延衰组合物对酵母细胞寿命及氧化应激的研究
2021-07-15王敏廖林锋孙晓宇任娇艳
王敏,廖林锋,孙晓宇,任娇艳,2*
(1.华南理工大学食品科学与工程学院,广东 广州 510641;2.中新国际联合研究院,广东 广州 510000)
衰老是生物随着时间的推移, 自发的必然过程,也是应激、劳损、营养失调和功能障碍等导致的结果[1]。 目前,关于抗衰老及相应抗衰老剂方面的研究对人类生命、社会、经济具有重要价值,仍需积极探索。衰老机制的研究是生命科学领域的前沿课题,由于机体衰老过程受到外界各种因素的干扰,所以衰老研究非常复杂。 酵母细胞与人类的体细胞非常相似,酵母细胞作为衰老的研究模型具有诸多优点:生活周期较短、遗传背景简单、基因组已完全破译,可进行遗传操作,重要的是酵母细胞内与衰老相关的新陈代谢及信号转导通路具有高度的保守性[2-4]。 几十年来,酵母细胞应用于衰老方面研究取得了较大进展。 在能量限制条件下,酵母细胞会退出有丝分裂细胞周期,并获得一系列特定的静息细胞特性以延长寿命,且抗逆性增加[5]。 酵母细胞内激酶(Rim15、Mck1、Yak1)、复合物SNF1/AMPK、细胞壁完整性通路以及许多细胞周期调节因子被证明是酵母建立适当的静息状态和延长其时序寿命所必需的。 此外,研究发现减少酵母细胞内的毒性氧化物[如活性氧(reactive oxygen species,ROS)]可显著延长酵母细胞寿命[6]。 因此,利用酵母细胞进行延衰活性物质筛选及其延衰机理研究,具有可行性和优越性。
中国的传统植物资源丰富,从中寻找高效、低副作用的活性物质具有重要的研究价值。 西洋参又称美国人参、花旗参,为五加科人参属植物,其性凉,味甘、微苦,具有补气养阴、清热(虚火)生津的功效[7];五味子味酸敛,可益气敛阴补肾,收耗散之气[8];葛根、麦冬和茯苓,具有健脾生津利水的功效[9-11]。 现代药理学研究表明,西洋参、五味子、葛根、麦冬和茯苓中均含有诸多抗氧化活性成分(如皂苷、多糖、多酚、氨基酸等),可有效清除衰老过程中的自由基, 减少机体的氧化损伤,因而具有潜在的延衰功效[12-14]。 但衰老是多组织和器官的问题,涉及的治疗靶点往往不止一个,因而单一活性成分研究已不能满足其要求。 近年来,基于中医药理论利用各原料进行科学合理的配伍而获得的组合物及其机理研究逐渐受到广泛的关注,并取得了显著的成效。 组方的配伍,一方面可以发挥协同作用,提高疗效以适应复杂多靶点的治疗;另外一方面可发挥拮抗作用,减少不良反应以利于治疗[15]。基于上述背景,本文将西洋参、五味子、葛根、麦冬和茯苓进行组合配比,利用酵母细胞模型,探讨单一原料及其组合物在延缓衰老方面的效果,这将为天然植物活性物质在研发延缓衰老的功能性食品和药品方面提供有益的参考。
1 材料与方法
1.1 材料与试剂
酵母细胞(野生型菌株BY4742、 共转染质粒pRS426-pSSA3-RFP 株和共转染质粒pRS425-pHsp26-VFP 株) 均由剑桥大学生物化学系Nianshu Zhang 教授提供;细菌培养用酵母抽提物(微生物用)、D-(+)-葡萄糖(生化试剂)、琼脂(生化试剂)、不含氨基酸的酵母氮碱(微生物用)、甘油(生化试剂)、聚乙二醇4000(生化试剂)、硫酸腺嘌呤、L-盐酸组氨酸、L-色氨酸、L-亮氨酸、L-盐酸赖氨酸:BBI Life Science 公司;西洋参片(批号19041705)、葛根(批号Y181008-01): 安国市一方药业有限公司; 五味子(批号181201): 广东杏林药业有限公司; 茯苓(批号Y180208-01、QKFL181104): 安徽金寨乔康药业有限公司;麦冬(批号180701):四川新荷花中药饮片股份有限公司。 