仔猪年龄对血清酶及代谢产物、免疫球蛋白与粪便微生物影响的研究
2021-07-17丁庆华赵航彬姜淑贞李会荣
丁庆华,刘 婕,赵航彬,姜淑贞,李会荣*
(1.山东省畜产品质量安全中心,山东济南 250100:2.山东农业大学动物科技学院,山东泰安 271018)
杜× 长× 大三元杂交商品猪的生长速度明显快于纯种猪,具有明显的杂交优势[1]。生长期内猪的各个部分大致按照神经组织、骨骼、肌肉和脂肪的生长顺序进行生长发育[2]。动物的年龄能够影响动物的健康状况、繁殖力及生产性能,动物机体许多生理指标都随年龄增长而发生相应变化[3]。年龄对仔猪健康的影响主要受消化器官发育完善程度的制约,但补饲时间和营养水平对仔猪健康也有重要影响。通过各种生理指标的测定与分析,能够更好地掌握仔猪不同生长发育阶段的规律,有利于指导仔猪生产。近年来,大量文献表明,日粮养分和添加剂对各生长阶段猪抗氧化能力、血清酶和代谢产物、免疫指标和微生物菌群有显著影响[4-7]。本研究旨在探讨仔猪年龄对抗氧化能力、血清酶和代谢产物、免疫指标和微生物菌群的影响规律,以期为仔猪生产提供参考。
1 材料与方法
1.1 试验动物与设计 选择三胎、体重(14.96±0.30)kg 的38~42 日龄健康断奶杜× 长× 大仔猪112 头,随机分为8 个重复,每个重复14 头,各重复仔猪初始体重差异不显著。试验饲粮按照ΝRC(1998)标准配制,原料组成和营养成分见表1。
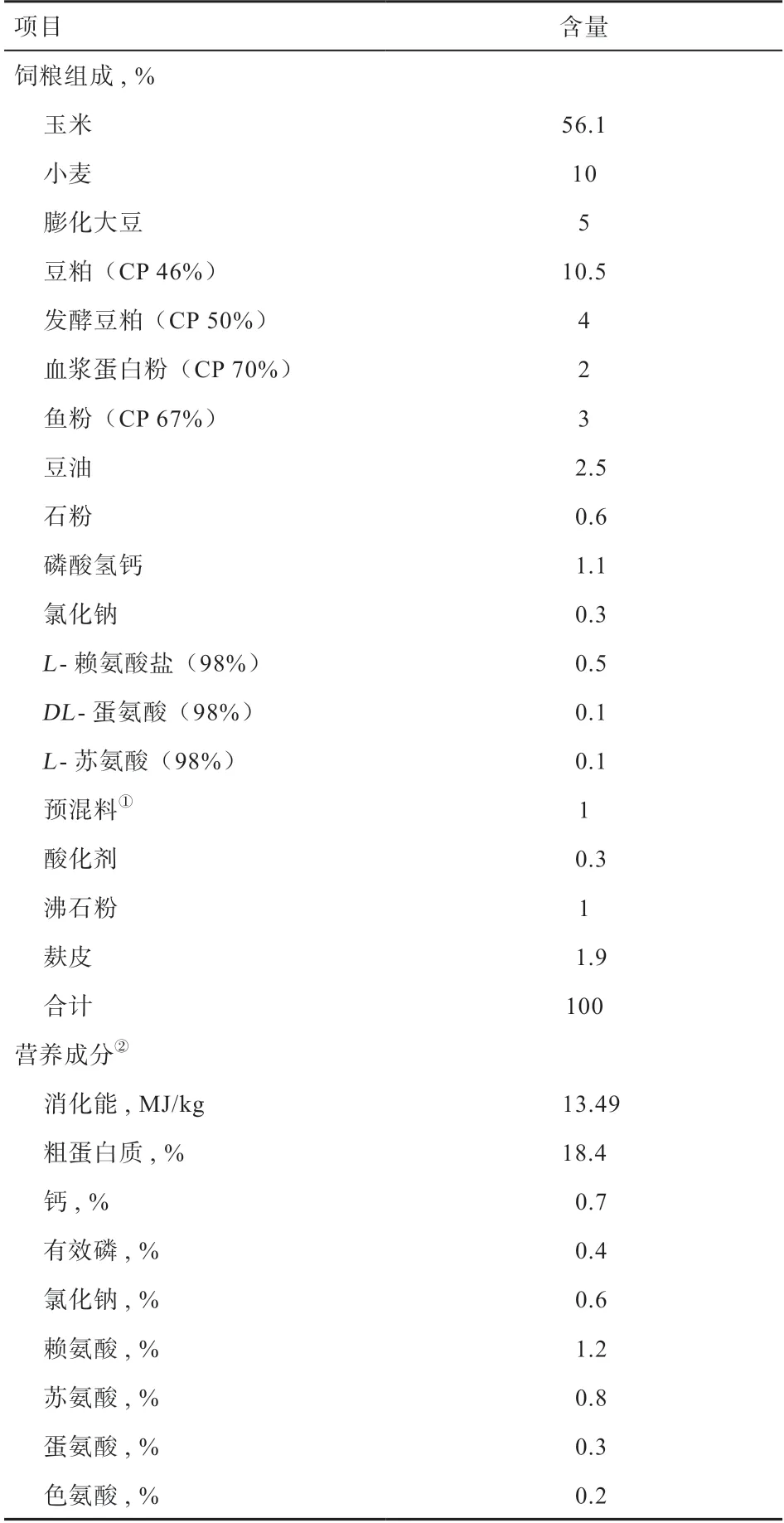
表1 试验饲粮组成和营养成分(风干基础)
1.2 饲养管理 试验仔猪饲养在同一猪舍内,每个重复的仔猪饲养在同一栏舍,试验开始前对仔猪舍进行彻底刷洗、消毒。试验过程中每天清扫猪舍,保证舍内清洁,自然通风,按时消毒,记录每天猪舍的温度和湿度,观察不同时间段猪的行为状态、毛色、采食和活动状况。试验期间仔猪自由采食饮水。预试7 d,正试期42 d。
1.3 生产性能 试验期间每周记录一次仔猪的进食量。试验开始、第21 天和42 天对仔猪进行称重,计算平均日增重(ADG)、平均日采食量(ADFΙ)和耗料增重比(F/G)。
1.4 样品采集与指标测定
1.4.1 血清样品采集 分别于试验第0、21、42 天前腔静脉空腹采血。用5 mL 促凝管4 000 r/min 离心10 min,分离血清,置于-20℃冰箱保存,用于测定血清抗氧化物酶、血清酶、血清代谢产物和免疫球蛋白。
