冻融和冻融+苏打盐碱复合胁迫下苜蓿幼苗的生理响应
2021-08-03方志坚冯成龙金姗姗王思琦石婉莹
李 红, 李 波, 方志坚, 杨 曌, 张 超, 冯成龙, 金姗姗, 王思琦, 石婉莹
(1.黑龙江省农业科学院畜牧兽医分院, 黑龙江 齐齐哈尔161005; 2.齐齐哈尔大学生命科学与农林学院, 抗性基因工程与寒地生物多样性保护黑龙江省重点实验室, 黑龙江 齐齐哈尔 161006; 3.齐齐哈尔市园艺研究所, 黑龙江 齐齐哈尔, 161006)
我国东北地区存在大面积的以碱土为主的盐碱地[1],同时春秋两季冻融频繁发生,冻融和盐碱复合环境胁迫对牧草的生长、生理特性的影响比单一胁迫更大[2-3]。近年来,对苜蓿(MedicagoSativaL.)耐盐性和低温的研究取得一定成果,苜蓿幼苗阶段是其耐盐碱和低温能力较弱的时期,也是对苜蓿材料进行耐盐性和低温筛选的关键时期。针对单盐和混合盐碱的生理生化研究表明,4种单盐(NaCl,Na2SO4,NaHCO3和Na2CO3)对苜蓿胁迫下,随着盐浓度增加,幼苗的脯氨酸(Proline,Pro)、可溶性糖(Sduble sugar,SS)和丙二醛(Malondialdehyde,MDA)含量增加,碱性盐胁迫各指标变化大于中性盐[5];对NaCl/Na2CO3胁迫苜蓿幼苗抗氧化酶活性和MDA含量的研究可知,苜蓿叶片超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化物酶(Peroxidase,POD)、过氧化氢酶(Catalase,CAT))活性及MDA含量呈增加趋势[5]。有关低温对苜蓿生理影响的研究也取得了一定成果,在紫花苜蓿低温胁迫生理特性研究中发现,SOD活性在4℃低温12 h内升高,24 h后下降,Pro含量随低温延长而增加,叶片电导率在温度高于—3℃时增加缓慢,—4℃时显著增加,说明苜蓿能够耐受—3℃的低温[6]。
目前,针对牧草的耐寒和耐盐碱能力研究主要集中于单因素胁迫条件,有关冻融与盐碱复合胁迫下牧草的响应机制研究甚少。苜蓿幼苗在冻融和碱性盐胁迫的生理反应发现,在中度碱性盐胁迫下,体内活性氧、MDA含量和CAT,SOD活性升高,蛋白含量降低,整个冻融过程中,Pro和MDA含量、CAT和SOD活性呈先升后降[7]。在冻融和碱性盐双重胁迫下,苜蓿幼苗的损伤更大,关于苜蓿对冻融和盐碱胁迫的响应已受到广泛关注,但温度进程和复合盐碱浓度控制及对苜蓿生理代谢影响的综合分析仍有待完善。
紫花苜蓿因其具有适应性广、产量高、抗寒、抗旱、耐盐碱、固氮能力强等特点,在国内外广泛种植,是全球范围内极具栽培价值的植物之一[8],但低温和盐碱可严重制约着苜蓿产量和畜牧业的发展,因此对苜蓿幼苗进行冻融锻炼能有效提高苜蓿的抗寒性。本研究以‘龙牧807’苜蓿为材料,探究冻融及冻融+盐碱复合胁迫对苜蓿幼苗生理生化的影响,为优质牧草选育及恢复寒性地区盐渍化土壤奠定基础。