蛋白胨(微生物用)、乙酸锂二水(分析纯)、三羟甲基氨基甲烷-乙二胺四乙酸缓冲液(tris-ethylene diamine tetraacetic acid,TE,分析纯):生工生物工程(上海)股份有限公司;丙氨酸、精氨酸、天冬酰胺、天冬氨酸、半胱氨酸、谷氨酰胺、谷氨酸、甘氨酸、肌醇、赖氨酸、异亮氨酸、甲硫氨酸、对氨基苯甲酸、苯丙氨酸、脯氨酸、丝氨酸、酪氨酸、苏氨酸、缬氨酸:上海源叶生物科技有限公司; 二氢罗丹明123(Dihydrorhodamine 123,DHR123,D1504)、海藻糖酶(2.6U/mg 蛋白)、淀粉葡萄糖苷酶(30 U/mg 蛋白~60 U/mg 蛋白)、葡萄糖检测试剂盒(GAGO-20):Sigma-Aldrich 公司;碘代3,3'-二己氧基羰花青试剂盒(3,3'-dihexyloxacarbocyanine Iodide,DiOC6(3)):Abbkine 科技有限公司;乙醇、浓硫酸等试剂均为分析纯:广州从源试剂有限公司。
1.2 仪器与设备
全温振荡培养箱(HZQ-F160):太仓市强乐实验设备有限公司;涡旋混合仪(XW-80A):麒麟贝尔实验室仪器有限公司;水浴恒温振荡器(THZ-82A):常州澳华仪器有限公司;分析天平(FA2104N):上海民桥精密科学仪器有限公司;高压灭菌锅(LDZC-40BI):上海申安医疗器械厂;超净工作台(SW-CJ):苏州安泰空气技术有限公司;高速冷冻离心机(H2050R):长沙湘仪离心机仪器有限公司; 全功能微孔板检测酶标仪(Synergy H1):美国伯腾仪器有限公司;流式细胞仪(CytoFlex):美国贝克曼库尔特公司;超声波清洗器(KQ5200DE):昆山市超声仪器有限公司。
1.3 方法
1.3.1 原料及组合提取物粉末的制备和配制
西洋参、葛根、五味子、麦冬和茯苓提取物制备:称取原料各100 g,粉碎过60 目筛,以1∶10(g/mL)的料液比加入65%乙醇进行微沸回流提取2 h, 过滤后取上清液;过滤后的滤渣继续以1∶8(g/mL)的料液比加入65%乙醇进行二次微沸回流提取,提取时间为2 h,过滤弃渣,5 000 r/min 离心15 min,合并滤液,(60±3)℃真空浓缩,冷冻干燥,干燥器避光保存备用。
组合提取物制备:将西洋参、麦冬、茯苓、葛根、五味子按4∶2∶3∶4∶3 的质量比例分别称取25、12.5、18.75、25、18.75 g 原料粉末(过60 目筛),混匀,以1∶10(g/mL)的料液比加入65%乙醇进行微沸回流提取2 h, 过滤后取上清液;混合物滤渣继续以1∶8(g/mL)的料液比加入65%乙醇进行二次微沸回流提取,提取时间为2 h,过滤弃渣,5 000 r/min 离心15 min, 合并滤液,(60 ±3)℃真空浓缩,冷冻干燥,干燥器避光保存备用。
提取物液配制:各提取物粉末用液体的酵母浸出粉胨葡萄糖培养基(yeast extract peptone dextrose medium,YPD)溶解成终浓度为2 mg/mL 的母液,此母液经0.22 μm 的无菌滤头过滤,放置4 ℃冰箱中保存备用。
1.3.2 酵母细胞培养
接种环于酒精灯外焰灼烧灭菌后,从装有野生型菌株(BY4742)的甘油管中蘸取一环菌液,以平板划线方式在固体酵母平板YPD 培养基上逐渐稀释菌液,划线后的固体平板在30 ℃培养箱中培养2 d。 取出划线生长2 d 后的固体YPD 平板, 挑单菌落于装有5 mL~10 mL YPD 液体培养基的50 mL 试管中, 置于30 ℃、200 r/min 的摇床中过夜培养,用于后续试验[5]。