1.4.2 血清抗氧化物酶的测定 总超氧化物歧化酶(T-SOD)活性采用黄嘌呤氧化酶法(羟胺法)测定,谷胱甘肽过氧化物酶(GSH-Px)活力和丙二醛(MDA)含量采用比色法测定。按照抗氧化测试盒(A001-1、A005 和A003-1,南京建成生物工程研究所)的步骤检测。
1.4.3 血清酶和血清代谢产物的测定 采用COBUS MΙRA Plus 全自动生化分析仪(Roche Diagnostic System Ιnc.,USA)测定血清生化指标。血清酶包括谷丙转氨酶(AST)、谷草转氨酶(ALT)、碱性磷酸酶(ALP)、乳酸脱氢酶(LDH);血清代谢产物包括总蛋白(TP)、尿素氮(SUΝ)、总胆固醇(TC)、甘油三酯(TG)、高密度脂蛋白(HDL)和低密度脂蛋白(LDL)。测定指标所用试剂盒购自南京建成生物工程研究所。
1.4.4 血清免疫球蛋白的测定 采用免疫比浊法,使用免疫球蛋白测试盒(E027、E026、E025,南京建成生物工程研究所)测定血清免疫球蛋白A(ΙgA)、ΙgG、ΙgM 的浓度。
1.4.5 粪便微生物测定 分别于试验第0、21、42 天晨饲前直肠取粪样,放入已灭菌的4 mL 离心管中,并将装有样品的离心管放入装有冰袋的泡沫箱中保存,用于测定微生物群落。采用细菌培养基培养法测定大肠杆菌和乳酸菌的数量,细菌数量采用平板菌落计数法进行统计。计算出每克粪便中所含菌数(用对数lgCFU/g 表示)。
1.5 统计分析 数据采用SAS 9.2 软件进行统计处理。采用单因素方差分析进行分析统计,各平均值之间用Duncan's 进行多重比较。采用正交多项式比较法对不同年龄的处理效应进行一次线性回归分析,试验数据用平均值和SEM 表示,P<0.05 表示差异显著。
2 结果与分析
2.1 生长性能随仔猪年龄增长的变化规律 由表2 可见,试验第0~21 天仔猪ADFΙ、ADG 和F/G 均显著低于试验第0~42 天和21~42 天,试验第0~42 天仔猪ADFΙ、ADG 和F/G 又显著低于试验第21~42 天。
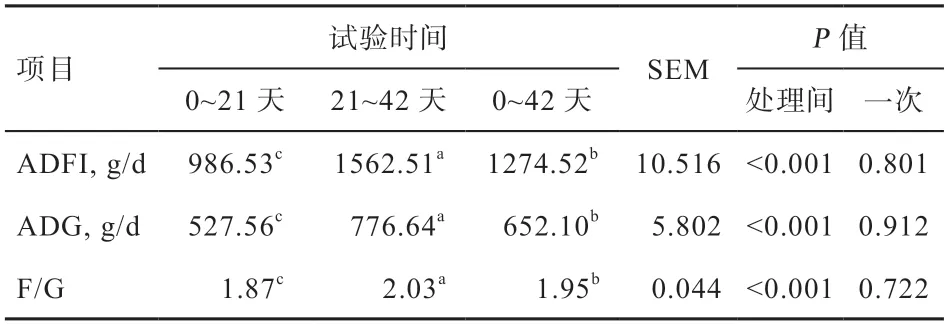
表2 生长性能随仔猪年龄增长的变化规律
2.2 血清抗氧化能力随仔猪年龄增长的变化规律 由表3 可见,血清T-SOD 和GSH-Px 活性随仔猪年龄增长呈线性升高(P<0.05),而MDA 含量随仔猪年龄增长呈线性降低(P<0.05)。方差分析表明,试验第21 天仔猪血清T-SOD 和GSH-Px 高于试验第0 天(P<0.05),试验第42 天血清T-SOD 和GSH-Px 高于试验第21 d(P<0.05)。试验第0 天和21 天血清MDA 含量高于试验第42 d(P<0.05),试验第0 天和第21 天血清MDA 含量差异不显著。

