胶州湾大型底栖动物群落的年次级生产力及其影响因素的研究*
2021-07-27杨湘君崔雯瑶张蒙生于子山
陈 晨, 杨湘君, 崔雯瑶, 张蒙生, 于子山**
(1. 中国海洋大学海洋生命学院,山东 青岛 266003; 2. 海昌(中国)投资有限公司, 上海 200050; 3. 上海勘测设计研究院有限公司, 上海 200050)
大型底栖动物在海洋生态系统的物质循环和能量流动中有着举足轻重的作用,在食物网中,它主要承担着消费者和分解者的功能[1-2]。次级生产力是指动物在生长和繁殖过程中增加的生物量或存储的能量[3]。研究大型底栖动物群落的次级生产力,可以了解大型底栖动物在生态系统的物质循环和能量流动过程中所起的作用,对海洋底栖生态系统的深入研究以及海洋生物资源的合理利用,也有一定帮助[4-6]。
国外学者对于大型底栖动物次级生产力的研究可追溯到1919年,Boysen-Jensen 对莱姆峡湾单个种群生产力的计算[1]。1970年代后,关于大型底栖动物种群次级生产力的研究逐渐增多,采用的是同生群法和非同生群法,这些方法结果较为准确,但野外采样工作量大,计算过程也十分复杂[7-8]。有鉴于此,Robertson根据Mann等[9]对P/B值的研究成果,提出了估算大型底栖动物种群次级生产力的回归方程。之后,Schwinghamer[10]和Edgar[11]又分别提出了大型底栖动物种群次级生产力与个体体重关系的估算公式。1990年代后,Brey[5]相继提出了Brey(1990)、Brey(2001)和Brey(2012)等大型底栖动物种群次级生产力与个体重量、生境类型、食性及移动方式等关系的公式[12]。以上这些公式,可以使人们依据比较简单的野外调查数据,就能估算出种群的次级生产力。
国内对于大型底栖动物种群次级生产力的研究起步较晚且较少,目前仅有吴宝玲等、周一兵等、林岿璇等和闫云君等[13-16],报道了多齿围沙蚕(Perinereisnuntia)、日本刺沙蚕(Neanthesjaponica)、小头虫(Capitellacapitata)、幽蚊(Chaoborussp.)和圆扁螺(Hippeutissp.)的次级生产力,采用了同生群法和非同生群法。
1990年代起,随着海洋生态动力学研究的深入,人们需要定量了解作为食物网中重要一环的底栖动物,在能流和物流中所起的作用,如生产量等,亦即整个底栖动物群落的生产力。而关于群落的次级生产力,没有现成的估算方法。于子山等[17]首次提出了估算大型底栖动物群落年次级生产力的简单方法,就是将野外获得的大型动物的年平均丰度和年平均生物量数据,利用Brey(1990)模型,在每站逐种计算年次级生产力,然后将该站位出现的所有种的次级生产力相加,得到该站位群落的年次级生产力,由此得到整个渤海的大型底栖动物年次级生产力为6.49 g·m-2·a-1。此后,多位学者[3,8,18-33]用此逐种累加的方法,使用Brey(1990)、Brey(2001)和Brey(2012)等模型,展开了对胶州湾、莱州湾、乳山湾、海州湾和大亚湾等海域大型底栖动物次级生产力的研究。
胶州湾是中国北方典型的半封闭海湾,湾周围河流提供了丰富的营养盐[18],使湾内拥有非常高的生物多样性和生产力,是中国开展海洋生态学研究最早的海湾之一。关于胶州湾大型底栖动物群落次级生产力的研究始于2005年,张崇良等[8]利用Brey(1990)模型,研究了胶州湾西北部潮间带大型底栖动物的次级生产力;袁伟等[20]利用Brey(1990)模型,研究了胶州湾西部海域大型底栖动物的次级生产力;刘旭东等[32]基于春夏秋三个季节的资料,用Brey(1990)、Brey(2001)和Brey(2012)模型,分别计算了胶州湾大型底栖动物次级生产力,得出Brey(2001)模型与Brey(2012)模型计算结果差异不显著的结论;李新正等[18]和王金宝等[22]利用Brey(1990)模型,研究了1998—2004和2005—2009年间,胶州湾大型底栖动物次级生产力长期的变化;全秋梅等[33]利用Brey(1990)模型,对比了胶州湾内外大型底栖动物次级生产力的差别。上述研究为胶州湾大型底栖动物次级生产力的研究,奠定了基础。但这些研究多使用的是最原始的Brey(1990)模型;或因为站位设置不够多,导致未能覆盖整个胶州湾海域,如研究中均未涉及胶州湾的牡蛎养殖在胶州湾大型底栖动物次级生产力中的贡献等,因此有必要对胶州湾大型底栖动物群落的次级生产力进行更深入的研究。另外,胶州湾大型底栖动物的群落次级生产力,受哪些环境因素的影响?目前仅见全秋梅等[33]给出的胶州湾大型底栖动物的次级生产力与环境因子的相关程度均不显著的报道,这方面的研究也有待继续深入。
本研究根据2018年春夏秋冬四个季节在胶州湾海域均匀设置的20个站位的调查资料,利用Brey(2001)模型,估算胶州湾大型底栖动物群落年次级生产力,分析养殖贝类在胶州湾大型底栖动物次级生产力中的贡献,探讨大型底栖动物群落年次级生产力与沉积物粒度、有机质、叶绿素及温盐深等环境因子的关系,并评估大型底栖动物群落年次级生产力与海域的环境承载量,为胶州湾底栖生态系统动力学的深入研究,以及底栖动物资源的持续利用,提供参考。
1 材料和方法
1.1 调查海域
于2018年1月(冬季)、5月(春季)、9月(夏季)、11月(秋季)对胶州湾海域20个站位的大型底栖动物调查取样,调查船为“天使1号”,站位分布如图1所示。