1.3.3 酵母细胞增殖检测
将过夜培养后的BY4742 菌液以1∶50(体积比)的接种比例接种于含药物浓度为250、500、1 000 μg/mL的YPD 培养液中,并设置各自溶剂对照,以200 μL/孔的液体量培养于96 孔板中,每个浓度设置4 个平行复孔, 于酶标仪中, 在30 ℃下以连续振板方式培养,在OD595nm下每15 min 读数一次,培养24 h,计算代时(即酵母繁殖一代所需时间)[5]。
1.3.4 酵母细胞时序寿命(chronological lifespan,CLS)检测
将酵母株按1∶50(体积比)接种在含1 000 μg/mL各原料和组合提取物的酵母氨基酸缺陷型合成培养基(yeast synthetic drop-out medium,SD)中振荡培养8 d,分别在培养的3 d 和8 d 后取出酵母培养液,采用单克隆计数法(colony forming units,CFU)测定干预后的酵母BY4742 株形成单菌落的能力。 干预后的酵母细胞按照1∶10、1∶50 和1∶250 的稀释梯度不断稀释,将最终稀释125 000 倍的酵母细胞液涂琼脂平板,在30 ℃条件的培养箱中生长3 d, 拍照并使用Image J 软件进行单菌落计数统计。 酵母细胞的存活率计算:将每组培养3 d 后的酵母形成单菌落个数标记为A,培养8 d 后酵母形成单菌落个数标记为a,3 d 后计数得到的单菌落数酵母细胞存活率为(A/A×100)%,标记为CLS0,此时各组的CLS0 存活率均为100%,8 d 后酵母细胞存活率为(a/A×100)%(即为相对CFU%)[16]。
1.3.5 酵母细胞内活性氧检测
将酵母株按1∶50(体积比)接种在40 mL 浓度为1 000 μg/mL 的组合提取物培养液中,在30 ℃、200 r/min的摇床中培养。 培养24 h 后,取样检测酵母细胞活性氧水平。 将OD595nm值为0.5~0.6 的酵母培养液,25 ℃、11 000 r/min 离心1 min,收集沉淀酵母细胞。用200 μL磷酸盐缓冲液(phosphate buffer saline,PBS)洗涤1 次,然后重悬于200 μL PBS 中。将50 μL 细胞加入到含有10 μmol 二氢罗丹明123(DHR 123)的1 mL PBS 中。避光,并在30 ℃下孵育60 min,用200 μL PBS 洗涤细胞一次,并重悬于1 mL PBS 中。在轻度超声处理后,使用流式细胞仪[激发波长:488 nm,发射波长:(530±30)nm]检测细胞荧光强度[17]。
1.3.6 酵母细胞内线粒体膜电位(mitochondrial membrane potential,MMP)检测
将酵母株按1 ∶50(体积比)接种在40 mL 浓度为1000μg/mL 的组合提取物培养液中,在30℃、200 r/min的摇床中培养。 培养24 h 后,取样检测酵母细胞活性氧水平。 将OD595nm值为0.5~0.6 的酵母培养液,25 ℃、11 000 r/min 离心1 min,收集沉淀酵母细胞,用200 μL PBS 洗涤1 次,然后重悬于200 μL PBS 中。将100 μL细胞加入到0.9 mL 用PBS 稀释后终浓度为200 nmol/L DiOC6(3)荧光探针中。 细胞悬液在30 ℃下避光孵育30 min,用PBS 洗涤一次并重悬于PBS 中,用流式细胞仪(激发波长:482 nm,发射波长:504 nm)中异硫氰酸荧光素(fluorescein isothiocyanate,FITC)通道检测细胞荧光强度。