表3 血清抗氧化酶活性和MDA 含量随仔猪年龄增长的变化规律
2.3 血清酶随仔猪年龄增长的变化规律 血清酶活性随仔猪年龄的变化规律见表4。血清ALT 和ALP 活性随仔猪年龄增长呈线性降低(P<0.05),而LDH 活性随仔猪年龄增长呈线性升高(P<0.05)。方差分析表明,试验第21 天和第42 天血清ALT 活性均低于试验第0天(P<0.05),试验第42 天血清ALP 活性低于试验第0 天和第21 天(P<0.05),其他均差异不显著。

表4 血清酶随仔猪年龄增长的变化规律 U/L
2.4 血清代谢产物随仔猪年龄增长的变化规律 血清代谢产物随仔猪年龄增长的变化规律见表5。血清SUΝ、TC 和HDL 含量随仔猪年龄增长呈线性升高(P<0.05),而血清TP 含量则线性降低(P<0.05)。方差分析表明,试验第21 天和第42 天血清TP 含量低于试验第0 天(P<0.05)。试验第42 天血清SUΝ 和TC 高于试验第0 天和第21 天(P<0.05)。试验第42 天血清HDL 高于试验第0 天(P<0.05),其他均差异不显著。
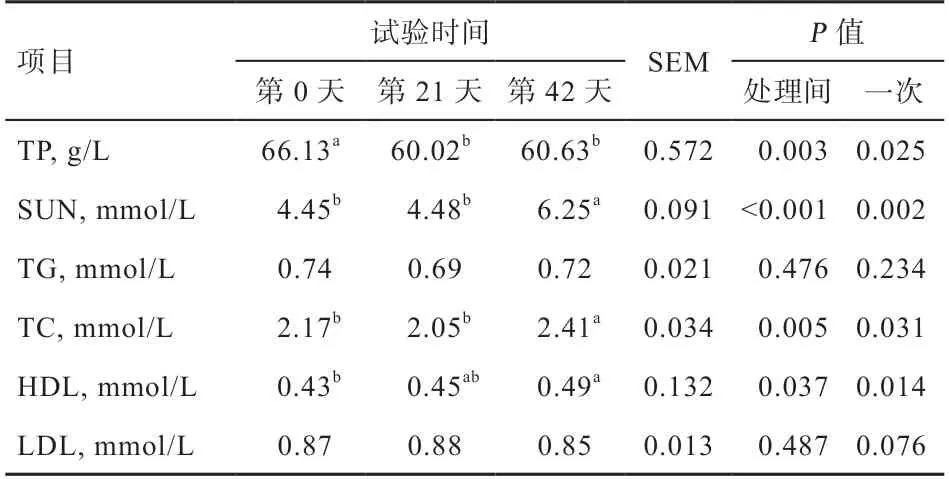
表5 血清代谢产物随仔猪年龄增长的变化规律
2.5 血清免疫球蛋白随仔猪年龄增长的变化规律 血清免疫球蛋白水平随仔猪年龄的变化规律见表6。血清ΙgA、ΙgG 和ΙgM 水平均随仔猪年龄增长呈线性降低(P<0.05)。方差分析表明,试验第21 天和第42 天血清ΙgA、ΙgG 和ΙgM 均低于试验第0 天(P<0.05),试验第0 天和第21 天血清ΙgA、ΙgG 和ΙgM 差异不显著。

表6 血清免疫球蛋白随仔猪年龄增长的变化规律 g/L
2.6 粪便微生物随仔猪年龄增长的变化规律 粪便微生物随仔猪年龄的变化规律见表7。粪便大肠杆菌含量随仔猪年龄增长呈线性降低(P<0.05)。方差分析表明,试验第21 天和42 天粪便大肠杆菌低于试验第0 天(P<0.05),其他均差异不显著。
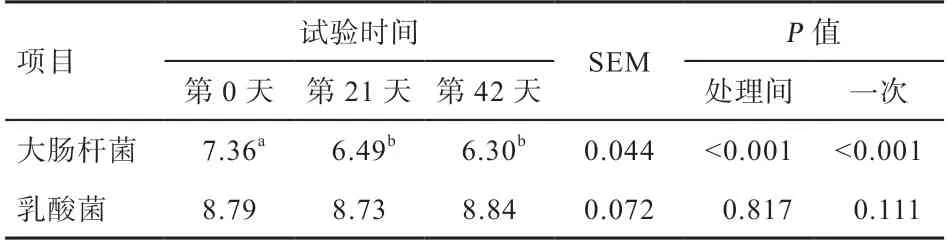
表7 粪便微生物随仔猪年龄增长的变化规律 lgCFU/g
3 讨论
