红掌花青素合成相关基因的研究进展
2021-08-06俞政男冯欣洁张志胜曾瑞珍谢利郭和蓉
俞政男 冯欣洁 张志胜 曾瑞珍 谢利 郭和蓉
摘要 红掌是仅次于兰花的第二大热带观赏植物。佛焰苞颜色是红掌育种的主要目标性状,与其所含的花色素种类和含量密切相关。研究表明,花青素是影响红掌花色的主要色素之一,其生物合成受结构基因和调节基因的共同调控。综述了红掌佛焰苞中花青素生物合成相关结构以及调节基因克隆和功能研究进展,为红掌花色的分子育种提供指导。
关键词 红掌;花青素;结构基因;调节基因;分子育种
中图分类号 S682.14 文献标识码 A 文章编号 0517-6611(2021)13-0005-05
doi:10.3969/j.issn.0517-6611.2021.13.002
开放科学(资源服务)标识码(OSID):
Research Progress of Anthocyanin Biosynthesis Related Genes in Anthurium andraeanum
YU Zheng nan,FENG Xin jie, ZHANG Zhi sheng et al
(College of Forestry and Landscape Architecture,South China Agricultural University,Guangzhou,Guangdong 510642)
Abstract Anthurium andraeanum is the second largest tropical ornamental plant after orchid. The color of spathe is the main target character in Anthurium andraeanum breeding, which is closely related to the anthocyanin type and content. The results show that anthocyanins are one of the main pigments affecting Anthurium andraeanum spathe color, and its biosynthesis is regulated by both structural and regulatory genes. In this paper, the research progress of anthocyanin biosynthesis related structure and regulatory gene cloning and function in anthocyanin biosynthesis in Anthurium andraeanum was reviewed, which could provide guidance for molecular breeding of anthocyanin biosynthesis in Anthurium andraeanum.
Key words Anthurium andraeanum;Anthocyanin;Structural gene;Regulator gene;Molecular breeding
红掌(Anthurium andraeanum)是天南星科(Araceae)花烛属(Anthurium)植物,原产于哥伦比亚,是世界上销量仅次于兰花的第二大贸易花卉[1]。因其具有较高的观赏价值,深受消费者喜爱。佛焰苞是红掌的主要观赏部位,其造型独特,色彩艳丽,以红色、粉色、橙色、珊瑚色和白色为主,还有紫色、绿色、褐色和复色等[2]。研究表明,影响红掌佛焰苞呈色的色素主要是花青素、黄酮、黄酮醇和叶绿素,其中,花青素是佛焰苞中最主要的色素[3]。