1.3.7 酵母细胞内应激转录因子Msn2/4、Gis1 和Hsf1激活程度的检测
利用含2%葡萄糖的酵母基础培养基(yeast synthetic condition minimal medium,SC) 复苏已转化质粒(pRS426-pSSA3-RFP 与pRS425-pHsp26-VFP) 的酵母菌株液,30 ℃、200 r/min 过夜培养。 将过夜培养的含质粒酵母细胞以1∶50(体积比)接种在低糖(0.6%葡萄糖)SC 培养基配制的含1 000 μg/mL 组合提取物培养液中,干预24 h 后利用流式细胞仪测定酵母细胞中由启动子控制的应激转录因子Msn2/4、Gis1 和Hsf1 的激活荧光变化。 2 000 r/min 离心5 min 收集干预后的酵母细胞,用PBS 清洗1 遍后,再用PBS 重悬。流式细胞仪上机检测,红色荧光蛋白(red fluorescent protein,RFP)的激发波长为(580±10)nm,发射波长为(610±10)nm;绿色荧光蛋白(vivid Verde fluorescent protein,VFP)的激发波长为(500±10)nm,发射波长为(540±10)nm[5]。
1.4 数据处理与统计分析
所有数据均进行平行测定3 次,数据结果以平均值±标准误的形式表示,采用Prism 6 软件对试验数据进行统计学分析,两组之间采用t 检验,多组之间通过方差分析(one way ANOVA)做显著性分析,p <0.05 为有统计学意义,图表采用GraphPad Prism 6 进行绘制。
2 结果与分析
2.1 不同原料提取物及组合提取物对酵母细胞生长的影响
代时表示酵母细胞分裂一次所需的时间,表征酵母细胞在药物或者溶剂下的生长情况。 酵母细胞生长曲线图见图1。
如图1 所示, 酵母细胞在0~5 h 处于生长的延滞期,即适应环境及慢速生长时期;5 h~10 h 处在生长对数期,酵母细胞汲取营养,营养消耗速率较快,酵母细胞快速生长,细胞数量以指数形式增加;在12 h 后,酵母细胞群体达到稳定,处于细胞生长的稳定期,细胞数量处于动态变化,整体数量基本不变。 数据处理选取了对数期(5 h~10 h)细胞生长数据,根据指数期生长曲线公式,计算细胞生长代时。
各原料提取物及组合提取物在3 个浓度(250、500、1 000 μg/mL)下对酵母细胞生长的影响见图2。
由图2 可知, 不同浓度下的西洋参、 五味子、麦冬、葛根及组合提取物对酵母生长没有明显的抑制或者促进作用(p>0.05)。 此外,茯苓干预组在浓度为500 μg/mL~1 000 μg/mL 范围内可显著抑制酵母细胞生长(p<0.05)。
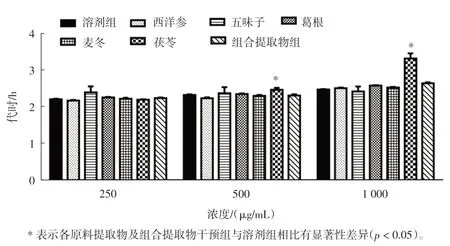
图2 各原料提取物及组合提取物对酵母细胞生长的影响Fig.2 Effects of individual extracts and combined extracts on the growth of yeast cells
2.2 不同原料提取物及组合提取物对酵母细胞时序寿命的影响
