小麦干物质积累与分配规律研究进展
2021-07-16柯媛媛倪芊芊张乐乐魏凤珍李金才
柯媛媛,陈 翔,倪芊芊,张乐乐,魏凤珍,李金才,2*
(1.安徽农业大学农学院/农业部华东地区作物栽培科学观测站,安徽 合肥 230036;2.江苏省现代作物生产协同创新中心,江苏 南京 210095)
自1860年工业革命以来,全球气候显著变暖[1]。据最新的全球表面温度观测数据集[2-3]分析得出,1900—2017年间全球陆地平均气温升高(1.00±0.06)℃/100年。气候变化增加了粮食生产的不稳定性,而2050年全球人口预计将达到96亿,保障全球96亿人口的粮食安全问题将成为2050年面临的最大挑战。为满足日益增长的粮食需求,小麦生产力每年需增加1.6%[4]。解伟等通过构建Meta回归模型得出,全球平均气温每升高1℃,将导致我国主要粮食作物的单位面积产量降低2.6%[5]。小麦作为世界三大粮食作物之一,全球约60%的人口以小麦为主食[6]。在全球气候复杂多变和人口激增的背景下,维护小麦生产的稳定性对保障粮食安全具有极其重要的意义。
小麦干物质的积累与分配过程反映了小麦群体生长状况的好坏,也是决定小麦籽粒产量的重要因素。了解小麦在大田生长中干物质积累与分配的一般规律,以及基因型、主要栽培因子和生态因子对小麦干物质积累与分配的影响,对小麦生长发育状态的调控及建立高产群体结构具有重要意义。本文系统综述了小麦干物质积累与分配的一般规律,并阐述基因型、栽培因子和生态因子对小麦干物质积累与分配的影响,以期为改善小麦的种植制度,完善农田生态系统以及提高我国小麦生产能力提供参考。
1 小麦干物质积累与分配的一般规律
1.1 小麦干物质的积累规律
作物产量的高低与干物质积累过程密切相关,分析作物生长过程中干物质的积累动态对揭示作物产量形成和构建高效群体具有重要实践价值[7]。研究表明,随着小麦生长发育进程的推进,叶片和茎鞘的干物质积累量呈单峰曲线,而穗和植株的干物质积累量呈“S”型曲线变化[8]。小麦在不同生育时期,干物质积累速率并不相同。陈宏等研究发现,开花期以前是小麦单茎干物质积累量的缓增期,开花期至灌浆期干物质积累量急剧加快,灌浆期后减缓[9]。赵姣等通过Logistic曲线拟合越冬后小麦干物质的积累过程,表明孕穗至开花期小麦干物质积累速率最大,在拔节中后期之前和花后20 d,干物质积累缓慢增加[10]。此外,张黛静等通过13C稳定性同位素示踪技术发现,不同取样时期小麦净输入光合13C总量范围为21.71~94.06 mg/盆,随着生育期的推进,叶和茎鞘的光合碳逐渐向穗部转运,故叶和茎鞘部13C含量减少,穗部13C含量增加,表明小麦生长发育各个阶段,干物质在不同器官的积累量也不同[11]。
1.2 小麦干物质分配规律
干物质在小麦各器官中的分配对于建立平衡的源库关系及指导小麦高产优质生产具有重要意义。开花期小麦干物质在不同器官中分配比例一般表现为茎鞘>叶>穗,且茎鞘和穗的干物质积累量与成熟期小麦植株干物质量显著相关[12]。郭增江等研究表明,灌浆前期籽粒中同化物占总生物量的35.25%~45.66%,颖壳+穗轴占7.40%~9.85%,茎秆占27.58%~39.75%;灌浆后期植株70%以上的同化物运送至籽粒中[13]。