3.1 仔猪年龄对血清抗氧化酶和MDA 的影响 血清中T-SOD 和GSH-Px 含量可以反映机体对抗自由基损伤能力的程度[8],T-SOD 和GSH-Px 含量越高,表明机体抗氧化能力越强。反之,则抗氧化能力越弱。MDA 是自由基攻击细胞膜上多不饱和脂肪酸而产生的过氧化脂质的代谢产物,血清中MDA 含量可以间接反映自由基对细胞的损伤水平及脂质过氧化程度。MDA 含量越高,表明自由基对细胞的损伤程度越大。反之,则损伤程度越小[9]。本研究结果表明,断奶仔猪血清T-SOD 和GSH-Px 随仔猪年龄增长呈线性升高,血清MDA 含量随仔猪年龄增长呈线性降低,表明机体的免疫力和抵抗力随着年龄增长而提高,自体抵抗氧化应激能力增强,能够及时清除增多的自由基,从而使脂质过氧化反应减少。但其影响的机制尚需进一步证实。
3.2 仔猪年龄对血清酶的影响 正常代谢条件下,ALT和AST 主要存在于肝脏和心脏细胞中,血清中的活性较低[10]。肝损伤或应激时,ALT 和AST 释放入血导致酶活升高,血清酶活性是衡量肝脏健康和应激反应程度的重要指标[11]。在骨代谢旺盛时,成骨细胞分泌的ALP 增多,ALP 大量存在于成骨细胞的周围及其表面,极易释放进入血液,因此血清ALP 活性上升[12]。本试验结果表明,随断奶仔猪年龄增长其血清ALT 和ALP活性降低,LDH 活性升高,AST 活性无显著影响。可能是年龄越小骨代谢旺盛,但其机制尚需进一步证实。也有研究发现,ALP 能够清除肠道有害微生物产生的脂多糖等内毒素[13],对肠道有一定保护作用。而仔猪早期断奶通常会降低小肠ALP 的表达,不利于肠道的消化吸收。今后的研究可通过对仔猪小肠绒毛上皮刷状缘ALP的测定,进一步研究仔猪年龄对ALP 活性的影响。
3.3 仔猪年龄对血清代谢产物的影响 血清代谢产物是衡量机体蛋白质代谢水平的重要指标。研究表明,血清TP 含量增高,表明蛋白质在机体内代谢旺盛,是机体对氨基酸和蛋白质的吸收利用率提高的重要表现,同时也体现了蛋白质在肝脏中合成代谢提高以及组织蛋白质的沉淀作用增强[14]。本试验结果第42 天的血清TP 比第21 天略高,表明42 d 猪的胃肠发育促进了蛋白质代谢。血清SUΝ 含量能够反映动物机体蛋白质和氨基酸代谢的吸收和平衡情况[15]。氨基酸代谢平衡良好时,蛋白质吸收好,血清SUΝ 浓度下降。研究表明,在相同遗传背景下,14 日龄超早期断奶组仔猪在28 日龄的血清白蛋白、TP、白球比、TG、葡萄糖和SUΝ 含量均显著低于母乳组[16]。本试验结果表明,随仔猪年龄增长,氨基酸的吸收下降。HDL 和LDL 是TC 运输的主要载体,TC 被LDL 运至肝脏以外的组织,而被HDL 运回肝脏内参与其他物质的代谢转化,脂代谢平衡良好时,血清TC、HDL 和LDL 浓度下降或保持恒定[17]。本研究结果表明,随仔猪年龄增长脂质代谢增强。但是,关于仔猪年龄对血清代谢产物的影响机制尚需进一步研究。