在裂殖酵母中,细胞寿命可分为两种形式:一种是复制寿命(replicative lifespan,RLS),指母细胞在衰老前可产生子细胞数量;另一种是时序寿命(CLS),指不分裂的细胞在稳定期中所能存活的时间[18]。 而酵母细胞时序寿命形式更能反映哺乳动物细胞在分裂完成后存活的时间长短,反映哺乳动物细胞寿命,因此,采用时序寿命指标评价各单一原料提取物及其组合提取物的延衰活性。
试验中检测了西洋参、五味子、麦冬、茯苓、葛根和组合提取物分别干预8 d 后,其酵母细胞存活情况,即形成单克隆菌群的能力,结果如图3 所示。
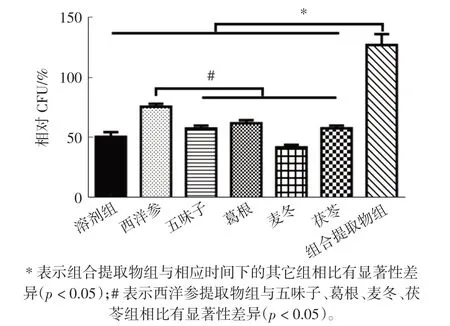
图3 各原料提取物及组合提取物对酵母细胞时序寿命的影响Fig.3 Effects of individual extracts and combined extracts on the CLS of yeast cells
由图3 可知,与溶剂组(51.00±3.94)%相比,在组合提取物干预酵母细胞8 d 后, 该组的细胞存活率为(126.81± 5.22)%(p<0.05);而五味子、葛根、麦冬、茯苓单一提取物组与溶剂组无显著性差异(p>0.05),与组合提取物组存在显著性差异(p<0.05)。 这些结果表明,相比于组合物中各单一提取物,组合提取物可显著延长酵母细胞的寿命, 其延衰效果优于单一原料,说明组合的原料之间具有潜在的协同增效作用。 此外,研究发现,西洋参组的延长酵母时序寿命效果优于其它原料单一成分效果(p<0.05),这说明组合物中延衰成分可能主要来自于西洋参。
2.3 组合提取物对酵母细胞内活性氧水平的影响
文献报道指出酵母的寿命与细胞内活性氧水平等密切相关[19]。 活性氧(ROS)的产生是造成细胞氧化损伤的一个主要原因, 伴随着衰老的发生,ROS 的产生逐步增加,引起胞内脂质、蛋白及核酸等大分子的损伤积累,最终可导致细胞功能损伤,引起细胞死亡,而增加细胞的抗氧化能力可以延长多种生物的寿命[20]。试验采用一种自由基荧光染料罗丹明123(DHR123)检测胞内的ROS 积累情况,根据活细胞中荧光的产生情况,可以判断ROS 含量的多少及变化。 组合提取物处理对酵母细胞内ROS 水平的影响见图4。

图4 组合提取物处理对酵母细胞内ROS 水平的影响Fig.4 Effects of the combined extracts on intracellular ROS levels of yeast cells
如图4 所示:与溶剂组(100 ± 3.74)%相比,组合提取物可高度显著降低酵母细胞内的ROS 水平(82.17±2.48)%(p<0.001),起到较好的细胞内抗氧化效果。
2.4 组合提取物对酵母细胞线粒体膜电位的影响
线粒体膜电位(MMP)是评价线粒体产能作用的重要参数,是维持氧化磷酸化(oxidative phosphorylation,OXPHO)期间能量储存过程中的重要因素[21]。 酵母线粒体膜电位的测定选择一种具细胞膜渗透性、发绿色荧光的亲脂性探针DiOC6(3)可在低浓度下偏向性选择染色线粒体。 组合提取物处理对酵母细胞内线粒体膜电位的影响见图5。
由图5 可知, 组合提取物干预组的相对荧光强度(54.44±1.23)%高度显著低于溶剂对照组(100±1.44)%(p<0.001)。 线粒体膜电位越高,线粒体产生的能量越多,促进细胞能量转化。 因而,组合提取物可通过降低线粒体膜电位, 使酵母产生和释放的能量显著减少。越来越多的研究指出, 热量限制能够有效延长寿命,这已经在多个物种(如酵母、线虫、果蝇和猴子等)中得以证实[5,22-24]。 此外,能量限制可通过整合多个通路信号,来调节酵母细胞的时序寿命[5]。 因此,组合提取物可通过调节线粒体膜电位以限制酵母细胞的能量,延长酵母细胞的时序寿命。
