犬瘟热病毒N基因克隆及生物信息学分析
2021-08-06郑庆礼潘书磊刘秉珲福建农业职业技术学院动物科学学院福州350119
郑庆礼 潘书磊 刘秉珲 福建农业职业技术学院动物科学学院 福州 350119
犬瘟热病毒(Canine distemper virus,CDV)可感染多种动物,可导致80%病死率[1]。该病毒可感染犬科类、鼬科和浣熊类动物,近年来发现大熊猫也可感染[2]。犬瘟热主要发生于5月龄以下,临床症状表现复杂多样,主要症状包括白细胞减少、双相热、咳嗽流鼻涕和腹泻便秘等,某些病例还可能会出现共济失调、肌肉强直等神经症状[3-4]。CDV是副粘膜病毒科RNA病毒,全基因组大约包含16689 bp,存在6个结构蛋白,其中N基因编码的N蛋白已被证明在疾病的转录和复制过程中起着十分关健的作用,同时是比较保守的免疫原性蛋白,可以有效诱导机体产生中和抗体[5-7]。目前还未见有人对CDV的N基因进行清晰的生物信息学分析,因此本文通过临床病料分离CDV,并把该病毒的N基因克隆测序后进行相关生物信息学分析,为今后N基因抗原多表位疫苗的研究提供前期基础。
1 材料与方法
1.1 样品 临床送检犬瘟热病料。
1.2 主要试剂与仪器 主要试剂有病毒RNA提取试剂盒,基因克隆试剂盒,高保真PCR mix(购自北京全式金生物技术有限公司),反转录试剂盒(购自Promega生物科技有限公司)。主要仪器有PCR扩增仪(T100,购自伯乐Bio-Rad),水平电泳系统(DYCZ-24KS,购自北京六一生物科技有限公司)。
1.3 引物的设计与合成 根据NCBI上GenBank公布的CDV核酸序列(AB932517.1),利用Oligo7设计特异性常规引物CDV-Ngene F:atggctagccttctcaagag;CDV-Ngene R:gtagctctctatcattatagacaggag,引物由上海生物工程股份有限公司合成。
1.4 CDV核酸提取及N基因克隆 根据操作说明书提取病料RNA,将RNA反转录成cDNA,以cDNA为模板,合成引物为模板进行PCR扩增,扩增体系见表1。将扩增结果凝胶电泳并拍照保存。
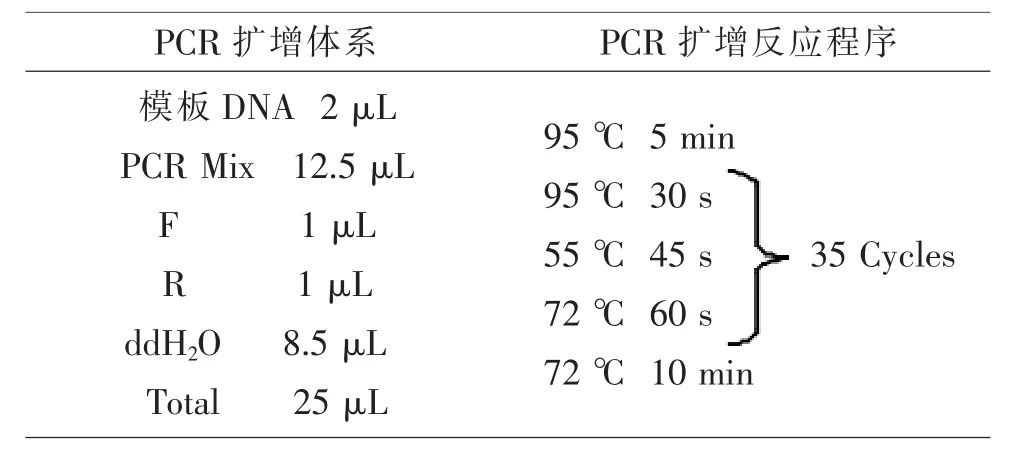
表1 PCR扩增反应条件及体系
对PCR产物根据操作说明进行胶回收纯化,将PCR产物与pEASY-T1克隆载体室进行连接。连接结束后将载体转入DH5a感受态细胞中并置于LB琼脂平板培养过夜。第2 d挑菌进行PCR鉴定,将鉴定正确的PCR产物送上海生物工程技术有限公司测序。
1.5 CDV N基因生物信息学分析 利用DNA Star、DNA MAN对CDV N基因进行遗传进化树和同源性分析;利用DNAMAN分析CDV N基因亲疏水性,跨膜性和抗原位点;利用利用在线软件GOR4预测蛋白质二级结构(https://npsa-prabi.ibcp.fr/cgibin/npsa_automat.Pl page=npsa_gor4.html);利用Net-NG-Lyc1.0和NetPhos 2.0在线分析软件预测N-糖基化位点和磷酸化位点;利用SWISS-MODEL在线软件预测CDV N基因的三级结构(https://swissmodel.expasy.org/interactive)。
2 结 果
2.1 CDV N基因PCR扩增结果 根据凝胶电泳结果显示,在1572 bp左右位置出现明显的特异性条带(见图1),与预期相符合。

