长足大竹象外切葡聚糖酶活性最适反应条件的初步探究
2021-07-19唐昊王昌吉王明珺甘晓凤张鑫月罗朝兵
唐昊,王昌吉,王明珺,甘晓凤,张鑫月,罗朝兵
(乐山师范学院生命科学学院,四川 乐山 614000)
竹纤维是储量巨大的可再生资源,利用生物法处理木质纤维素并使之能够进行能源转化是当前国际研究热点之一[1-2].自然界中除微生物降解纤维素外,还存在着超过100种昆虫依靠自身肠道系统对木质纤维素实现高效降解,其中纤维素酶系统在其中发挥着重要作用[3-4].
昆虫是种类繁多的生物种群,研究发现具有天然生物质利用系统(natural biomass utilization systems,NBUS)的植食性昆虫是开发工业纤维素分解酶的潜在资源[5].外切葡聚糖酶是植食性昆虫消化道纤维素酶系中重要组分,降解过程中使得纤维素分子结构变得疏松而有利于其它纤维素酶进入纤维素内部,从而加速纤维素降解[6],然而外切葡聚糖酶对纤维二糖的耐受性差且酶活较低,因而迫使需要挖掘新的酶资源.宁娜等[7]对台湾乳白蚁和黄翅大白蚁消化道中纤维素降解外切葡聚糖酶活性比较后发现,黄翅大白蚁外切葡聚糖酶集中于中肠,而台湾乳白蚁主要集中于后肠;Wang等[8]发现福寿螺(Ampullariacrossean)外切葡聚糖酶对高浓度纤维二糖具有耐受性;佟新新[9]研究纤维素酶解蔗渣工艺中发现黑曲霉外切葡聚糖酶具有较好的热稳定性和pH稳定性;赵宣等[10]克隆了草酸青霉外切葡聚糖酶基因并在毕赤酵母中成功表达.这些研究结果表明,不同的昆虫的肠道分段中,外切葡聚糖酶活性具有较大的差异,同时具有潜在的利用价值并可应用于纤维素的利用.
长足大竹象(Cyrtotrachelusbuqueti)是慈竹、青皮竹和大叶慈等丛生竹类植物的重要害虫,危害竹材生长,严重阻碍了竹产业的发展[11].已有研究表明,长足大竹象消化道内具有较高的纤维素酶活,同时其发育转录组表明其幼虫和成虫阶段,存在大量的碳水化合物活性酶(CAZy)基因表达[12].目前,长足大竹象消化道纤维素酶中内切葡聚糖酶酶活性质已探明[13],但外切葡聚糖酶反应条件尚不清楚.因此,本研究设计单因素试验和正交优化试验探究长足大竹象外切葡聚糖酶的最适反应条件,为长足大竹象外切葡聚糖酶的挖掘提供借鉴并为竹木质纤维素的能源转化和生产提供新的酶资源.
1 材料与方法
1.1 材料
1.1.1 昆虫样本的收集及粗酶液的制备 样本长足大竹象采集于四川省沐川县(E 103°90′,N 28°97′).样本处理流程如下:分别取40只雌虫、雄虫及幼虫饥饿24 h后,在无菌室超净工作台里解剖出消化道放置于无菌水及生理盐水中清洗4~5次,低温无菌研钵器皿中研磨至匀浆,离心10 min后收集上清液(4 ℃、4球000 r/min),经0.22 μm的滤膜处理后作为粗酶液.所有试验生物学重复5次.
1.1.2 试剂与仪器 微晶纤维素(MCC),3,5-二硝基水杨酸(DNS),葡萄糖(D-(+)-glucose),标准蛋白质(BSA),柠檬酸缓冲液(pH 5.6),考马斯亮蓝(Coomassie Brilliant Blue-250);酶标仪,高速冷冻离心机,恒温混匀仪,pH仪.