图1 胶州湾海域大型底栖动物采样站位
1.2 样品采集与处理
使用采样面积为0.05 m2的HNM箱式采泥器采集样品,每站采样3次,合为一个样品。现场分选沉积物泥样所用网筛孔径为0.5 mm,留在筛子上的标本和渣滓全部装样品瓶,5%福尔马林溶液固定。每站取一定量的表层沉积物,-20 ℃冷冻保存,用于沉积物粒度、有机质和叶绿素的测定。使用船载CTD仪现场测定各站位的水深、底温和底盐。
在实验室对大型底栖动物进行分选、鉴定、计数和称重。其中管栖多毛类去管称重,软体动物、寄居蟹去壳称重,称重(湿重)使用的电子天平感量为 0.000 1 g。野外及实验室内样品的采集和处理,均按《海洋调查规范:第6部分海洋生物调查》[34]进行。
沉积物中有机质含量的测定采用重铬酸钾-硫酸氧化法[35]。沉积物粒度利用Cilas940L型激光粒度仪测定,部分站位因底质颗粒较大采用筛析法[35]。沉积物中叶绿素a(Chla)和脱镁叶绿素a(Pha)含量的测定采用荧光分光光度法[34]。
1.3 数据处理
利用相对重要性指数(IRI)[36]确定胶州湾大型底栖动物优势种。计算公式为:
IRI=(W+N)×F。
式中:W为物种生物量占总生物量的百分比;N为物种丰度占总丰度的百分比;F为物种出现频率。
Brey(2001)模型[37]:
Lg(P/B)=7.947-2.294 lg(M)-
2 409.856/(T+273)+0.168/D+
0.194SubT+0.180InEpi+0.277MoEpi+
0.174Taxon1-0.188Taxon2+0.33Taxon3-
0.062Habitat1+582.851 lg(M)/(T+273)
式中:P为大型底栖动物年均次级生产力,kJ·m-2·a-1;M为大型底栖动物年平均个体体重能值,kJ;T为调查海域平均底温,℃;D为调查海域平均水深,m;B为大型底栖动物年平均生物量能值,kJ·m-2·a-1;SubT为栖息地类型:潮下带值为1,潮间带为0;InEpi为栖息方式:底内动物值为1,底表动物为0;MoEpi为移动方式:移动动物值为1,固着动物为0;Taxon为动物类群,Taxon1:多毛类或甲壳类值为1,其它类群为0;Taxon2:棘皮动物值为1,其它类群为0;Taxon3:昆虫值为1,其它类群为0;Habitat1为生境类型:湖泊生境值为1,其它生境值为0。
该模型需要将湿重数据转换为能值,但是为方便后续与历史资料比较,还需要将模型的结果转换为去灰分干重,转换关系见表1[7,24]。