花青素又称花青素苷(anthocyanin),是植物类黄酮代谢的中间产物,属有色类黄酮一族,主要累积在叶片、花瓣、果实表皮细胞的液泡内,是构成植物花和果实的主要色素之一[4]。花青素的合成受两类基因的共同调控,一类是编码花青素生物合成途径中各种酶的结构基因;另一类是通过编码转录因子来调控结构基因时空表达的调节基因,调控花青素的种类和数量。目前,已经鉴定和克隆出部分与花青素合成相关的结构基因和转录因子。该研究拟综述红掌花青素合成相关基因的研究进展,揭示红掌佛焰苞颜色形成的机制,为红掌花色改良提供参考。
1 花青素与红掌花色形成
花青素具有类黄酮C6-C3-C6的C骨架的典型结构,由于花色基团在结构中R1和R2位置上的取代基不同,形成了花青素的颜色差异[5]。目前已知的花青素种类超过550种,最常见的有6种:矢车菊色素(cyanidin)、天竺葵色素(plelargonidin)、飞燕草色素(delphinidin)、芍药花色素(peonidin)、牵牛花色素(petunnidin)和锦葵色素(malvidin)[6-7]。矢车菊素通常使花瓣呈现红色或粉红色,天竺葵色素呈现橙红色,飞燕草色素呈现蓝色,这3种色素通常单独存在于不同的花中。牵牛花色素、芍药花色素和锦葵色素均是以上3种色素不同程度甲基化作用的产物[8-9]。由于花青素在植物体内的存在不稳定,常与葡萄糖、半乳糖、芸香糖等单糖结合形成花青素苷[4]。花青素苷的化学性质不稳定,会随pH的变化而变化,酸性条件下花色呈红色、紫色,碱性条件下花色呈蓝色[10]。
红掌佛焰苞最主要的颜色是红色系(包含粉色)和橙色系(包含珊瑚色),由天竺葵色素和矢车菊色素控制[11-12]。当只有天竺葵色素存在时,佛焰苞呈橙色和珊瑚色;天竺葵色素和矢车菊色素同时存在时,佛焰苞则呈红色和粉色,且橙色和红色佛焰苞中的花青素浓度均高于珊瑚色和粉色佛焰苞[13]。除了花青素之外,红掌色素还包括叶绿素、类胡萝卜素、黄酮和黄酮醇等化合物。淡黄色和白色佛焰苞色素的主要成分是黄酮和黄酮醇类化合物,绿色佛焰苞中的主要色素是叶绿素,类胡萝卜素只存在于少部分棕色佛焰苞和肉穗花序中[14]。研究表明,佛焰苞中的花青苷含量越高,颜色越偏向红色;叶绿素含量越高,佛焰苞颜色越绿;2种色素同时存在时佛焰苞会呈现棕红色[3]。由于紅掌佛焰苞中缺少飞燕草色素或芍药花色素,因此目前还没有发现蓝色佛焰苞的红掌品种。
2 红掌花青素生物合成途径与相关基因研究
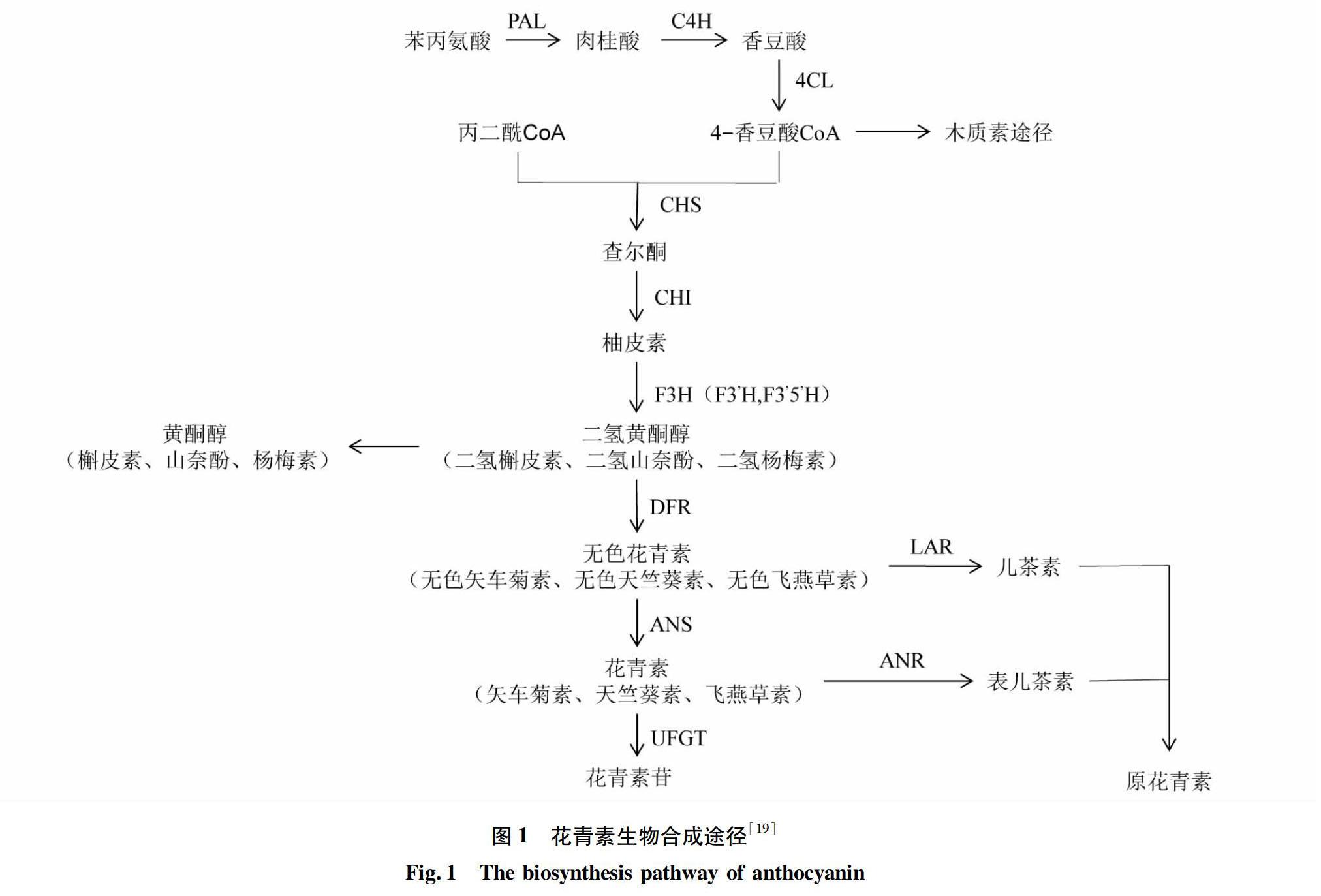
花青素的生物合成受到结构基因和调节基因的共同调控[15],作为植物类黄酮途径的一个分支,花青素生物合成途径研究得比较透彻。目前已经从矮牵牛(Petunia hybrida)、非洲菊(Gerbera jamesonii)、石竹(Dianthus chinensis L.)、金鱼草(Antirrhinum majus L.)等花卉中分离和克隆出一些结构基因和调节基因[16]。根据红掌佛焰苞中花青素合成相关基因的研究结果,参考拟南芥(Arabidopsis thaliana)、矮牵牛(Petunia hybrida)等模式植物的花青素生物合成途径,绘制出红掌的花青素生物合成途径(图1)。花青素生物合成分3个阶段,第1阶段是苯丙烷途径,苯丙氨酸在苯丙氨酸解氨酶(PAL)、肉桂酸4-羟化酶(C4H)、4-香豆酸辅酶A连接酶(4CL)等酶的作用下生成花青素的合成前体——香豆酰辅酶A;第2阶段是类黄酮代谢,查尔酮合成酶(CHS)催化合成柚皮素查爾酮,启动花青素的合成,经过类黄酮3-羟化酶(F3H)的催化生成二氢黄酮醇;第3阶段是花青素的生成,二氢黄酮醇在类黄酮3-羟化酶(F3H)、二氢黄酮醇还原酶(DFR)的作用下生成各种无色花青素,再经过花青素合成酶(ANS)的催化生成花青素[17],最后经过类黄酮3-O-糖基转移酶(UFGT)催化为稳定的花青素苷[18]。无色花青素和花青素也可能分别经过LAR和ANR催化生成原花青素。
2.1 结构基因
2.1.1 C4H(cinnamic acid 4 hydroxylase)。