Zhang等研究表明,成熟期小麦籽粒、茎叶、穗轴+颖壳和旗叶的13C分布所占比例分别为40.89%、31.09%、4.50%和2.19%[14]。此外,小麦各器官干物质分配率达到峰值的时间不同。叶片的分配比例在拔节期达到最大,而后随着生育进程的推进呈现出不断降低的趋势,茎鞘所占比例呈先增后降的变化趋势且在孕穗期达到最大;穗所占比例呈不断增加趋势,并在成熟期达到最大值[15]。小麦干物质在源库之间的合理分配利用对发挥最大产量潜力至关重要,应通过栽培措施的优化来改善源库关系,从而协调干物质在源库间的分配,实现小麦的稳产、丰产与优质。
1.3 小麦干物质积累、分配与产量的关系
小麦产量形成的过程,实质上是同化物的生产制造及转运分配的过程。小麦籽粒产量主要来自于花前贮存的同化物向籽粒的转运及花后叶、茎鞘等营养器官中光合同化物的积累[16]。花前是小麦营养生长的重要阶段,此时干物质的积累可为小麦产量的形成奠定基础。研究发现,拔节-开花期小麦群体干物质积累量与总结实粒数呈显著二次曲线关系,说明此时期干物质积累量不宜过大,否则会导致无效分蘖增多,不利于建成平衡的群体生态结构,从而导致减产[17]。杨佳凤等研究指出,孕穗-开花期干物质积累量与籽粒产量呈二次曲线关系,此时期适宜的干物质积累量是形成高产的基础[18]。花后是籽粒灌浆的关键时期,该阶段较高的干物质积累量能保证小麦正常的灌浆能力[17]。研究发现,小麦花前干物质转运量对产量的贡献率为43.10%~45.22%,与产量呈线性负相关;花后干物质积累量对产量的贡献率为54.78%~56.92%,与产量呈线性正相关[19]。因此,小麦群体后期干物质积累量是实现高产的关键,在合理调控拔节-开花期群体干物质积累量的基础上,应重点扩大花后干物质积累量。
2 小麦干物质积累与分配的影响因素
2.1 基因型对小麦干物质积累与分配的影响
因发育特点不同,不同基因型小麦的“源”“库”器官在调控干物质转运分配的作用上存在一定差异(图1)。研究表明,小麦基因型是影响干物质转运和分配最主要的因素[20],不同品种小麦花后干物质的积累量和花前干物质的运转能力对籽粒的贡献率存在差异[21]。吴金芝等研究发现,晋麦47较偃展4110花前积累的干物质多,花前干物质运转能力强,且花前贮藏干物质对籽粒的贡献率大,从而获得较高产量[22]。仝锦等研究表明,超高产品种能显著提高花后干物质积累量及其对籽粒的贡献率,最终产量较高[23]。
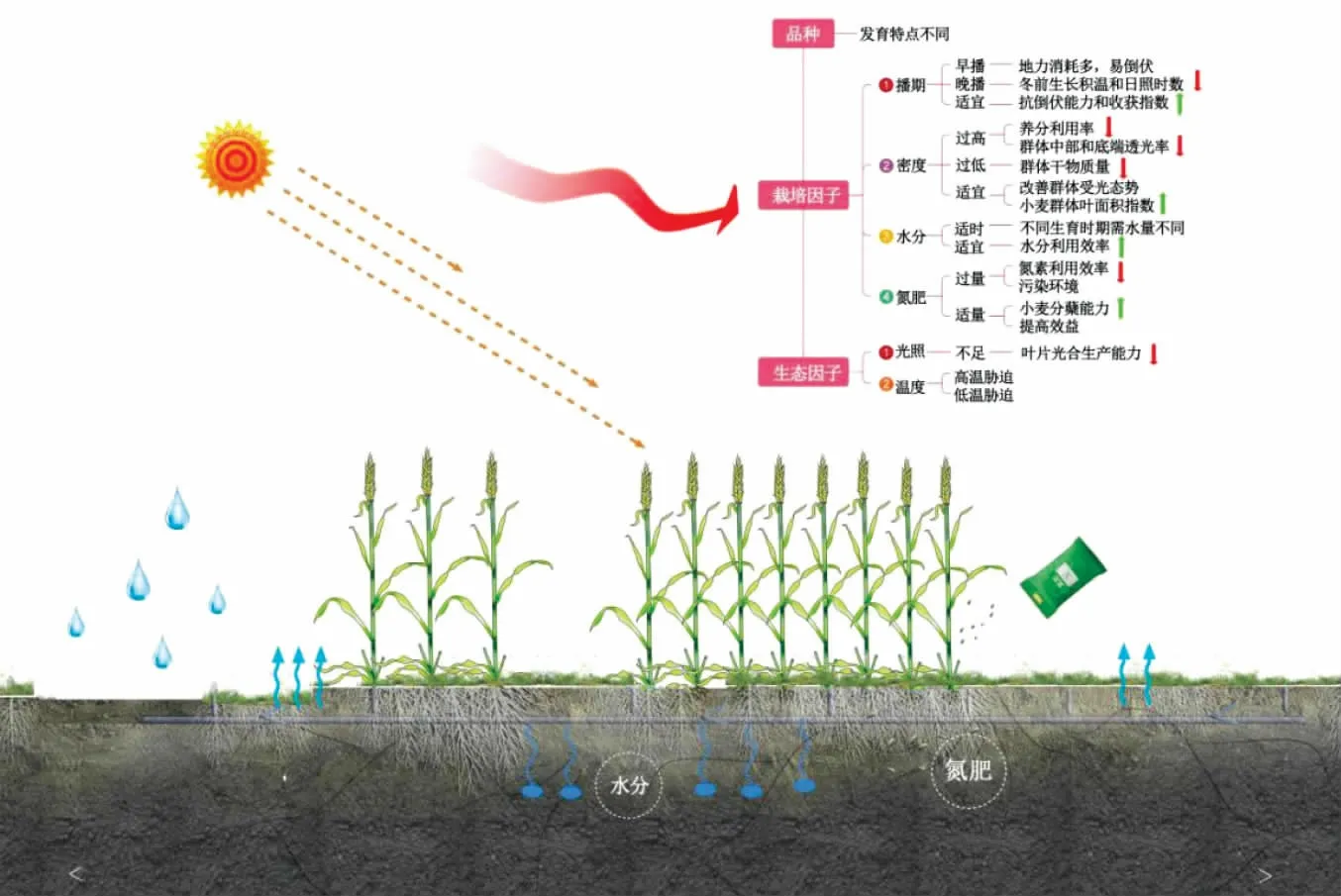
图1 影响小麦干物质积累与分配的因素
此外,不同基因型品种小麦的干物质积累速率不同,从而影响成熟期小麦籽粒产量。张利等研究发现,晚熟品种小麦较早熟品种的干物质积累平均速率高,最大速率也明显高于早熟品种,从而具有更强的生产潜势[24]。李瑞珂等研究指出,高产小麦品种花前和花后干物质积累速率较低产小麦分别高2.5倍和1.6倍,适宜的干物质积累速率有利于小麦的高产优质[25]。因此,掌握不同基因型小麦干物质积累及分配特性,通过改变其遗传特性选取高产高效型小麦品种,可增加小麦“源”器官的生产能力和“库”器官的容量,促进强“源”畅“流”扩“库”,从而实现高产。
2.2 栽培因子对小麦干物质积累与分配的影响