表1 Brey(2001)模型中各参数转换关系
本研究将2018年四个季节所采到的每种大型底栖动物丰度和生物量取平均值,作为年平均丰度和年平均生物量,利用Brey(2001)模型估算各站位每个物种的年次级生产力,然后将这些计算结果相加,得到每个站位大型底栖动物群落年次级生产力。本文中生物量和次级生产力结果均为去灰分干重结果。
使用SPSS 22.0软件进行次级生产力与环境因子的Pearson相关分析。
2 结果
2.1 胶州湾大型底栖动物种类组成和优势种
本研究共采集大型底栖动物354种,其中多毛类124种,甲壳类119种,软体动物87种,棘皮动物8种,其它类群16种,多毛类、甲壳类和软体动物的物种数占总物种数的93.2%。冬、春季物种数最多,夏季次之,秋季最少;各季节大型底栖动物的物种组成类似,都是多毛类物种数最多,甲壳类、软体动物和棘皮动物次之(见表2)。

表2 不同季节胶州湾大型底栖动物物种组成及其物种数
本研究四个季节的大型底栖动物优势种如表3所示,优势种以多毛类和软体动物为主,其中,菲律宾蛤仔R.philippinarum在四个季节中均为第一优势种,且相对重要性指数明显高于其它优势种。
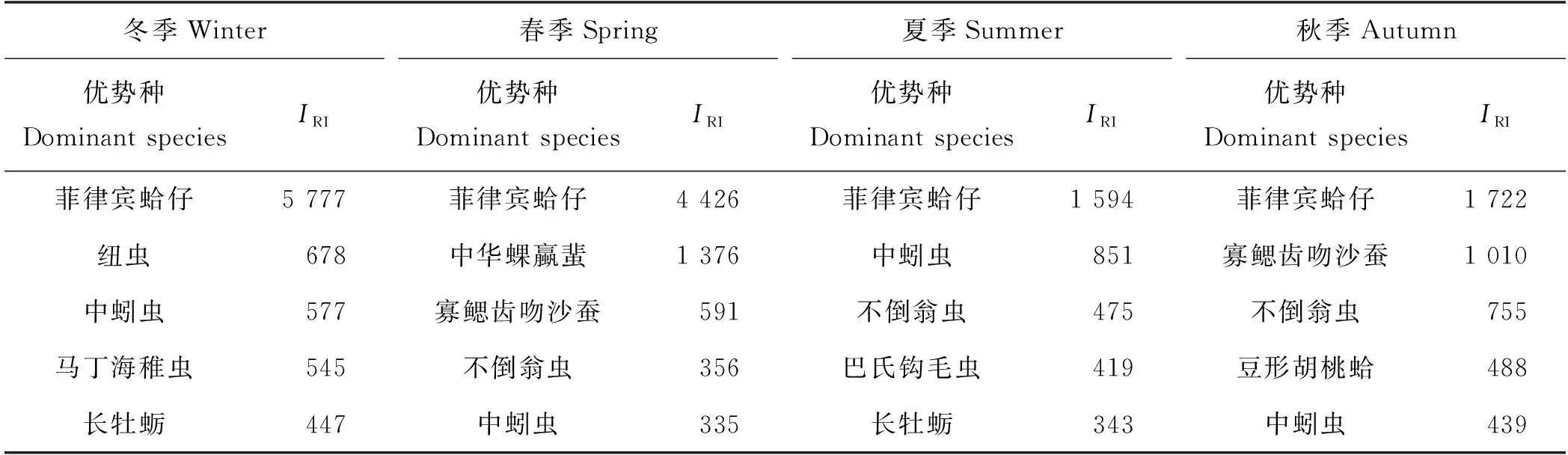
表3 不同季节胶州湾大型底栖动物优势种
2.2 胶州湾大型底栖动物的丰度、生物量和年次级生产力
胶州湾各站位大型底栖动物的丰度、生物量和年次级生产力的结果如表4所示。
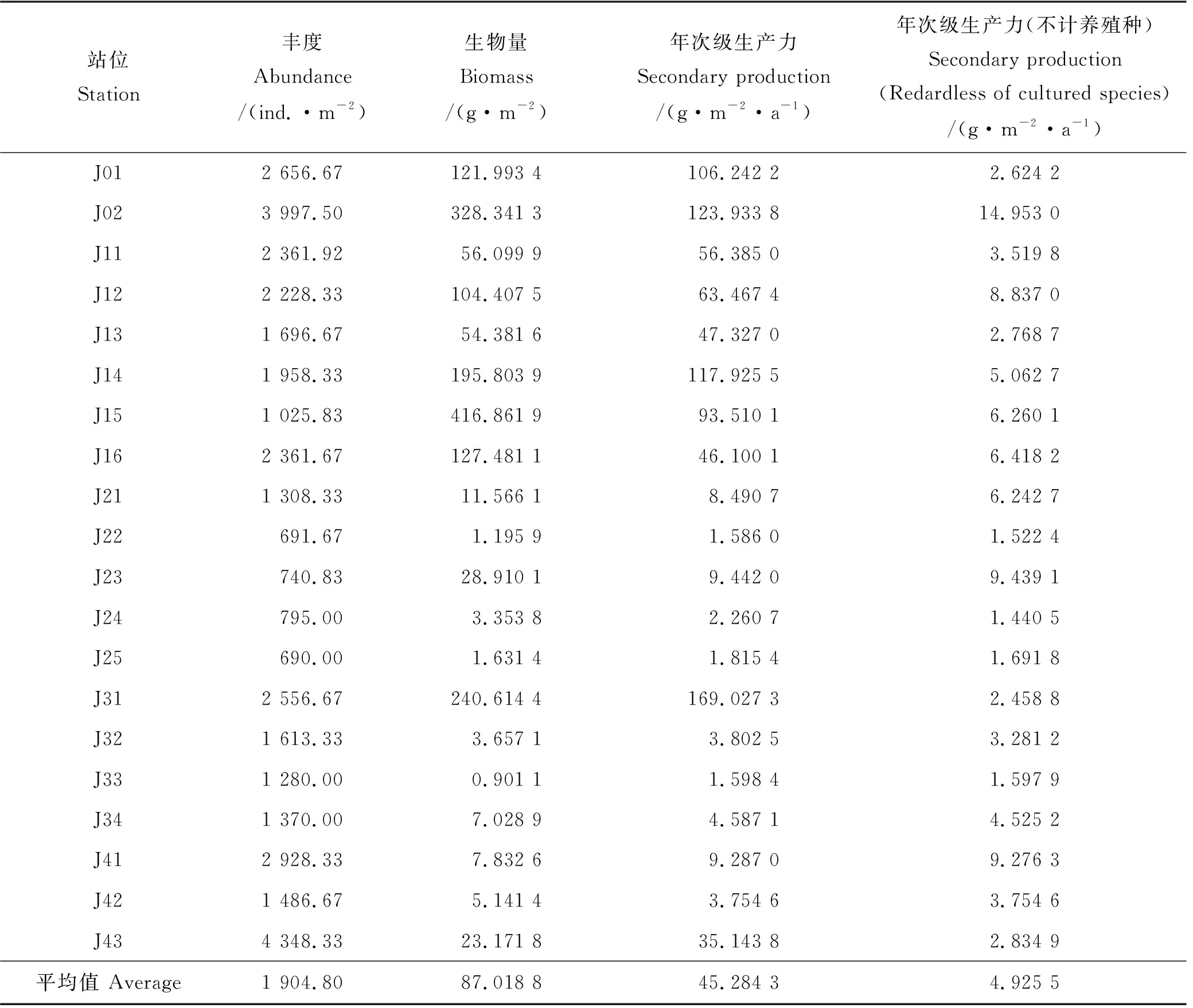
表4 胶州湾各站位大型底栖动物丰度、生物量和年次级生产力
胶州湾大型底栖动物年平均丰度为1 904.80 ind./m2,丰度分布呈现出由胶州湾中部向南北两侧增大的趋势(见图2),丰度最高的站位为湾口附近J43(4 348.33 ind./m2),菲律宾蛤仔(R.philippinarum)的丰度占该站位总丰度的61.08%。丰度最低值出现在胶州湾中部的J25站位,该站位大型底栖动物的丰度仅为690.00 ind./m2。