1.2 试验方法
1.2.1 标准曲线的确定 采用Bradford法确定蛋白质标准曲线[14].取7只试管分别加入0.00,0.01,0.02,0.03,0.04,0.05,0.06 mL 1 mg/mL的标准蛋白质溶液,柠檬酸缓冲液(pH 5.6、0.1 mol/L)定容到0.1 mL,加入3 mL的Brilliant Blue G-250,595.0 nm处所测定吸光值作为纵坐标,蛋白质的浓度标准作为横向的坐标,绘制蛋白质标准曲线.
DNS法确定葡萄糖标准曲线[15].取8支试管分别加入0.00,0.01,0.02,0.03,0.04,0.05,0.06,0.07 mL的葡萄糖标准溶液(1 mg/mL)和0.02 g微晶纤维素底物,蒸馏水定容至1 mL,加入1.5 mL DNS显色剂加热5 min后冷却至室温,540 nm波长处测得各个浓度的吸光值作为纵坐标,葡萄糖含量为横坐标,绘制葡萄糖标准曲线.
1.2.2 长足大竹象外切葡聚糖酶活性测定方法 外切葡聚糖酶活性采用3,5-二硝基水杨酸(DNS法)显色法测定[15].以微晶纤维素作为反应底物,0.1 mmol/L的柠檬酸缓冲液为溶剂,分别取50 μL酶液和950 μL底物充分混匀,恒温37 ℃水浴反应15 min后加入1.5 mL的DNS终止反应,沸水加热5 min后定容至20 mL,使用酶标仪测定吸光值(λ=540 nm).对照组酶液进行灭活处理,步骤与上述步骤相同,每组试验进行5次重复.
外切葡聚糖酶活力单位的定义:底物在相应温度下反应每分钟酶解产生1 μmol葡萄糖的酶量.酶活力及酶比活力的计算公式如下:
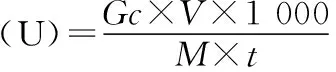