C4H 是苯丙烷代谢途径上的重要调控酶,催化肉桂酸生成4-香豆酸。除了花青素合成途径外,C4H还是原花青素途径、木质素途径等不同次生代谢途径上的关键基因[20],在植物生长发育过程中发挥着重要作用[21]。研究表明,AnC4H在红掌各组织中均有表达但主要集中在佛焰苞中,与佛焰苞花青素含量呈正相关[22]。用乙烯利处理能够提高AnC4H的表达量,从而促进花青苷的累积[23],进一步说明AnC4H基因在佛焰苞花青素合成过程中起正向调控的作用[23]。
2.1.2 CHI(chalcone isomerase)。
CHI 是类黄酮物质代谢途径上的第2个关键酶[24]。研究发现,CHI基因具有组织表达特异性,在花青素含量高的植物组织中表达量较高,其表达水平也会随着发育阶段的变化有所不同[25]。利用RNAi技术抑制烟草CHI基因的表达,查尔酮含量上升,花瓣呈现黄色[26];矮牵牛中CHI基因发生突变后花粉会因查尔酮的积累而成黄色或绿色。相反,提高CHI的活性会使黄酮类化合物含量显著上升[27]。研究表明,AnCHI的表达具有组织差异性和时空特异性,其在佛焰苞中的表达量明显高于根、茎和肉穗花序,并且在佛焰苞早期的表达量高于成熟期,AnCHI基因表达量与花青素的含量呈正相关,表明AnCHI基因可能是控制佛焰苞花青素合成的关键基因,在花青素的合成过程中起正向调控作用[28-29]。但也有研究认为由于CHI属于类黄酮生物合成的上游基因,不仅参与花青素的合成还参与了所有类黄酮化合物的合成,因此CHI的表达量变化与红掌佛焰苞花色的相关性不高[30]。
2.1.3 F3H(flavonone 3 hydroxylase)。
F3H位于花青素合成途径上的中枢位点,既可以催化无色的二羟基黄烷酮生成二氢黄酮醇,也可以生成黄烷酮醇。F3H可以单独调控花青素的合成,但大多情况下都是与CHS等酶共同催化黄酮类化合物的生成。通过改变植物体内的F3H基因的表达量可以对植物体内的花青素进行调节,进而改变花色,在花青素的合成和累计中具有重要作用[31]。Collette等[32]最早获得了红掌F3H的全长序列,并发现F3H与CHS、ANS共同调控红掌花青素的合成。佛焰苞中F3H的表达量下降将导致二氢黄酮醇等下游产物含量降低,表明F3H是改变红掌佛焰苞颜色的关键基因[33]。在部分红掌品种中,F3H表达量的增加会使佛焰苞的明度增加,红色减弱[14]。此外,当植物面临逆境胁迫时,F3H能够促进体内花青素的积累,提高抗氧化能力,从而抵御逆境。
2.1.4 F3H和F35H(flavonone 3-hydroxylase and flavonone 35-hydroxylase)。
F3H催化二氢黄酮醇生成的二氢槲皮黄酮是合成红色矢车菊色素的重要中间产物,对粉色和红色花的形成具有重要作用[34]。在一个突变为橙色的佛焰苞中发现,F3H在突变部位的表达量下调,矢车菊色素的含量也大幅下降,突变佛焰苞中累积的色素主要是天竺葵色素,证明F3H是控制红掌佛焰苞呈现红色的关键基因[35]。通过控制F3H基因的表达,可以改变红掌佛焰苞中矢车菊色素的累积,对于改良佛焰苞颜色具有重要意义。
F35H是合成飞燕草色素、牵牛花色素、锦葵色素的关键酶,催化生成的二氢杨梅黄酮是合成飞燕草类花青素的前体物质,缺乏F35H就无法形成蓝色,因此F35H也被称为蓝色基因[36]。F35H自然界中天然蓝紫色花卉较少,石竹、玫瑰、郁金香等花卉都没有蓝色品种,因此克隆F35H基因为培养出蓝色的花卉提供了可能[34]。如果将F35H转入植株内,可以提高飞燕草色素的累积,改变花色素的组成,从而改良花色[37]。目前,在红掌中未发现F35H基因表达的研究。
2.1.5 DFR(dihydroflavonol 4 reductase)和ANS(anthocyanidin synthase)。