3.4 仔猪年龄对血清免疫球蛋白的影响 血清免疫球蛋白含量是反映动物机体免疫功能的重要指标。本试验中,试验第0 天仔猪的血清ΙgA、ΙgG 和ΙgM 水平最高,因为新生仔猪能从初乳中获得免疫球蛋白,从而获得被动免疫;第42 天仔猪血清免疫球蛋白水平降低,因为仔猪此时尚未健全机体的免疫机能,血清免疫球蛋白含量在试验期内逐渐减少。研究表明,在相同遗传背景下,14 日龄超早期断奶组仔猪在28 日龄的血清ΙgA、ΙgG和ΙgM 均显著高于母乳组[16]。肠道是仔猪最大的消化器官和免疫器官,断奶仔猪由于断奶应激,导致细胞免疫能力受到抑制、采食量下降,肠道无法摄入充足的营养物质而发育水平较低,仔猪免疫系统一般要到4~7 周龄达到基本成熟[18]。今后可以通过判定小肠绒毛的发育状况,进行仔猪年龄对血清免疫球蛋白影响的机制研究。
3.5 仔猪年龄对粪便微生物的影响 粪便中大肠杆菌和乳酸菌含量是反映仔猪肠道健康状况的重要指标。本试验中,粪便大肠杆菌含量随仔猪年龄增长呈线性降低,乳酸菌含量无明显变化。在断奶阶段,许多应激因子刺激仔猪肠道使其形态结构和生长发育状况受到影响[13]。早期断奶仔猪胃壁内源性盐酸分泌受到抑制,致使胃肠道内pH 升高,高pH 环境下不适宜乳酸菌的生长繁殖,导致乳酸菌数量减少、大肠杆菌数量增加[19],但肠道黏膜损伤后能够快速修复,在断奶后5~10 d 就能够恢复至正常水平[20],这与本试验结果一致。研究表明,藏仔猪粪便微生物菌群结构在不同生长发育阶段存在差异,随着藏仔猪的生长,其肠道中可以降解纤维(厚壁菌门)和碳水化合物(拟杆菌门)等物质的菌群和具有抑制致病菌(乳杆菌科)作用的菌群相对丰度升高,而条件性致病菌(梭杆菌门)的相对丰度下降,说明其对营养物质的降解能力和肠道免疫能力提高[21]。本次试验经过了10 d 预试期,仔猪的胃肠道已经逐渐适应了该变化,机体消化道和消化腺逐渐成熟,胃肠道内的pH 逐渐恢复正常,良好的胃肠道条件为乳酸菌的生存提供了适宜环境,同时该环境也抑制了大肠杆菌的生长繁殖。今后也可通过探究仔猪胃肠道黏膜及微绒毛的发育状况,进一步研究仔猪年龄对粪便微生物的影响机制。
4 结论
本试验条件下,仔猪年龄显著影响血清酶活性、代谢产物和免疫球蛋白水平,改变粪便微生物数量,且随仔猪年龄增长,血清抗氧化能力增强,脂质代谢旺盛,粪便有害菌数量下降,但是蛋白质代谢水平下降,母体抗原水平降低。