1 材料与方法
1.1 试验材料
以抗苏打盐碱能力较强的‘龙牧807’苜蓿(MedicagoSativaL.‘Longmu 807’)种子为试验材料,该材料由黑龙江省农业科学院畜牧兽医分院所提供。
1.2 幼苗的培养
2019年5月15日将供试材料播种在土培育苗钵(高13 cm,下径14 cm,上径16 cm)中,培养基质为黑土∶针叶土=5∶1,每钵均匀播种30粒种子,覆盖1 cm厚培养基质,在25℃的培养室中培养,每2 d供水一次。培养至 45 d后苗高 10~15 cm,可用于胁迫处理。
1.3 冻融胁迫处理
对培养 45 d后的幼苗进行间苗处理,每个营养钵中保留 10 株长势均匀的苜蓿幼苗,并进行冻融胁迫处理,处理温度融冻(T-F)试验为每 2 h温度降低5℃或3℃,同样冻融(F-T)试验为每2 h温度升高5℃或3℃,整个循环试验的温度设计梯度为10℃(T-F10),5℃(T-F5),0℃(T-F0),—3℃,0℃(F-T0),5℃(F-T5),10℃(F-T10),每2 h对变温冰箱的温度进行调节,3次重复,以常温未胁迫处理的幼苗为对照(CK),将冻融和融冻处理的幼苗叶片取下,置于—80℃冰箱中备用。
1.4 冻融+苏打盐碱胁迫处理
对培养 45 d后的苜蓿幼苗进行间苗处理,每个营养钵中保留 10 株长势均匀的幼苗,每个营养钵中浇灌150 mmol·L-1的混合苏打盐碱溶液100 mL(NaHCO3∶Na2CO3=9∶1),处理 2 d,然后进行同1.3冻融胁迫处理,每个处理3次重复,将冻融+苏打盐碱胁迫的幼苗叶片取下,置于-80℃冰箱中备用。
1.5 苜蓿幼苗叶片各指标的测定方法
采用考马斯亮蓝法测定可溶性蛋白(Soluble protein,SP)含量;SS含量采用蒽酮比色法测定;Pro含量采用酸性茚三酮法[9]测定;POD活性采用愈创木酚比色法测定;CAT、抗坏血酸过氧化物酶(Aseorbate peroxidase,APX)活性采用分光光度法测定;SOD活性采用NBT光化还原法[11]测定;相对电导率(Relative conductivity,RC)采用电导法测定;MDA含量采用硫代巴比妥酸法[10]测定。
1.6 数据的处理与分析
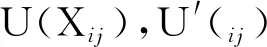
灰色系统的建立:
(1)灰色系统的建立:将各抗逆指标的平均隶属值作为参考序列X0,可溶性糖等9项抗性指标的平均值为比较序列(子序列),记为Xi(i=1,2,…,9)。
(2)数据标准化处理:由于各指标的量纲不一致,需对原始数据作“初值化”处理,根据公式(1)将数据无量纲化:
(1)
式中Xi(k)为原始数据,Xi′(k)为无量纲化后的数据,X和Si分别为同一指标的平均值和标准差。