DFR和ANS是花青素合成末期的关键酶,共同将二氢黄酮醇催化为有色花青素,对花色最终形成起了决定性的作用[38-39]。DFR在进化过程中高度保守,并且具有组织表达特异性和时空表达特异性[40]。由于DFR对底物有特异性,表达水平不同,植物颜色各异。失去DFR活性的植物突变体会变成象牙色或白色,将DFR异源转入白色的康乃馨(Dianthus ‘Carnation)中,花瓣变为紫色;橙花龙胆(Gentiana scabra Bunge)中的DlaDFR1和DlaDFR2过表达会使矢车菊色素含量降低,颜色偏红[19,41]。作为花青素合成途径中的关键节点基因,DFR常被用于植物的红花分子育种中[38]。研究发现AnDFR在早晨和傍晚的佛焰苞中表达量偏高,中午最低,因此其表达具有昼夜节律[42],AnDFR在佛焰苞中的表达量与花青素含量的变化趋势一致,表明AnDFR基因的表达量与佛焰苞花青素的含量呈正相关[43]。ANS在花器官中的表达量高于根、茎、叶等组织,在颜色较深或者鲜艳的部位中表达量也显著高于其他部位。ANS的过表达会增加花青素的累积,表达量降低会导致花青素含量的减少。NtANS1和NtANS2在烟草花成熟期的表达量增加,发育后期降低;蓝猪耳(Torenia concolor Lindl.)中ANS的缺失会导致花由蓝色变为白色[44-45]。研究发现,ANS与DFR在佛焰苞中的表达量越高,佛焰苞的颜色越红[30],是红色花色遗传模型相关的关键基因[46]。
2.1.6 ANR和LAR(anthocyanidin reductase and leucoanthocyanidin reductase)。
ANR(花青素还原酶)是合成原花青素的关键酶,该酶可直接将花青素催化生成原花青素,而不经过UFGT(类黄酮糖基转移酶)的催化生成稳定的花青素苷,因此ANR基因与花青素的合成竞争同一底物,在花青素的合成途径中有负调控的作用[47]。LAR(无色花青素还原酶)是植物类黄酮化合物合成途径中的一个关键酶,催化无色花青素转化为儿茶素[48],LAR由ANR经过MYB转录因子家族基因共同调控。目前已经在拟南芥、茶树、芒果等植物中克隆得到了LAR基因[49]。
最早在拟南芥中发现ANR对种皮的颜色起负调控作用[50],缺少ANR基因会促进花青素的合成[51]。研究发现ANR在红色系的佛焰苞中表达量较低,在白色系的佛焰苞中表达量较高,表明ANR基因的表达量与花青素的含量呈负相关,推测ANR可能是调控红掌佛焰苞中原花青素合成的关键基因[52]。LAR与DFR共同影响无色花青素的合成[53],ANR和LAR基因表达会促进原花青素的合成,减少花青素的累积,对花青素的累积起负调控的作用,可能是调节红掌佛焰苞花青素和原花青素平衡的关键酶[30]。但ANR和LAR如何作用于佛焰苞花青素的合成有待进一步研究。
2.2 调节基因
调节基因编码转录因子,转录因子与结构基因启动子中相应的顺式作用元件结合,从而调控结构基因的时空表达[16]。研究发现,对花青素合成途径中的结构基因起调控作用的转录因子主要有3类:MYB蛋白、bHLH蛋白和WD40蛋白[54]。研究表明,红掌花青素合成主要受R2R3-MYB家族基因和 bHLH家族基因调控。Elibox等[55]认为红掌佛焰苞颜色受到M、O、R 3个显性基因控制,R基因编码一个转录因子,调控CHS、F3H、ANS的表达,O基因编码一个单独的转录因子调控DFR的表达。显性的O和R产生有色佛焰苞,O和R只要有一个为隐性则产生白色佛焰苞,M基因调控F3H基因决定佛焰苞属于红色系还是橙色系。O和R的调控机制尚未被证实,很有可能代表MYB家族和bHLH家族的转录因子[56]。
2.2.1 R2R3-MYB转录因子。
MYB蛋白属于DNA结合蛋白,具有一段高度保守的MYB结合域,其结构域由52个氨基酸构成,其中有高度保守的氨基酸和间隔序列,这些保守的氨基酸使MYB蛋白结构域折叠成HHTH(helix-helix-turn-helix)结构[57]。根据R基序数目的不同,可以将MYB蛋白分为R2R3 MYB、1R MYB、3R MYB、4R MYB。其中,R2R3 MYB是花青素合成途径中最广泛的调控因子,能够激活一个或多个结构基因的表达[58],在决定花青素的着色方式中起关键作用[59-60]。现已从百合(Lilium brownii var.)、月季(Rosa chinensis Jacq.)、玫瑰(Rosa rugosa Thunb.)等植物中分离并鉴定出参与花青素合成调控的R2R3 MYB转录因子[58,61]。