2.2.1 播期。播期主要通过影响小麦生育前期的积温及光照等生态条件,使小麦生长发育过程中干物质的运转分配也相应发生变化,从而影响小麦个体生长和群体建成[26]。播期不同导致小麦冬前积温不同,造成苗期单株和群体分蘖的干物质积累量存在较大差异,进而影响小麦后续干物质的积累和产量形成。研究表明,因气温连续偏高,早播使小麦冬前出现旺长,甚至提前拔节,导致小麦遇春季低温易遭受冻害,进而使群体干物质积累量下降[27]。播期推迟,使小麦冬前生长所需的有效积温大幅度降低,导致小麦冬前生长量不足,虽返青后干物质积累速率加快,但无法弥补叶面积减少所带来的干物质积累量的损失,因此难以形成高产群体基础[28-29]。适期晚播可提高小麦拔节—成熟期干物质生产量,从而增加成熟期干物质积累量,并结合较高的抗倒伏能力和收获指数获得较高产量[30]。气候、土壤及农民耕种习惯等因素均会影响播期,从而影响小麦最终经济产量。因此在实际生产中,应通过多年播期试验,结合作物品种、气候、土壤条件、栽培措施等因素,同时考虑播期与密度、水分及肥料等因素的互作效应,探究播期对小麦干物质积累与分配的调控效应,以实现最优的栽培因子组合,为小麦的高产优质提供理论依据。
2.2.2 密度。密度作为高产栽培技术措施中重要的一环,主要通过影响小麦群体结构和光热资源的分配来影响小麦产量形成[31]。密度过高会降低小麦对养分的利用率,加剧群体内部竞争,不利于产量形成[32];群体密度过低,小麦干物质在各生育时期积累量均较低,同样不利于群体产量的提高[33]。因此,明确密度与干物质积累与分配之间的关系,对于构建合理的群体结构从而实现小麦高产具有重要意义。刘娟等基于归一化法,认为小麦干物质积累中期(快增期)是干物质积累速率对密度的敏感反应期[34]。而播量过大会降低小麦叶片的光能利用率,过度消耗土壤水分,导致叶片的光合能力下降,干物质积累减少。丁位华等研究发现,播量过大会使小麦整个生育期群体光合速率较高,但中部和底端透光率较小,光能利用率较低,从而导致小麦群体干物质积累量和产量降低[35]。Reddy等认为,过高的种植密度会消耗过多的土壤水分,使小麦开花期土壤含水量较低,导致气孔导度和光合能力下降,从而影响干物质的积累[36-37]。适宜播量会改变小麦群体结构,改善群体受光态势,促进群体对光能充分利用,增加干物质的积累,从而提高产量[38]。因此,适宜的密度会提高小麦群体数量和叶面积指数,延长叶片光合功能期,优化群体光合性能,从而促进小麦干物质的积累。
2.2.3 水分。不同生育时期小麦对水分的需求不同,土壤水分过多或不足及灌水时期都会对小麦干物质的积累与分配造成影响。Zhang等研究表明,灌溉至季节性水分蒸发量的94%时,小麦成熟期干物质积累量最高[39]。姚宁等研究发现,在越冬期和返青期对小麦进行干旱处理,拔节后生物量显著降低且后期复水无法弥补[40]。韩惠芳等研究指出,拔节期灌溉有利于小麦贮藏物质的积累,同时拔节水和抽穗水更有利于提高贮藏物质的转运效率[41]。此外,水分胁迫对小麦不同营养器官干物质累积的影响不同,其中对叶片的影响最大,对穗的影响最小[42],复水后小麦植株各器官的干物质累积和产量均有不同程度的恢复[43]。