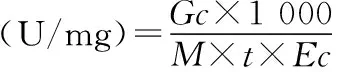
式中:Gc:葡萄糖标准曲线中葡萄糖相应的浓度,mg/mL;V:反应体系总体积,为20 mL;M:葡萄糖相对分子质量,180;t:反应时间;Ec:蛋白质标准曲线中蛋白质相应的浓度:mg/mL.
1.2.3 单因素试验设计 选取pH、底物浓度和反应温度作为单因素条件探究每种因素的最适反应条件(表1),确定长足大竹象外切葡聚糖酶的最佳理论反应条件.
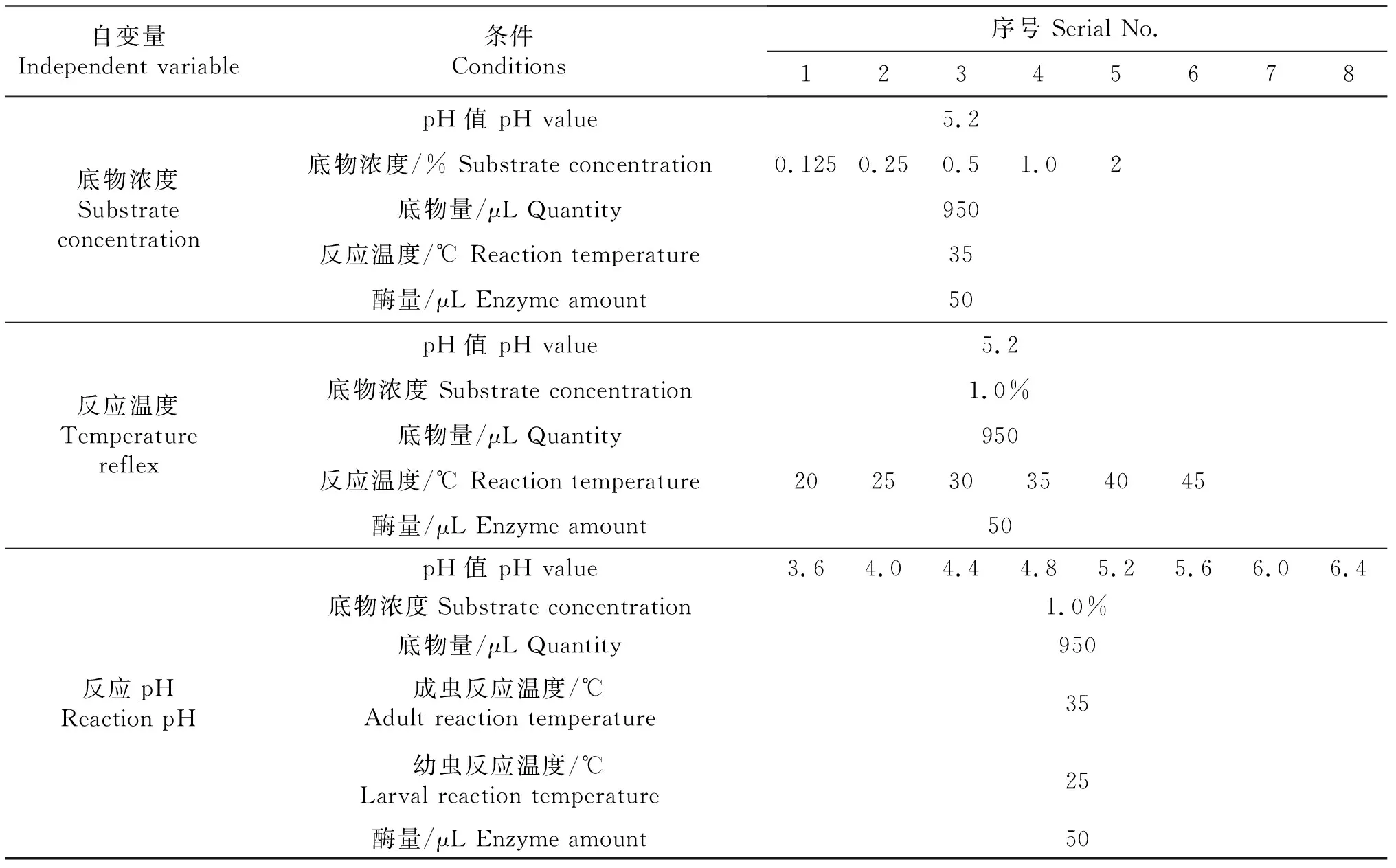
表1 单因素试验设计
1.2.4 正交试验设计 为进一步优化外切葡聚糖酶pH、底物浓度和反应温度最适反应条件,本试验设计了正交试验(表2).

表2 正交试验设计

2 结果与分析
2.1 标准曲线
通过Bradford法确定了蛋白质的标准曲线,曲线方程为y=3.785x-0.002 8(R2=0.996 8).同时,由DNS法确定了葡萄糖标准曲线,曲线方程为y=0.977 9x-0.025 2(R2=0.992 4).
2.2 单因素试验确定长足大竹象外切葡聚糖酶的最适反应条件
2.2.1 长足大竹象外切葡聚糖酶的最适反应温度 根据表1的试验设计,以温度为横坐标,长足大竹象外切纤维素酶比活力为纵坐标比较了不同反应温度下长足大竹象雌虫、雄虫及幼虫外切葡聚糖酶的活性.由图1可知,长足大竹象外切葡聚糖酶比活力随着温度的上升均呈现出先上升后下降的趋势,雌虫在30 ℃出现最大值,雄虫在30~40 ℃之间出现最大值,而对于幼虫而言,其在40 ℃出现最大值,由此可推断长足大竹象外切葡聚糖酶活性最适反应温度介于30~40 ℃之间.