图2 胶州湾大型底栖动物丰度平面分布(丰度单位:ind.·m-2)
胶州湾大型底栖动物年平均生物量为87.018 8 g/m2,生物量分布也呈现出由胶州湾中部向南北两侧增大的趋势(见图3)。软体动物生物量占大型底栖动物生物量的96.60%,长牡蛎(C.gigas)和菲律宾蛤仔的生物量占软体动物生物量的49.96%和41.84%。生物量最高的站位为J15(416.861 9 g·m-2),其次为J02(328.341 3 g·m-2),这两个站位均采集到大量的长牡蛎。生物量最低和次低的站位为J33(0.901 1 g·m-2)和J22(1.195 9 g·m-2),这2个站位均位于胶州湾中部海域。

图3 胶州湾大型底栖动物生物量平面分布(生物量单位:g·m-2)
将20个站位的次级生产力平均,得到胶州湾大型底栖动物群落年次级生产力为45.284 3 g·m-2·a-1,它也呈现出由胶州湾中部向南北两侧增大的趋势(见图4),其最高值的站位为J31(169.027 3 g·m-2·a-1),J02站位次之,为123.933 8 g·m-2·a-1,这两个站位都出现了大量的菲律宾蛤仔。J22站位的年次级生产力最小,仅为1.586 0 g·m-2·a-1,该站位物种组成以多毛类、端足类为主,物种的丰度和生物量均较小。

图4 胶州湾大型底栖动物年次级生产力的平面分布
2.3 胶州湾大型底栖动物次级生产力与环境因子相关性
本研究计养殖种的和不计养殖种的次级生产力与环境因子相关性分析结果见表5、6,由表5可知,计养殖种的胶州湾大型底栖动物群落年次级生产力与底盐呈显著负相关(P<0. 05),与沉积物中叶绿素a呈显著正相关(P<0.05),与其余环境因子无显著相关性。由表6可知,不计养殖种的胶州湾大型底栖动物群落年次级生产力与环境因子无显著相关性。

表5 大型底栖动物年次级生产力与环境因子的相关性
①Parameters;②Secondary production;③Temperature;④Salinity;⑤Depth;⑥Average particle size;⑦Median diameter;⑧Chlorophylla;⑨Pheophytina;⑩Organic matter

表6 不计养殖种的大型底栖动物年次级生产力与环境因子的相关性
3 讨论
3.1 环境因子对胶州湾大型底栖动物次级生产力的影响
本研究胶州湾底盐的平均值为31.06,变化范围为30.15~31.54,位于湾口附近的J33、J41、J42和J43站位的底盐较高,位于湾北部的站位底盐较低,如J02、J16站位等,总体上呈现出由湾口向湾内逐渐降低的趋势。湾口位置,海水交换能力强,底盐维持在较高水平;越靠近湾北部,海水交换能力越弱,再加上陆源淡水的注入,使得底盐逐渐降低。有研究表明[38],底盐通过限制大型底栖动物生理机能,对大型底栖动物群落的物种组成、空间分布以及数量特征产生影响。本研究中,底盐升高,大型底栖动物群落次级生产力下降,这可能是因为较高的底盐会限制大型底栖动物的生长发育繁殖。
有研究报道[21,23],局域尺度上沉积物表层的叶绿素a,是影响大型底栖动物群落次级生产力的重要因素,本文也如此。本研究沉积物中叶绿素a的平均浓度为1.37 μg/g,变化范围为0.52~2.10 μg/g。位于湾中部的J23和J24站位,沉积物中叶绿素a浓度较低,位于湾北部的J02、J13站位和湾口附近的J32、J33站位,沉积物中叶绿素a浓度较高,其总体分布趋势与次级生产力分布趋势基本一致,呈现出由湾中部向南北两侧逐渐增大的趋势。叶绿素a的浓度代表沉积物中底栖微藻生物量的多少,而底栖微藻是贝类重要的饵料,也是许多小型底栖动物的饵料来源。小型底栖动物通过作为多毛类、虾、蟹和幼鱼等大型底栖动物的饵料,影响着大型底栖动物的生长发育繁殖[39-40]。
3.2 胶州湾主要养殖区养殖贝类年次级生产力的估算
本研究的部分站位,分别位于红岛、红石崖和黄岛养殖区,各站位养殖贝类次级生产力见表7。红岛养殖区贝类年次级生产力为73.893 5 g·m-2·a-1;红石崖养殖区养殖贝类的年次级生产力为166.568 6 g·m-2·a-1;黄岛养殖区养殖贝类的年次级生产力为32.308 9 g·m-2·a-1。
本研究中大型底栖动物群落年次级生产力以及养殖贝类的年次级生产力分别为45.284 3和40.358 9 g·m-2·a-1,养殖贝类对次级生产力的贡献率高达89.12%。
如果不计养殖种,本研究大型底栖动物群落次级生产力结果和分布状况如表4、图5所示。大型底栖动物群落年次级生产力仅为4.925 5 g·m-2·a-1,远低于45.284 3 g·m-2·a-1,不计养殖种的胶州湾大型底栖动物次级生产力无明显分布趋势。J02站位的年次级生产力最高,为14.953 0 g·m-2·a-1,对J02站位次级生产力贡献较大的为杂色纹藤壶(Amphibalanusvariegatus)。位于非养殖区的J23和J41站位的年次级生产力次之,对J23站位次级生产力贡献较大的为毛蚶(Scapharcasubcrenata),对J41站位次级生产力贡献较大的为豆形胡桃蛤(Nuculakawamurai)和豆形短眼蟹(Xenophthalmuspinnotheroides)。