大部分R2R3 MYB转录因子对花青素的累积有促进作用。Collette[42]克隆得到了红掌第一个R2R3 MYB转录因子——AaMYB1,将AaMYB1和调控玉米花青素合成的bHLH基因ZmLc转入白色花的大花蕙兰(Cymbidium hybrid),花瓣出现含有花青素的斑点[62]。AaMYB1转入烟草后得到花青素显著累积的转化体[63]。AaMYB1的单独过表达导致烟草积累大量花青素,同时使花青素合成的关键基因NtDFR、NtANS和R2R3 MYB基因NtAn2表达量大幅上升,表明AaMYB1具有正调控花青素合成的功能[64]。Li等[65]研究发现AaMYB2转录因子与AaF3H、AaANS、AaCHS等基因的表达规律一致,推测AaMYB2在红掌佛焰苞花青素累积中起正调控的作用。也有一些R2R3 MYB转录因子对花青素的累积起抑制作用。如AaMYB3单独表达或与AabHLH1共同作用时,转基因烟草花瓣中的花青素含量会减少,而原花青素含量上升,导致花色变浅,证明AaMYB3对花青素的合成起负调控作用[66]。
2.2.2 bHLH(basic helix loop helix)。
bHLH是根据其蛋白保守结构域命名的一类转录因子家族,主要调节类黄酮和花青素合成以及参与植物的生长发育、生理代谢和逆境应答等[67]。在花青素合成途径的转录调控过程中,bHLH通常与MYB蛋白共同起作用,MYB负责结合和活化转录的功能,bHLH负责增强MYB蛋白结构的稳定性和转录活性。bHLH和MYB之间的相互作用是花青素合成途径调控研究的新热点[8,66]。Jiang等[68]研究发现红掌叶色突变体会累积大量花青素并且叶色更绿、抗逆性更强,将分离得到的AaMYB3和AabHLH35基因转入拟南芥中,能够提高其抗寒性和抗旱性,證明AabHLH35在花青素合成累积和提高植物抗性方面具有重要作用。并不是所有的bHLH调节基因都对花青素的累积起促进作用,也有一些起抑制作用。比如玉米中缺失bHLH基因的突变体中,UFGT基因的表达量上升,花青素苷的含量也随之上升;Ph MYB27与矮牵牛bHLH蛋白JAF13、AN1结合时,会导致花青素的合成受到抑制[69]。
3 问题与展望
红掌花青素的研究取得了一定的进展,已经从红掌中克隆和分离得到了一些与花青素合成相关的结构基因和调节基因,有利于科研工作者通过遗传改良等方法使红掌花青素的合成和累积朝着市场需求的方向发展。目前,已经克隆出来的与红掌佛焰苞颜色相关的基因有PAL、C4H、CHI、CHS、F3H、DFR、ANR、LAR、AaMYB1、AaMYB2、AaMYB3、AabHLH35等,但是与模式作物相比,红掌的花青素合成途径以及调控网络还不够明确,花青素的合成代谢过程也非常复杂,还需要进一步完善。此外,在一些模式植物中发现花青素合成受到3类转录因子(MYB转录因子家族基因、bHLH转录因子家族基因和WD40蛋白家族)的调控,这3类转录因子相互作用,共同调控花青素的合成。但是在红掌中尚未报道有关WD40蛋白家族的研究,这类蛋白对红掌花青素的合成是否有影响还需要进一步研究。
佛焰苞的顏色是红掌育种工作者关注的重要性状。目前主要通过传统育种方式获得不同佛焰苞的颜色,但杂交育种花色选择的目标性不强。开展红掌花青素合成相关基因功能研究,完善花青素合成途径的代谢机制,运用到红掌新品种分子育种研究中,为定向改良和修饰红掌花色提供新的思路,对红掌花色育种工作具有一定的指导意义。
参考文献
[1] 欧文军,李洪立,尹俊梅.红掌切花栽培中常见病虫害及防治[J].云南农业科技,2002(4):34-37.