(3)计算关联系数及关联度:根据公式(2)计算参考数列X0与比较数列Xi各对应值的关联系数:
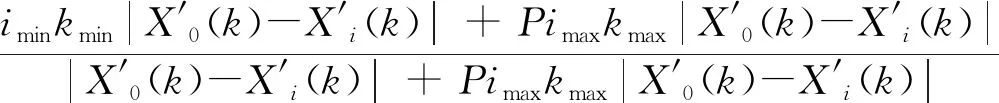
(2)
式中i为某个指标,ξi(k)为比较数列Xi对参考数列X0在第k点的关联系数,p为分辨系数,p∈ (0,1],本文分辨系数取p=0.5。
(4)根据关联系数计算关联度,公式为:
(3)
式中r0i为各序列关联度,i=1,2……,9。
2 结果与分析
2.1 冻融及冻融+盐碱胁迫对苜蓿幼苗渗透调节能力的影响
冻融处理下,苜蓿叶片中SS,Pro和SP含量在整个冻融循环(T-F10至F-T10)变化中呈先升后降的变化趋势(图1),在-3℃低温时3种渗透调节物质含量最高,SS,Pro和SP含量分别比CK高129.46%,329.63%和57.90%。-3,F-T0,F-T5与CK处理SS含量差异显著(P<0.05);冻融胁迫组Pro含量与CK处理差异显著(P<0.05);除T-F10胁迫组SP含量与CK处理差异不显著外,其余各胁迫组与CK处理差异显著(P<0.05)。冻融+盐碱胁迫处理下,苜蓿叶片中的SS,Pro和SP含量在整个循环中呈先升后降的变化趋势(图1)。在-3℃+盐碱胁迫组3种渗透调节物质含量最高,SS,Pro和SP含量分别比CK高289.56%,344.86%和97.34%。冻融+盐碱胁迫下各胁迫组的3种渗透调节物质含量均与CK差异显著(P<0.05)。
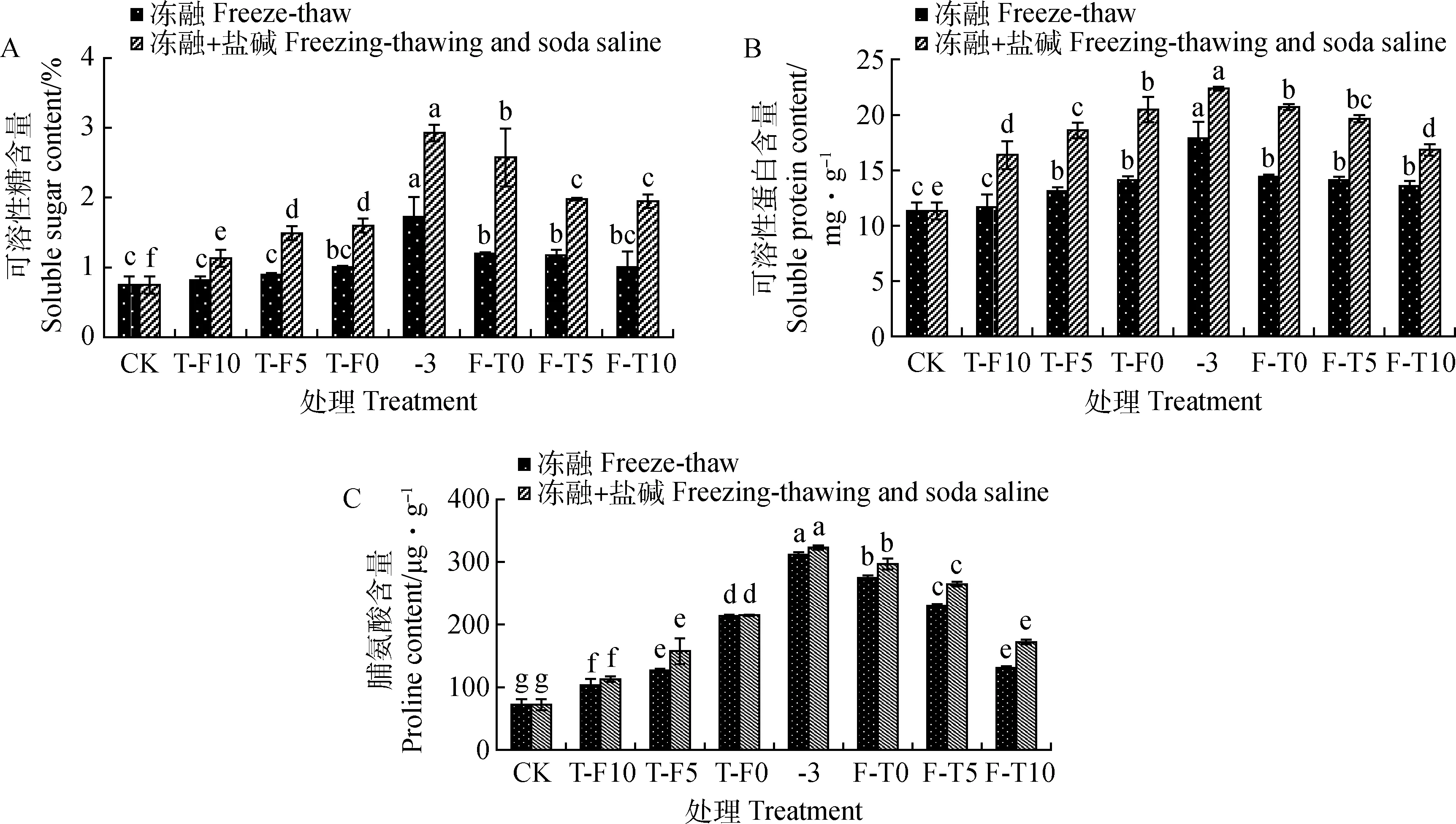
图1 冻融及冻融+盐碱胁迫下苜蓿叶片渗透调节物质的变化
对3种渗透调节物质含量变化进行分析表明,冻融过程(F-T0至F-T10)中SS含量高于融冻过程(T-F10至T-F0),冻融+盐碱各胁迫组的SS,Pro和SP含量高于对应冻融各胁迫组,SS含量分别为冻融各胁迫组的1.39,1.67,1.60,1.70,2.14,1.68和1.94倍,Pro含量分别为冻融各胁迫组的1.09,1.24,1.00,1.04,1.08,1.15和1.31倍,SP含量分别为冻融各胁迫组的1.40,1.42,1.45,1.25,1.43,1.39和1.24倍。冻融及冻融+盐碱胁迫处理下,融冻过程中各胁迫组SS含量变化量较冻融过程大,说明3种渗透调节物质变化在胁迫初期更敏感。
2.2 冻融及冻融+盐碱胁迫对苜蓿幼苗叶片抗氧化酶活性的影响
2.2.1POD和SOD活性 冻融胁迫处理下,苜蓿叶片中POD和SOD活性在整个冻融循环中呈先升后降的变化趋势(图2A,B),在—3℃低温时POD和SOD活性最高,分别较CK增加了32.16%和101.87%(P<0.05)。整个冻融循环中,除F-T处理胁迫组的POD活性比CK低2.16%外,其余各胁迫组均比CK处理高。冻融+盐碱胁迫处理下,苜蓿叶片中的中POD和SOD活性在整个循环中呈先升后降的变化趋势(图2A,B),在—3℃+盐碱胁迫组时达到最高值,分别较CK增加了34.86%和111.76%(P<0.05)。-3℃+盐碱、F-T0+盐碱胁迫组的POD活性与CK处理差异显著(P<0.05),冻融+盐碱胁迫组的SOD活性均与CK处理差异显著(P<0.05)。