水分是小麦干物质积累与分配的重要限制因子,不同的灌水处理造成土壤水分的差异,而小麦营养器官贮存的同化物受土壤水分含量的调控[44]。适时适量灌溉可缓解土壤水分缺乏,提高水分利用效率,促进小麦干物质积累[45]。谷艳芳等研究发现,干旱胁迫下,冬小麦植株和器官水平上干物质积累量均显著下降,干物质分配趋向生长中心,花后同化物的积累量减少50%,但营养器官干物质的转运量会增加30%以上[46-47]。适宜的水分有利于增加小麦地上部植株和不同器官(叶片、茎鞘、颖壳)的干物质量,促进籽粒产量形成[48]。因此在小麦生产中,可在小麦关键生育时期通过合理供水来提高水分利用效率,从而增加干物质积累量和调控物质转运,最终提高产量。
2.2.4 氮肥。氮肥是调节小麦干物质转运能力从而实现小麦高产栽培的重要养分限制因子。合理增施氮肥对小麦器官建成和营养物质分配有正向调控作用,能够协同提高干物质积累量和干物质转运能力,从而实现高产[49]。研究表明,在一定范围内适量增施氮肥可以提高小麦花后干物质积累量及其向籽粒中的转运效率[50]。蔡瑞国等研究指出,适量施氮提高了小麦干物质积累量,同时促进花前贮存干物质向穗部的转运,有利于产量三因素的协调发展[51]。李娜等研究表明,适宜的施氮量有助于增强植株生长后期光合作用能力和提高分蘖能力,协调小麦花前和花后干物质积累,促进营养物质向籽粒快速转移,从而提高花后光合产物对籽粒的贡献率,对小麦增产具有重要意义[52]。
然而,在小麦生产中为追求高产过量施用化肥的现象屡见不鲜,这不仅极易造成氮素损失,导致小麦氮素利用效率以及氮农学利用效率降低,不利于资源可持续发展,同时也给农田环境带来巨大压力[53-54]。研究表明,目前农业系统中的氮肥盈余量已经达到175 kg/hm2[55],过量施氮使群体结构变差,导致干物质积累量减少,小麦花后物质生产量增幅较小甚至下降[52]。因此,根据地区和品种的不同来判断最适宜的氮施用量,不仅可以有效控制成本、提高效益,还可以合理分配和充分利用氮肥资源,减少农业生产过程中养分损失和对环境造成的负面影响,使小麦同化物合成、贮存和转运三者相协调,挖掘小麦产量潜力。
2.3 生态因子对小麦干物质积累与分配的影响
2.3.1 光照。光照是影响小麦干物质积累和产量的重要环境因子,小麦的生产潜力很大程度取决于接受太阳光照的多寡以及光能利用率的高低。在黄淮麦区,雾霾、阴雨寡照等因素引起的弱光问题已成为限制小麦高产的重要因素[56],弱光造成小麦接受有效光合辐射减少,使光化学反应速率和气孔开度下降,从而影响小麦干物质的积累。据报道,黄淮海地区的光照度以每年0.6%的速度下降,导致小麦叶片光合速率降低,严重影响小麦生产[57]。顾蕴倩等通过构建SUCROS模型发现,日总光合有效辐射低于3.71 MJ/m2且持续4 d以上对小麦干物质的积累和产量产生显著影响[58]。此外,弱光会降低小麦“源”器官叶片干物质的生产能力,同时阻碍干物质向“库”器官穗部的转运,最终导致产量降低。Xu等研究发现,弱光会影响小麦干物质积累和同化物向籽粒的运输,导致产量下降[59]。刘希伟等研究指出,花后不同阶段遮光处理均会导致小麦光合速率下降,干物质生产不足,籽粒灌浆速率降低,进而影响产量[60]。李刘龙等研究发现,弱光条件下籽粒的“库吸力”降低,使植株贮藏的同化物更多地滞留在营养器官中,用于维持“源”器官的功能代谢,导致干物质在小麦“库”器官穗部干物质的积累量减少[61]。因此,光照不足会使小麦功能叶片光合产物供应不足,直接影响到干物质的积累与分配,从而导致减产。