[2] 牛俊海,冷青云,林兴娥,等.红掌基因工程育种进展、问题与对策[J].基因组学与应用生物学,2014,33(5):1147-1153.
[3] 杨澜,李崇晖,黄素荣,等.红掌佛焰苞中花色素与颜色形成的关系[J].北方园艺,2012(15):68-73.
[4] 曹琳娇,李晓杰,焦棒棒,等.蔬菜花青苷生物合成及转录调控的研究进展[J].中国瓜菜,2019,32(12):1-7.
[5] 邵文婷,刘杨,韩洪强,等.茄子花青素合成相关基因SmMYB的克隆与表达分析[J].园艺学报,2013,40(3):467-478.
[6] 贾赵东,马佩勇,边小峰,等.植物花青素合成代谢途径及其分子调控[J].西北植物学报,2014,34(7):1496-1506.
[7] YOU Q,WANG B W,CHEN F,et al.Comparison of anthocyanins and phenolics in organically and conventionally grown blueberries in selected cultivars[J].Food chemistry,2011,125(1):201-208.
[8] GROTEWOLD E.The genetics and biochemistry of floral pigments[J].Annual review of plant biology,2006,57(1):761-780.
[9] KOWALCZYK E,KRZESIN'SKI P,KURA M,et al.Anthocyanins in medicine[J].Polish journal of pharmacology,2003,55(5):699-702.
[10] 单建伟,刘晓津,索海翠,等.植物花色苷的研究进展[J].南方农业学报,2019,50(2):278-285.
[11] IWATA R Y,TANG C S,KAMEMOTO H.Anthocyanins of Anthurium andraeanum Lin.[J].J Amer Soc Hort Sci,1979,51(104):1-6.
[12] MARUTANI M,TANG C S,PAULL R,et al.Anthocyanins in the lavender Anthurium[J].HortScience,1987,22:620-622.
[13] 丛汉卿,徐立,信彩云,等.红掌突变体花青素含量的测定与比较[J].安徽农业科学,2009,37(17):7832-7834.
[14] 杨澜.红掌佛焰苞中花色素组成和含量及关键基因的表达与颜色形成的关系[D].海口:海南大学,2014.
[15] GROTEWOLD E.The genetics and biochemistry of floral pigments[J].Annual review of plant biology,2006,57(1):761-780.
[16] 李莹,高振蕊,张驰,等.花青素合成途径中分子调控机制的研究进展[J].生态学杂志,2015,34(10):2937-2942.
[17] 侯泽豪,王书平,魏淑东,等.植物花青素生物合成与调控的研究进展[J].广西植物,2017,37(12):1603-1613.
[18] 孙传范.原花青素的研究进展[J].食品与机械,2010,26(4):146-148,152.
[19] HOLTON T A,CORNISH E C.Genetics and biochemistry of anthocyanin biosynthesis[J].The plant cell,1995,7(7):1071-1083.
[20] GRAVOT A,LARBAT R,HEHN A,et al.Cinnamic acid 4 hydroxylase mechanism based inactivation by psoralen derivatives:Cloning and characterization of a C4H from a psoralen producing plant Ruta graveolens exhibiting low sensitivity to psoralen inactivation[J].Archives of biochemistry & biophysics,2004,422(1):71-80.