图1 CDV基因PCR扩增结果
2.2 CDV N基因遗传进化树及同源性分析 将CDV N基因克隆测序,将测序结果命名为FJ-CDVN gene并与NCBI库的参考序列构建遗传进化树及进行同源性分析,分析结果表明(见图2-图3):测序得到的FJ-CDV-Ngene株与11株参考序列同源在96.9%~99.2%,其中与HM596310.1的同源性最高,同源性为99.2%,与MT136724.1的同源性最低,同源性为96.9%。

图2 CDV基因PCR扩增结果

图3 FJ-CDV-Ngene株同源性分析
2.3 CDV N蛋白N-糖基化及磷酸化位点预测利用NetNGlyo1.0在线分析软件对CDV N蛋白氨基酸序列进行分析,分析结果显示该蛋白不存在N-糖基化位点。利用Netphos2.0在线软件分析结果显示(见图4),CDV N基因氨基酸序列存在32个丝氨酸位点,分别位于第3,7,75,78,83,98,122,125,169,191,228,243,290,298,325,328,355,372,377,395,399,414,422,439,465,469,471,489,504,509,512,513位点;存在15个苏氨酸位点,分别位于9,14,115,200,290,272,279,294,347,402,409,434,460,500,508位点;存在5个酪氨酸位点,分别位于199,260,306,311,516位点。
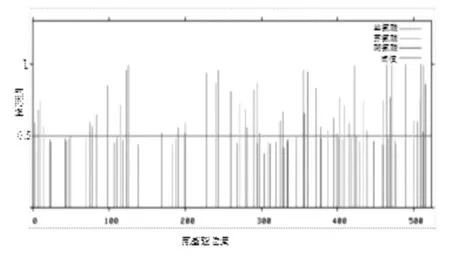
图4 CDVN蛋白磷酸化位点预测
2.4 CDV N蛋白亲疏水性及跨膜结构分析 利用DNAMAN对CDV N蛋白氨基酸序列进行亲疏水性分析,结果显示该蛋白亲水性及疏水性主要分布区间为-4~4,亲水氨基酸明显多于疏水氨基酸,占据了该蛋白的主要成分(见图5-A)。跨膜分析结果显示(见图5-B),N蛋白存在4个跨膜片段,跨膜区域分别为:第29-52aa,多肽长度为24aa;第66-85aa,多肽长度为20aa;第172-196aa,长度为25aa;第249-277aa,长度为29aa。

图5 CDV N蛋白疏水性及跨膜结构分析
2.5 CDV N蛋白二级结构预测 二级结构预测结果显示(见图6),CDV N蛋白氨基酸序列由α-螺旋结构、β-折叠和随机卷曲三种结构组成。其中α-螺旋结构占比为33.84%,β-折叠占16.44%,无规则卷曲结构占49.72%。

图6 CDV N蛋白二级结构预测
2.6 CDV N蛋白三级结构预测 在SWIS-MODEL数据库中匹配到CDV N蛋白的模板(见图7-A),CDV N蛋白与模型蛋白的同源性为54%(见图7-B),N蛋白的QMEAN为-1.69(见图7-C)。CDV N蛋白氨基酸与模型氨基酸的对应序列见图7-D,从建模结果得知CDV N蛋白的三维结构与预测结果基本一致。

图7 CDV N蛋白三级结构预测
2.7 CDV N蛋白抗原位点分析 利用DNAMAN对CDV N蛋白进行抗原位点分析,结果显示该蛋白共含有22个抗原位点,最大分值为1.225(见表2)。

表2 CDV N蛋白抗原位点分析
3 讨论
生物信息学分析是利用计算机对基因编码的蛋白结构进行预测,是现代分子生物学重要的研究方法之一。CDV N基因编码的N蛋白具有良好免疫原性,能够诱导机体产生细胞免疫和体液免疫。N基因同源性分析发现。经亲疏水性预测,发现N蛋白亲水作用较强,因此推测该蛋白可溶于水中。蛋白质磷酸化修饰与DNA损伤修复、转录、信号传导、细胞凋亡等生物学过程相关,通过研究蛋白质磷酸化可阐述多种生物学过程机理[8]。磷酸化主要指肽链中有丝氨酸、苏氨酸、酪氨酸的侧链被修饰,经过生物学软件分析可知N蛋白存在32个丝氨酸位点,15个苏氨酸位点,5个酪氨酸位点,为今后研究该蛋白的相关生物学过程机理奠定前期基础。糖基化能改变多肽构象,因而增加蛋白质的稳定性[9]。对N蛋白进行生物信息学分析,发现该蛋白不存在糖基化位点,无糖基化位点对该蛋白稳定的影响还有待进一步研究。通过对N蛋白的跨膜域分析显示存在4个跨膜域,推测该蛋白有可能作为膜受体起作用或离子通道蛋白。蛋白质不同的二级、三级结构对其功能起着至关重要作用,通过对N蛋白分析发现该蛋白二级结构主要是由无规则卷曲构成,并在数据库中找到与该蛋白相似的模型结构。抗原肽一般指具有免疫原性的多肽或抗原衍生肽,对动物免疫功能具有重要意义[10-11]。经分析,CDV N蛋白有22个抗原位点,具有丰富的抗原位点,为今后多抗原表位筛选做好前期工作基础。
本研究通过对CDV N基因进行生物学分析,为进一步对CDV N相关功能及机理的阐释及研究提供前期理论基础。