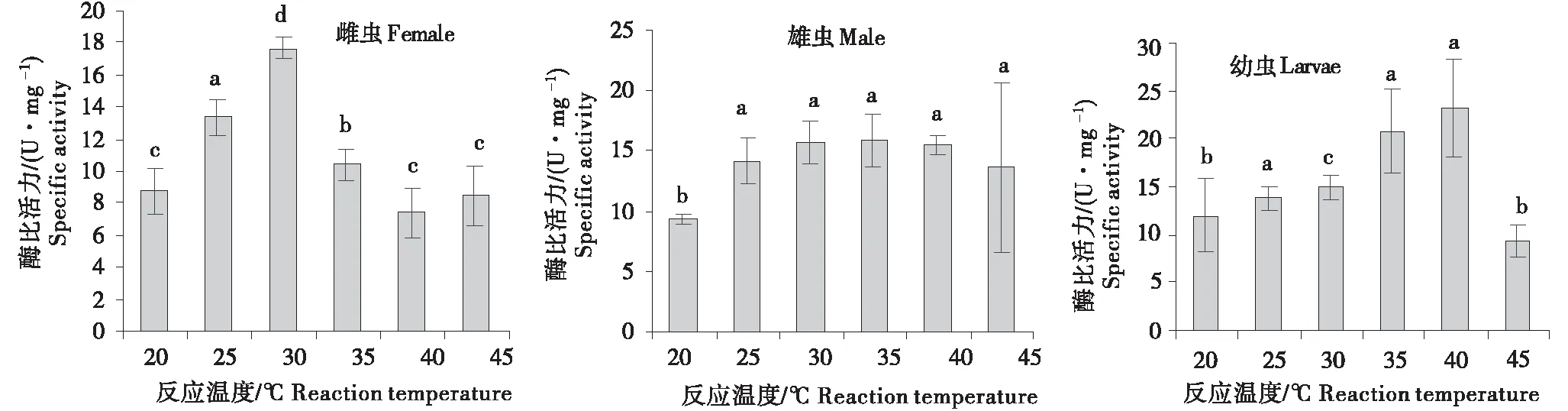
小写字母的不同表示差异显著(P<0.05).Different letters represent the significant differences(P<0.05).
2.2.2 长足大竹象外切葡聚糖酶的最适底物浓度 对于长足大竹象成虫,外切葡聚糖酶比活力随着底物浓度的升高逐渐上升,当底物浓度为2%时,酶比活力达到最大值;而幼虫在底物浓度为0.5%时比活力达到最大值,随着底物浓度的升高,外切葡聚糖比活力有下降的趋势(图2).以上结果表明长足大竹象成虫的外切葡聚糖的表现出最大酶活力时,最适底物浓度为2%,而幼虫的为0.5%.
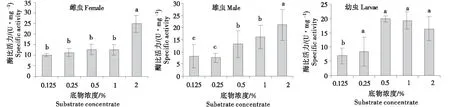
*小写字母的不同表示差异显著(P<0.05).Different letters represent the significant differences(P<0.05).
2.2.3 长足大竹象外切葡聚糖酶的最适pH条件 由图3可知,长足大竹象外切葡聚糖酶活性随着酶反应pH的上升,呈现出先上升后下降的趋势.其中长足大竹象成虫(雌)的外切葡聚糖酶的最大比活力出现在pH 4.8处,成虫(雄)的外切葡聚糖酶的最大比活力出现在pH 4.8~5.2的范围内且在pH 4.8和pH 5.2之间酶比活力未表现出显著的差异;而幼虫的酶比活力在pH 4.4~5.6出现最大值,在该pH区间内,酶比活力也未表现出显著的差异.以上结果说明成虫和幼虫外切葡聚糖酶的最适反应pH分别介于区间4.8~5.2和4.4~5.6之间.