图5 胶州湾大型底栖动物年次级生产力(不计养殖种)平面分布

表7 养殖区各站位养殖贝类年次级生产力
3.3 胶州湾大型底栖动物群落年次级生产力与历史资料的对比
本研究结果与历史资料的对比见表8。由表8可知,本研究的大型底栖动物次级生产力要显著高于渤海的三大湾以及乳山湾、桑沟湾和海州湾的值,原因是胶州湾具有高的初级生产力水平(见表9)。本研究的大型底栖动物次级生产力也高于表8中所列胶州湾其它报道的值,原因很可能是胶州湾贝类养殖量的逐年增加所致[41-43]。

表8 本研究与历史资料对比

表9 胶州湾及其邻近海湾初级生产力水平
其实,不同的研究之间,因调查时间和站位设置等的不同,所得到的年次级生产力的可比性不大。因为一个站位大型底栖动物群落次级生产力值的大小,取决于该站位出现的各种的大型底栖动物的平均个体数和平均生物量,一年中调查取样次数越多(即取样时间间隔越短),这两个值就越准确,估算的群落的次级生产力就越接近真值。一个海域的大型底栖动物群落的年次级生产力,是各站位次级生产力的平均,站位数越多,其平均值越接近海域的大型底栖动物群落年次级生产力的真值。因此,站位数不同和一年中调查次数的不同,是不同研究结果之间出现差别的重要原因。
3.4 胶州湾大型底栖动物高次级生产力水平与养殖容量
由表9可知,胶州湾初级生产力水平(以碳计)明显高于相邻海湾,仅莱州湾初级生产力水平与胶州湾接近。董世鹏等[43]基于初级生产力水平估算的胶州湾菲律宾蛤仔(R.philippinarum)的养殖容量为429.00 ind./m2,本研究养殖种的平均丰度已达546.00 ind./m2,超过了上述的养殖容量。
3.5 大型底栖动物次级生产力估算模型的选择
目前估算大型底栖动物次级生产力使用最多的为Brey(1990)模型,该模型未区分不同的大型底栖动物类群,也未考虑环境因素的影响。Brey(2001)模型区分了大型底栖动物不同类群,并增加了环境因素,包括底温、水深、群落的栖息地类型等。Brey(2012)模型中又增加了大型底栖动物食性和移动方式等参数,模型日趋完善。因为大型底栖动物类群众多,很多种的食性及移动方式尚不清楚,使用Brey(2012)模型有一定的难度,所以本研究采用了Brey(2001)模型,来估算胶州湾大型底栖动物群落的年次级生产力。
4 结论
(1)胶州湾大型底栖动物群落次级生产力的分布,呈现出从湾中部向南北两侧逐渐增大的趋势。
(2)底盐和叶绿素a是影响胶州湾大型底栖动物次级生产力的重要环境因子。