对POD和SOD活性变化进行分析表明,冻融过程中POD和SOD活性高于融冻过程,冻融+盐碱各胁迫组的POD和SOD活性高于对应冻融各胁迫组,冻融+盐碱各胁迫组的POD活性分别为冻融各胁迫组POD活性的1.01,1.02,1.02,1.02,2.03,1.02和1.08倍,SOD活性分别为冻融各胁迫组SOD活性的1.04,1.04,1.03,1.05,1.01,1.01和1.03倍。
2.2.2CAT和APX活性 冻融胁迫处理下,CAT和APX活性在整个冻融循环变化过程中呈逐渐上升的变化趋势(图2C,D),F-T10处理CAT和APX活性最高,分别较CK增加了262.38%和134.00%。除T-F10和T-F5处理外,冻融胁迫组CAT和APX活性与CK处理差异显著(P < 0.05)。在冻融+盐碱胁迫处理中,苜蓿叶片中CAT和APX活性整个循环中呈逐渐升高的变化趋势(图2C,D),F-T10+盐碱胁迫组CAT和APX活性最高,分别较CK增加了292.08%和184.00%。冻融+盐碱胁迫下各胁迫组的CAT和APX活性均与CK处理差异显著(P< 0.05)。

图2 冻融及冻融+盐碱胁迫下苜蓿叶片抗氧化酶活性的变化
对CAT和APX活性变化进行分析表明,冻融+盐碱胁迫下各胁迫组的CAT和APX活性均高于冻融循环各胁迫组,冻融+盐碱各胁迫组CAT活性分别为冻融各胁迫组的2.04,2.04,1.10,1.09,1.08,1.04和1.08倍,APX活性分别为冻融各胁迫组的1.47,1.54,1.35,1.30,1.29,1.30和1.21倍。
2.3 冻融及冻融+盐碱胁迫对苜蓿幼苗叶片膜透性物质的影响
冻融胁迫处理下,苜蓿叶片中MDA含量在整个冻融循环中呈先升后降的变化趋势(图3A),在F-T0胁迫组MDA含量最高,较CK增加了202.89%;RC在整个冻融循环中呈逐渐上升的变化趋势(图3B),F-T10处理时RC最高,较CK增加了74.54%。冻融循环各胁迫组MDA和RC与CK处理差异显著(P<0.05)。

图3 冻融及冻融+盐碱胁迫下苜蓿叶片膜透性物质的变化
冻融+盐碱胁迫处理下,苜蓿叶片中的MDA含量在整个循环中呈先升后降的变化趋势,在F-T0+盐碱胁迫时达到最高值,较CK增加了291.88%;RC在整个循环变化过程中呈逐渐上升的变化趋势,在F-T10+盐碱胁迫时达到最高值,较CK增加了82.93%。冻融+盐碱各胁迫组MDA和RC与CK处理差异显著(P<0.05)。
