lateral roots in Malus hupehensis[J]. Plant Growth Regulation, 2017, 82(3): 1-11.