2.3.2 温度。温度是小麦进行营养生长和生殖生长的重要生态因子,小麦完成其生命周期需要一定的积温[62]。马倩倩等通过皮尔逊相关性分析等方法研究发现,小麦全年≥10℃的积温为3 500℃·d左右,播种-成熟期>0℃积温约为2 200℃·d[63]。小麦的生长发育在不同的生育时期所要求的温度也不同。小麦播种的适宜温度为15~16℃,而出苗后温度需要降到5~6℃,拔节到抽穗期则要求温度逐渐升高,最后达到21~22℃[64]。然而,全球气候变暖的加剧导致极端温度事件频发,严重威胁到小麦生产[65]。高温胁迫会加速叶片衰老,缩短叶片中光合产物向籽粒中的运输时间,导致籽粒灌浆终止早于叶片完全衰老,叶片光合产物被分配到其他营养器官中[66],抑制了籽粒中干物质的积累,最终影响粒质量和产量。张姗等研究发现,高温胁迫导致的花后干物质积累量和花前干物质运转率共同降低可能是籽粒产量降低的主要原因[67]。
而低温则会导致小麦叶片的光合速率、蒸腾速率和叶片气孔导度显著下降,胞间CO2浓度大幅升高,导致干物质积累减少,从而造成减产[68]。张溪荷等研究发现,低温胁迫导致茎秆干物质向穗部的转运受阻,营养物质累积在茎秆中,所以茎秆分配比例升高,穗分配比例降低[69]。因此,温度胁迫主要通过减少小麦籽粒灌浆时间和降低叶片光合速率等一系列生理生化过程来降低干物质的积累量和分配,最终导致小麦减产。
3 展望
3.1 运用分子生物学技术选育小麦新品种
小麦的产量和品质是受多基因和环境互作影响的复杂数量性状,单纯依靠现有常规育种技术耗时长、成本高[70],难以满足全球日益增长的粮食需求,小麦基因组和分子遗传育种的发展为进一步解决世界粮食问题提供了新的契机和突破口。因此,应利用CRISPR/Cas9基因编辑技术[71]和分子标记辅助育种,借助转录组、蛋白质组、代谢组等多组学技术加快对小麦功能基因的定位,从而加快推动种质资源的利用和新品种的选育。
3.2 利用作物表型组学、作物模型等探索小麦干物质积累过程机理
定量分析小麦生长过程中各项指标的动态变化是揭示产量形成和掌握群体调控指标的重要内容。高通量表型组学通过从微观到宏观、从超微到器官再到个体乃至群体的多维度数据以实现对作物不同层次的监控,从而获取作物不同生长发育阶段的动态性状,对作物群体的早期诊断和定量调控具有重要意义。今后,应充分利用各种作物表型数据库,并结合Richards、Logistic、Compertz等生长模型探索干物质积累过程机理[71-73],深入研究各影响因子与干物质积累的定量关系,为提升小麦干物质积累的途径提供新思路。
3.3 集成多项生产技术实现农业可持续发展
随着农业资源供需矛盾的加剧,小麦生产的可持续性成为研究的焦点。未来应集成多项节水、绿色、高效生产技术模式,通过推广自动驾驶、卫星平地、精量播种、精准施肥、智能灌溉、无人植保、智慧收获等多项智能化技术,推动小麦生产向节水、稳产、绿色、优质、高效种植发展,并且结合地区的气候特点、灾害发生规律以及生产条件等,在提高系统生产效率和可持续性的基础上实现精细化农业管理,从而实现小麦生产的可持续发展。