对MDA含量和RC变化进行分析表明,冻融过程中MDA含量和RC高于融冻过程,冻融+盐碱各胁迫组的MDA含量和RC高于对应冻融各胁迫组,冻融+盐碱各胁迫组的MDA含量分别为冻融各胁迫组MDA含量的1.78,1.50,1.13,1.22,1.29,1.08和1.00倍,冻融+盐碱各胁迫组的RC分别为冻融各胁迫组的1.15,1.13,1.13,1.14,1.19,1.16和1.05倍。
2.4 冻融和冻融+盐碱胁迫下苜蓿幼苗叶片各指标隶属函数分析
应用隶属法对冻融及冻融+盐碱胁迫下各生理生化指标进行分析(表1)表明,冻融胁迫下各处理隶属值由高到低依次为:—3℃>F-T0>F-T10> F-T5> T-F0>T-F5>T-F10>CK,冻融+盐碱胁迫下各处理隶属值由高到低依次为:-3℃+盐碱>F-T0+盐碱>F-T10+盐碱>F-T5+盐碱>T-F0+盐碱>CK+盐碱>T-F5+盐碱>T-F10+盐碱>CK。隶属函数综合评价值越大,表明该处理下苜蓿幼苗渗透调节物质积累越多、抗氧化酶活性越高及对膜损伤越小。

表1 冻融及冻融+盐碱胁迫下各指标的隶属值
2.5 灰色关联度分析
关联系数大小代表各指标对该处理的敏感程度,对冻融及冻融+盐碱胁迫下苜蓿幼苗叶片9项指标进行灰色关联分析,根据不同胁迫下各指标的测定值计算关联度(表2)。结果表明,冻融胁迫下,SS,Pro,SOD和CAT具有较大的关联系数,而SP关联系数最小,说明SS,Pro,SOD和CAT指标对冻融胁迫最敏感,其余指标敏感度较低。冻融+盐碱胁迫下,Pro,SOD和SP指标关联系数较大,而RC关联系数最小,说明Pro,SOD和SP指标对冻融+盐碱胁迫最敏感。综合分析表明,各指标与冻融和盐碱胁迫的关联度由高到低依次为:Pro>SOD>CAT>SS>APX>SP>MDA>POD>RC。

表2 不同胁迫处理下各指标的关联度
3 讨论
冻融和混合盐碱胁迫是限制植株生长发育的重要因素,冻融胁迫引起植物细胞内水分结冰,植株无法利用结冰的水分,导致植物体内水分短缺;混合盐碱胁迫下,植株生存环境中的盐离子含量较高,外环境渗透压升高,导致植物体渗透失水[11-12]。本研究在冻融及冻融+盐碱胁迫下,整个冻融循环变化过程中Pro,SS和SP含量等均呈先增后降变化趋势,在—3℃低温胁迫时各渗透调节物质含量最高,冻融和盐碱双重因子胁迫下3种渗透调节物质含量明显高于单因子胁迫,说明冻融条件下的盐碱环境对苜蓿幼苗的伤害更大。这是因为在双重胁迫下,苜蓿不仅要通过渗透调节物质调节其渗透能力,还要抵抗低温伤害,使生长受到抑制。本研究结果与巩泽等[13]研究黑麦草(LoliumperenneL.)幼苗在冻融及NaHCO3复合胁迫下的渗透调节物质变化结果一致。植物在遭受冻融和盐碱等胁迫时,体内积累了大量Pro,SS和SP等渗透调节物质,以降低冻融和盐碱对植物造成伤害,其积累指数与植物抗逆性密切相关。