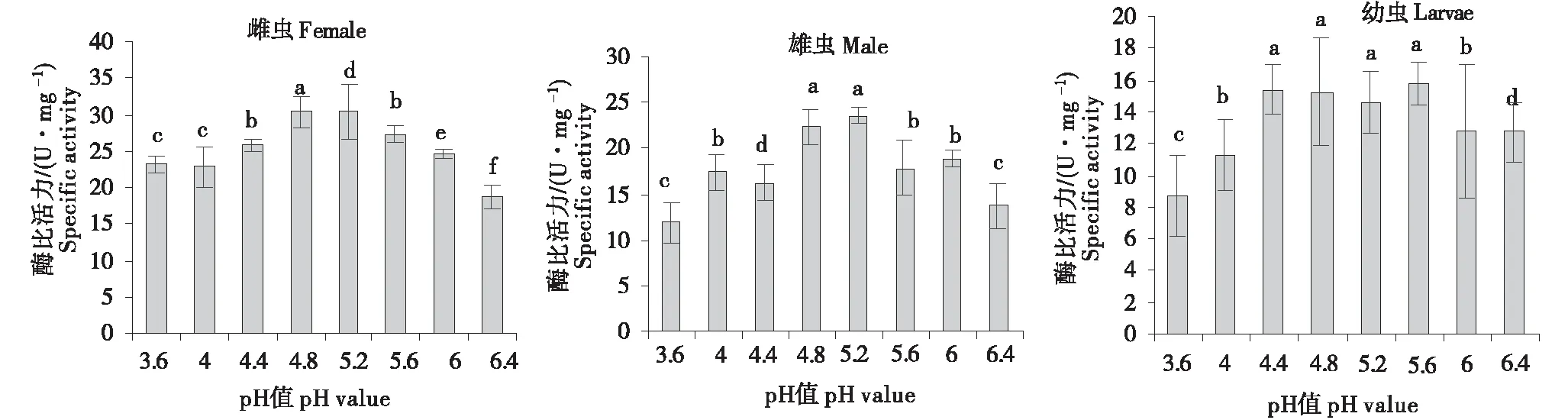
*小写字母的不同表示差异显著(P<0.05).
2.2.4 长足大竹象外切葡聚糖酶单因素试验结果 单因素试验分别探讨了外切葡聚糖酶表现出最大酶比活力的的单个最适反应条件,结果表明,长足大竹象外切葡聚糖酶最大酶活力的单个最适反应条件为:雌虫在30 ℃出现最大值,底物浓度为2%,pH为 4.8;雄虫在30~40 ℃之间出现最大值,底物浓度为2%,pH为 4.8~5.2;而对于幼虫而言,反应温度为40 ℃,底物浓度为0.5%,pH为 4.4~5.6.
为了进一步验证3个单因素对外切葡聚糖酶的影响,进行了ANOVA分析,结果表明底物浓度与pH对长足大竹象外切葡糖酶活性具有显著影响,而反应温度仅对雌虫酶活性表现出显著影响(表3).
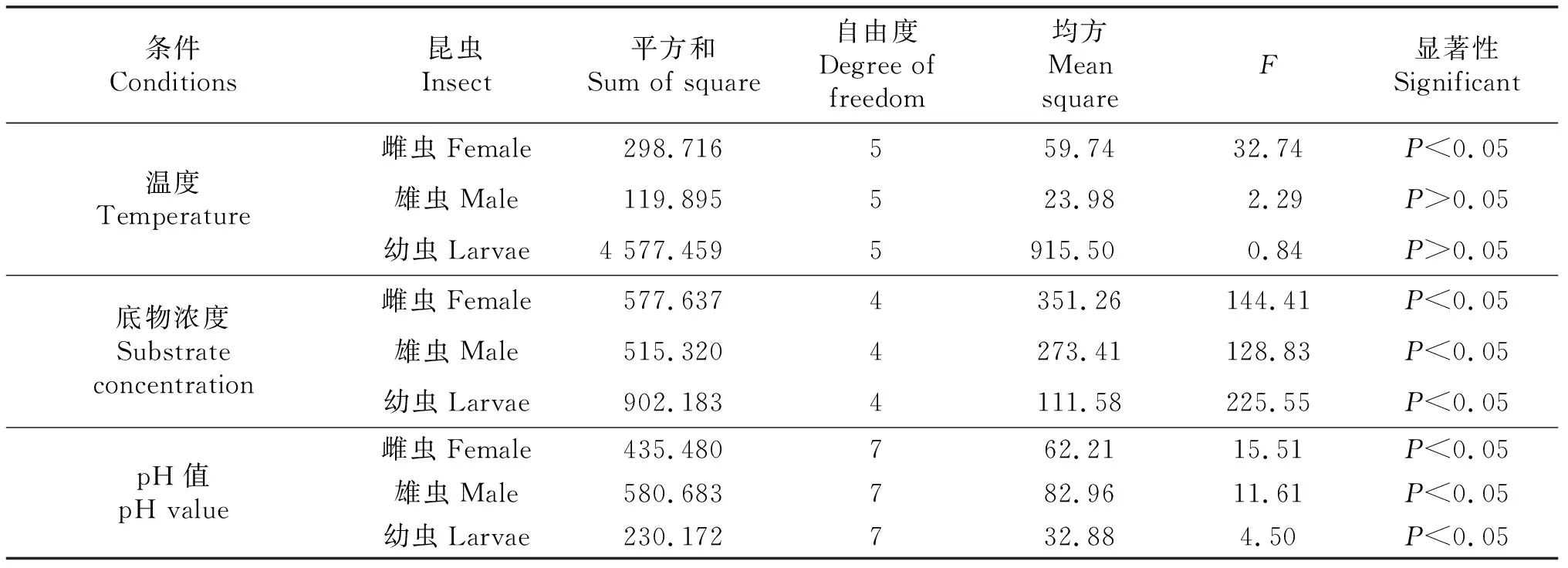
表3 单因素试验结果的ANOVA分析
2.3 正交试验
单因素试验得到了外切葡聚糖酶在单个因素影响下的酶最适反应条件,通过设计正交优化试验.进一步验证和优化长足大竹象外切葡聚糖酶在多因素影响下的最适反应条件(表2).对正交试验结果进行极差分析,结果表明反应温度对长足大竹象外切葡聚糖酶活性影响最大,其次是pH,而底物浓度对酶活性的影响最小.