[21] CHEN A H,CHAI Y R,LI J N,et al.Molecular cloning of two genes encoding cinnamate 4 hydroxylase(C4H)from oilseed rape(Brassica napus)[J].Journal of biochemistry & molecular biology,2007,40(2):247-260.
[22] 楊哲,刘克林,彭佳佳,等.红掌查尔酮异构酶基因的克隆与表达分析[J].园艺学报,2016,43(7):1402-1410.
[23] 郑高言,张黎,刘克林,等.乙烯利对红掌‘特伦萨花青苷合成及相关基因的影响[J].分子植物育种,2018,16(24):8041-8047.
[24] MUIR S R,COLLINS G J,ROBINSON S,et al.Overexpression of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols[J].Nature biotechnology,2001,19(5):470-474.
[25] ZHANG Z R,QIANG W,LIU X Q,et al.Molecular cloning and characterization of the chalcone isomerase gene from sweetpotato[J].African journal of biotechnology,2011,10(65):14443-14449.
[26] NISHIHARA M,NAKATSUKA T,YAMAMURA S.Flavonoid components and flower color change in transgenic tobacco plants by suppression of chalcone isomerase gene[J].FEBS letters,2005,579(27):6074-6078.
[27] LI F,JIN Z,QU W,et al.Cloning of a cDNA encoding the Saussurea medusa chalcone isomerase and its expression in transgenic tobacco[J].Plant physiol biochem,2006,44(7/8/9):455-461.
[28] 杨哲.红掌花青素合成相关基因克隆与表达分析的研究[D].北京:中国农业科学院,2016.
[29] 郑高言.红掌佛焰苞颜色相关基因AnCHI和AnC4H表达分析及功能验证[D].北京:中国农业科学院,2018.
[30] 沈成晨.红掌突变体花色素代谢机理的初步研究[D].苏州:苏州大学,2019.
[31] 范晶,黄明远,吴苗苗,等.山茶属三个F3H基因的分子特性、系统进化及蛋白结构差异分析[J].基因组学与应用生物学,2016,35(5):1195-1205.
[32] COLLETTE V E,JAMESON P E,SCHWINN K E,et al.Temporal and spatial expression of flavonoid biosynthetic genes in flowers of Anthurium andraeanum[J].Physiologia plantarum,2004,122(3):297-304.
[33] 彭佳佳,杨哲,李佩愉,等.花青素生物合成基因的表达对红掌佛焰苞颜色变化的影响[C]//中国园艺学会2014年学术年会论文摘要集.北京:《园艺学会》编辑部,2014.
[34] 赵云鹏,陈发棣,郭维明.观赏植物花色基因工程研究进展[J].植物学通报,2003,38(1):51-58.
[35] 李崇晖,牛俊海,黄素荣,等.红掌‘Arizona佛焰苞颜色部分突变的原因初探[J].分子植物育种,2015,13(3):627-633.
[36] 贾效成,李美茹,陈贻竹.蓝色玫瑰研究进展[J].生物学通报,2003,38(9):12-14.
[37] NAONOBU N,RYUTARO A,SANAE K,et al.Genetic engineering of novel bluer colored chrysanthemums produced by accumulation of delphinidin based anthocyanins[J].Plant & cell physiology,2013,54(10):1684-1695.
[38] 李方殷.葡萄风信子花色形成关键基因DFR原核表达及功能验证[D].杨凌:西北农林科技大学,2018.
[39] 占丽英,林义章.紫色芸薹属蔬菜花青苷生物合成调控研究进展[J].东南园艺,2016,4(1):35-41.
[40] 祝婷,李成磊,吴琦,等.苦荞和甜荞二氢黄酮醇4-还原酶基因(dfr)的克隆及序列分析[J].食品科学,2010,31(13):219-223.
[41] 于婷婷.橙花龙胆二氢黄酮醇4-还原酶(DFR)基因的功能分析[D].长春:长春师范大学,2019.
[42] COLLETTE V E.A molecular analysis of flower colour development in an ornamental monocot(Anthurium anadraeanum):A thesis presented in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Plant Molecular Biology at Massey University,Palmerston North,New Zealand[D].New Zealand:Massey University,2002.
[43] 李雪.红掌花青素合成途径相关基因的克隆与分析[D].海口:海南大学,2013.
[44] 卜星星,雒晓鹏,白悦辰,等.金荞麦花青素合酶基因的克隆及其表达与花青素量的相关性研究[J].中草药,2014,45(7):985-989.
[45] 蒋明,陈孝赏,李金枝.紫菜薹花青素合成酶基因BcANS的克隆、表达与序列分析[J].浙江大学学报(农业与生命科学版),2011,37(4):393-398.