逆境胁迫下,植物细胞膜损伤和通透性加大,导致电解质外泄,膜质过氧化增加,产生大量MDA[14-15]。本试验结果表明,冻融及冻融+盐碱胁迫下,整个冻融循环变化过程中RC呈逐渐增加的变化趋势,且冻融过程RC的变化幅度较融冻过程低,而MDA含量在整个冻融循环变化过程中呈先增后降的变化趋势,说明在冻到融过程中,随着温度回升,电解质渗透量减小,植物体通过自我调节机制进行质膜修复,增强质膜的选择透性,降低膜质过氧化程度,导致MDA含量下降。而冻融+苏打盐碱双重胁迫,植物所受到的膜损伤和膜质过氧化和增加,导致该循环变化过程中RC和MDA含量高于对应冻融单因子胁迫,本研究结果与李晨等[16]人研究黑麦草幼苗对冻融及酸沉降的生理响应特征时得到结论相似。MDA含量和RC的变化反映了逆境对植物的伤害程度,在冻融和盐碱逆境胁迫时,通过积累渗透调节物质可减少膜脂过氧化和膜的损伤。
植物体正常生长环境中,活性氧(ROS)的产生和清除速率保持动态平衡,逆境胁迫下,ROS的产生速率增加,植物体可激活体内的抗氧化酶(POD,SOD,CAT,APX)活性来清除过量的ROS,以抵御逆境胁迫带来伤害[17-19]。本研究中,冻融及冻融+盐碱胁迫下,POD和SOD活性在整个冻融循环变化过程中均呈先增后降的趋势,CAT和APX活性在整个冻融循环变化过程中则呈逐渐增加的趋势,冻融和盐碱双因子共同胁迫,导致苜蓿叶片的抗氧化酶活性均高于单因子胁迫,研究结果与何访淋等[20]人研究醋酸钙镁盐环保融雪剂及冻融胁迫对高羊茅幼苗的生理影响,朱悦等[22]人研究白三叶叶片对酸沉降及冻融胁迫的生理响应特征的研究结果一致。在逆境条件胁迫下,植物通过提高抗氧化酶系统的活性来清除过量ROS,以抵抗冻融及盐碱胁迫导致的植物机体损伤。
苜蓿对冻融和冻融+盐碱胁迫的生理生化变化过程是一种综合反应,苜蓿对盐胁迫和冻融胁迫表现出一定的抗性,对紫花苜蓿寒冷地区盐碱的确切机理有待进一步研究。本试验仅从生理生化指标方面探讨了其对苗期苜蓿的影响,应进一步探讨这种复合胁迫对苜蓿形态结构和基因差异表达的影响。
4 结论
苜蓿幼苗在冻融循环和冻融循环+苏打盐碱胁迫下,通过提高抗氧化酶活性与增加渗透调节物质的含量,可缓解胁迫导致的伤害。隶属函数法对冻融循环、冻融循环和苏打盐碱胁迫下9项生理生化指标变化分析,冻融循环胁迫下隶属值由高到低依次为—3℃>F-T0>F-T10> F-T5> T-F0>T-F5>T-F10> CK;冻融循环+盐碱胁迫下隶属值由高到低依次为—3℃+盐碱>F-T0+盐碱>F-T10+盐碱>F-T5+盐碱>T-F0+盐碱>CK+盐碱>T-F5+盐碱>T-F10+盐碱>CK;冻融循环和冻融循环+盐碱胁迫与各指标综合关联度由高到低依次为Pro>SOD>CAT>SS>APX>SP>MDA>POD>RC。