此外,在3种因素交互作用中pH和底物浓度对成虫的酶活性无显著性影响(P>0.05),而温度仅对幼虫的酶活性有显著性影响(P<0.05)(表4~5).
表4 酶活性正交试验结果
2.4 单因素试验与正交试验结果比较分析
在正交试验多因素交互作用分析结果中pH和底物浓度对成虫的酶活性影响并不显著,所以对于成虫酶活性的最适反应条件的确定主要以单因素试验的结果为主;而幼虫酶活性的最适反应条件优先考虑温度,其次为底物浓度,最后为pH值.综合单因素度验和正交试验的结果,初步推断长足大竹象幼虫的酶活性最适反应条件为温度40 ℃,pH 4.8,0.5%底物浓度;成虫(雌)温度为30 ℃,pH为5.2,底物反应浓度为2%;成虫(雄)温度为30 ℃,pH为4.8,底物反应浓度2%.为了验证单因素最佳组合及正交最佳组合反应条件下外切葡聚糖的酶活性.我们重新测定了相应条件下外切葡聚糖的酶活性.结果如图4所示,长足大竹象雄成虫和幼虫的外切葡聚糖酶活性正交优化结果优于单因素最佳组合酶活(P<0.01),而雌成虫的长足大竹象外切葡聚糖酶活性在组间无显著差异(P>0.05).同时在单因素最佳组合或正交最佳组合反应条件下,外切葡聚糖酶活性在虫态间呈现出雌虫>雄虫>幼虫的趋势,表明在该酶不同虫态间存在差异.