[46] COLLETTE V E,JAMESON P E,SCHWINN K E,et al.Temporal and spatial expression of flavonoid biosynthetic genes in flowers of Anthurium andraeanum[J].Physiologia plantarum,2004,122(3):297-304.
[47] 马敬,苏磊,袁美,等.花生C4H和ANR基因的克隆与表达研究[J].核农学报,2012,26(1):43-48.
[48] 李蒙,鲍锋.不同植物无色花青素还原酶及其基因的生物信息学分析[J].江西农业学报,2017,29(9):5-9.
[49] 陈春艳,马晖玲,董文科,等.CPB-LAR-GFP表达载体的构建及其在紫花苜蓿愈伤组织中的表达[J].草原与草坪,2017,37(1):25-30.
[50] XIE D Y,SHARMA S B,DIXON R A.Anthocyanidin reductases from Medicago truncatula and Arabidopsis thaliana[J].Archives of biochemistry and biophysics,2004,422(1):91-102.
[51] ALBERT S,DELSENY M,DEVIC M.BANYULS,a novel negative regulator of flavonoid biosynthesis in the Arabidopsis seed coat[J].Plant journal,1997,11(2):289-299.
[52] 李雪,李志英,丛汉卿,等.红掌ANR基因克隆及其表达与佛焰苞颜色的相关性分析[J].分子植物育种,2013,11(6):825-830.
[53] 彭佳佳.红掌转录组测序及花青素生物合成途径差异表达基因的分析[D].银川:宁夏大学,2015.
[54] 宋雪薇,魏解冰,狄少康,等.花青素转录因子调控机制及代谢工程研究进展[J].植物学报,2019,54(1):133-156.
[55] ELIBOX W,UMAHARAN P.Inheritance of major spathe colors in Anthurium andraeanum Hort.is determined by three major genes[J].HortScience,2008,43(3):787-791.
[56] TEIXEIRA DA SILVA J A,DOBRNSZKI J,ZENG S J,et al.Genetic transformation and molecular research in Anthurium:Progress and prospects[J].Plant Cell Tiss Organ Cult,2015,123:205-219.
[57] 赵云鹏,陈发棣,郭维明.观赏植物花色基因工程研究进展[J].植物学通报,2003,38(1):51-58.
[58] 邹凯.玫瑰花青苷合成相关R2R3-MYB基因的克隆与表达分析[D].泰安:山东农业大学,2018.
[59] DUBOS C,STRACKE R,GROTEWOLD E,et al.MYB transcription factors in Arabidopsis[J].Trends in plant science,2010,15(10):573-581.
[60] HSU C C,CHEN Y Y,TSAI W C,et al.Three R2R3 MYB transcription factors regulate distinct floral pigmentation patterning in Phalaenopsis spp.[J].Plant physiology,2015,168(1):175-191.
[61] 丁芳兵,孫伟博,原雅玲,等.百合花色及相关育种研究进展[J].北方园艺,2017(18):170-175.
[62] ALBERT N W,ARATHOON S,COLLETTE V E,et al.Activation of anthocyanin synthesis in Cymbidium orchids:Variability between known regulators[J].Plant cell tissue & organ culture,2010,100(3):355-360.
[63] 马广莹,史小华,朱开元,等.红掌AaMYB1基因的克隆、表达及异源转化研究[J].核农学报,2017,31(9):1708-1716.
[64] 李崇晖,杨光穗.红掌AaMYB1的表达特征及其在烟草中的过表达[J].分子植物育种,2019,17(6):1898-1905.
[65] LI C H,QIU J,YANG G S,et al.Isolation and characterization of a R2R3 MYB transcription factor gene related to anthocyanin biosynthesis in the spathes of Anthurium andraeanum(Hort.)[J].Plant cell reports,2016,35(10):2151-2165.
[66] LI C H,QIU J,HUANG S R,et al.AaMYB3 interacts with AabHLH1 to regulate proanthocyanidin accumulation in Anthurium andraeanum(Hort.) another strategy to modulate pigmentation[J].Horticulture research,2019,6(14):1-16.
[67] 杨鹏程,周波,李玉花.植物花青素合成相关的bHLH转录因子[J].植物生理学报,2012,48(8):747-758.
[68] JIANG X,LIU Y,LI W,et al.Tissue specific,development dependent phenolic compounds accumulation profile and gene expression pattern in tea plant[Camellia sinensis][J].PLoS One,2013,8(4):1-14.
[69] AHARONI A,DE VOS C H R,WEIN M,et al.The strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in transgenic tobacco[J].Plant journal,2001,28(3):319-332.