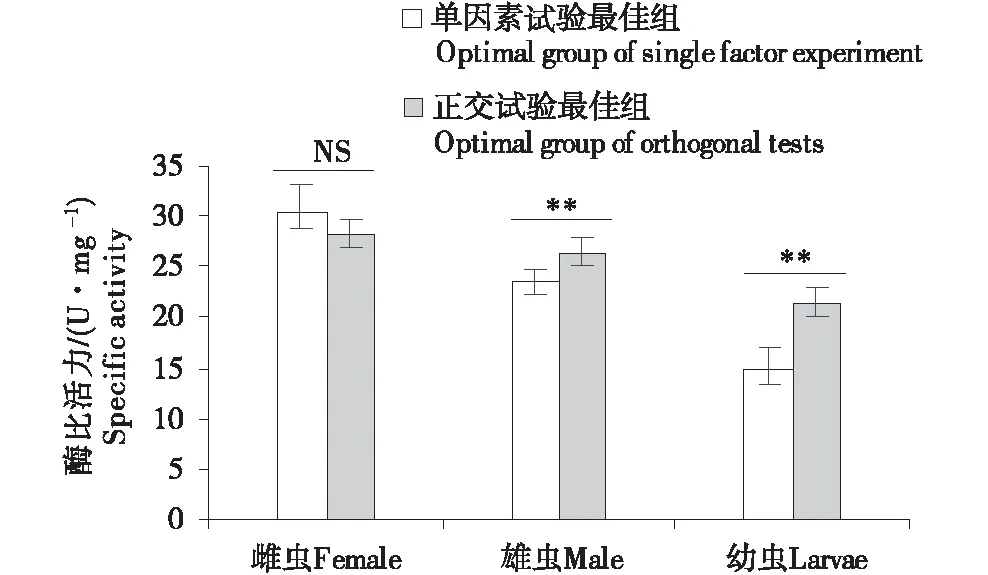
NS:差异不显著;**:差异显著(P<0.01).
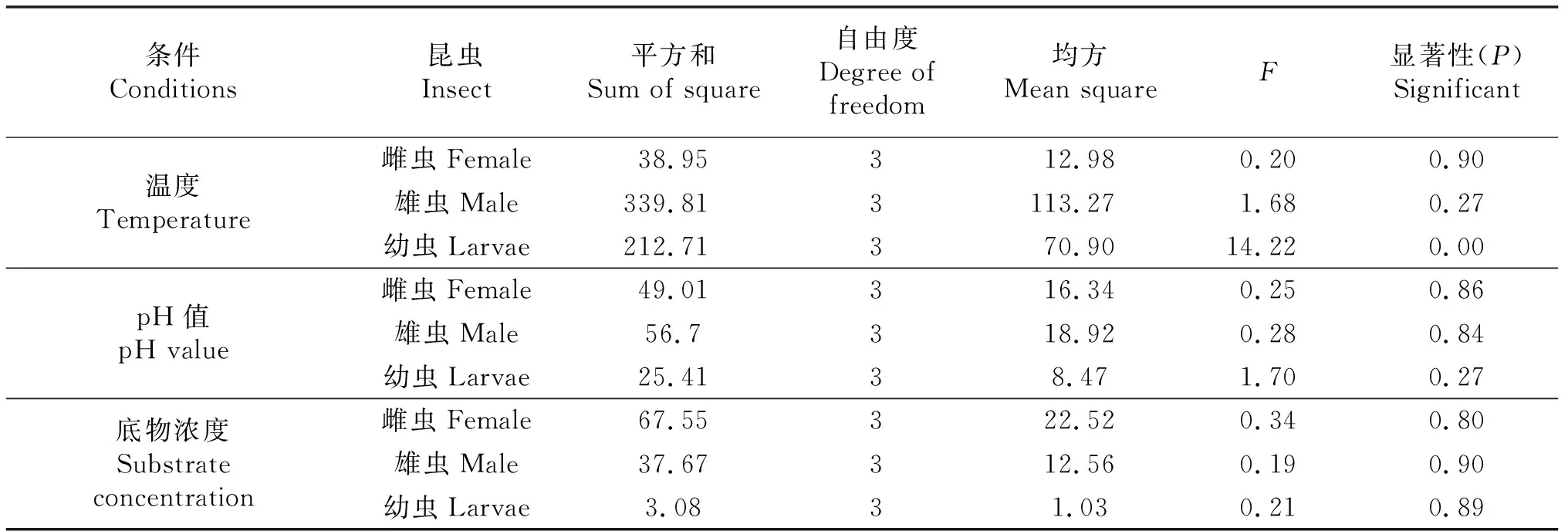
表5 正交试验结果方差分析
3 讨论
我国的拥有丰富的竹林资源,但竹资源中竹纤维素的能源化利用率较低,从节能环保及生态的角度而言,通过生物降解的方式来逐步利用竹木质纤维素中含有的纤维素可能将成为一种趋势,因此迫切的需要挖掘新的生物酶资源,其中长足大竹象拥有一套优秀的天然生物质利用系统[11-12,16],其消化道中的纤维素酶在生物降解利用方面具有重要的研究价值.
本研究以纤维素酶类中的外切葡聚糖酶为研究对象,通过单因素试验和正交试验设计,得出长足大竹象在最适反应条件下外切葡聚糖酶比活力,相较于取食木材木质纤维素的桑肩天牛、光肩星天牛和松褐天牛(酶活分别为0.091、0.123、0.141 U/mL)[17],长足大竹象外切葡聚糖酶比活力更高.此外,就酶的比活力而言,长足大竹象的内切葡聚糖酶[13]高于外切葡聚糖酶.在本实验中,长足大竹象外切葡聚糖酶的最适pH位于4.8~5.2之间,与Li等[18]研究来源于Caldicellulosiruptor外切葡聚糖酶反应条件pH 4.7~5.5较为接近.长足大竹象外切葡聚糖酶最适反应温度30 ℃~40 ℃,与家蝇幼虫外切葡聚糖酶最适温度50 ℃相比[19],长足大竹象外切葡聚糖酶的最适温度较温和.本研究还发现长足大竹象成虫外切葡聚糖酶活性大于幼虫,存在虫态间显著差异,可能原因是取食习惯的差异所致[20].
外切葡聚糖酶主要参与纤维素的降解,高树广等[21]发现芝麻茎点枯病菌(Macrophominaphaseolina)产的纤维素酶对芝麻秸秆具有降解作用,同时于慧娟和郭夏丽[22]在筛选高效秸秆降解菌的同时发现外切葡聚糖酶参与了纤维素的降解过程.尽管目前在多种植食性昆虫和农田废弃物中筛选和发现了多种纤维素降解细菌和真菌,但如何提升这些微生物分泌的纤维素酶的耐受力和提升纤维素酶的降解效率仍是难题.李建树等从牛粪自然堆肥样品中筛选出较耐高温的纤维素降解细菌和真菌各一株[23].Xue等[24]从海洋黑曲霉中提取出外切葡聚糖酶并发现其具备耐盐性,这些研究对纤维素降解菌或纤维素酶的耐受性方面的后续研究提供了参考价值.此外,唐自钟等[25]利用分子生物学技术手段实现外切葡聚糖酶基因在毕赤酵母中成功表达,并在60 ℃下保温一个小时仍能保持较高活性;孔芹等[26]采用基因重组技术提高了外切葡聚糖酶活性并发现可以提升纤维素酶总活力;此外,通过将内切葡聚糖酶和外切葡聚糖酶2种酶中存在的同源碳水化合物(CBM-2)融合得到融合蛋白CxnA和CxnA,在大肠杆菌和弗氏柠檬酸杆菌中表达有效改善了纤维素降解能力[27-28].
4 结论
通过单因素试验和正交试验的结果,初步推断长足大竹象幼虫的酶活性最适反应条件为温度40 ℃,pH 4.8,0.5%底物浓度;成虫(雌)温度为30 ℃,pH为5.2,底物反应浓度为2%;成虫(雄)温度为30 ℃,pH为4.8,底物反应浓度2%,在对应的最适反应条件下,长足大竹象外切葡聚糖酶比活力分别为雌成虫28.26 U/mg、雄成虫26.42 U/mg、幼虫21.50 U/mg,同时也表明该酶在不同虫态间存在差异.综上,本研究对长足大竹象外切葡聚糖酶最适反应条件的研究,可以更加深入的了解长足大竹象消化道的纤维素降解酶系统,为挖掘外切葡聚糖酶基因和后期进一步解决竹材木质纤维素的生物降解过程的某些技术难题提供部分参考信息.