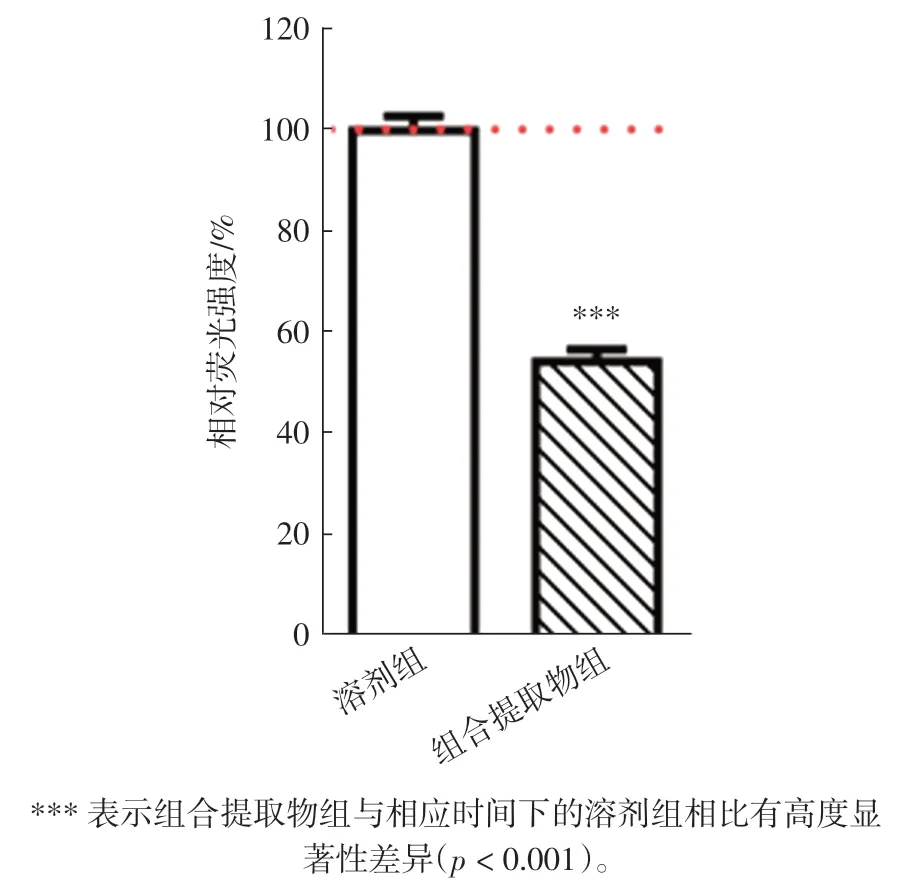
图5 组合提取物处理对酵母细胞内膜电位水平的影响Fig.5 Effects of the combined extracts on intracellular MMP levels of yeast cells
2.5 组合提取物对酵母细胞内应激转录因子Msn2/4、Gis1 和Hsf1 激活的影响
当缺乏葡萄糖或任何其它大量营养物质时,酵母细胞会退出有丝分裂细胞周期,并获得一系列特定于静息细胞的特性,以确保其寿命[5]。 由于TOR1Δ 或CH9Δ 缺失所导致酵母细胞时序寿命CLS 延长也依赖于Rim15 激酶及其下游效应物Msn2/4 和Gis1 来激活应激反应[25]。 Wei 等[25]预测除Rim15 激酶外,其它因素也参与了通过限制热量来调节应激反应和延长寿命的过程。 Zhang 等[5,26]为了便于自动识别调节应激反应的这些因子,采用两种“饥饿”诱导的报告基因(pHSP12-HSP12-VFP 和pSSA3-RFP)筛选酵母基因组中编码大多数“信号”分子的272 个基因缺失突变体,发现酵母细胞内ROS 的清除(应激反应)和提供能量的碳水化合物积累(能量)是受到转录因子Msn2/4、Gis1和Hsf1 的激活调控的。 转录因子Msn2/4、Gis1 和Hsf1的激活一方面可抑制酵母细胞内ROS 产生,另一方面可促进酵母细胞内碳水化合物的积累。
为了进一步探讨组合物延长酵母时序寿命作用的机理,本试验中分别采用了两种不同质粒构建的报告基因酵母株(pRS426-pSSA3-RFP 与pRS425-pHsp26-VFP)[5]。 酵母细胞株中两种不同启动子连同的报告基因都被三类应激转录因子靶向:Msn2/4、Gis1 和Hsf1, 而这三类应激转录因子可协调抗氧化防御系统和分子伴侣的表达;并且带有红色荧光的RFP 报告基因或者绿色荧光的VFP 报告基因的表达水平高低反映出酵母细胞中的对应激的反应程度。 组合提取物对pSSA3-RFP 或pHsp26-VFP 启动子控制的酵母细胞株内应激转录因子Msn2/4、Gis1 和Hsf1 激活的影响见图6。

图6 组合提取物对pSSA3-RFP 或pHsp26-VFP 启动子控制的酵母细胞株内应激转录因子Msn2/4、Gis1 和Hsf1 激活的影响Fig.6 Effects of the combined extracts on the activation of Msn2/4,Gis1 and Hsf1 stress transcription factors in two different yeast cells with pSSA3-RFP or pHsp26-VFP promoters
结果表明(图6):与溶剂对照组的相对荧光强度(100%)相比,组合提取物组不仅可显著激活pSSA3-RFP 启动子控制下酵母细胞内的红色相对荧光强度显著增加(151.85 ± 3.50)%(p<0.001),还可促进pHsp26-VFP 启动子控制的酵母细胞内绿色相对荧光强度的显著性增加(154.44±5.13)%(p<0.001)。 同时,研究结果中红色和绿色荧光强度增加强度无显著差异,说明组合提取物具有较好的激活酵母细胞内应激转录因子Msn2/4、Gis1 和Hsf1 的功效。
3 结论
对数种植物原料提取物及其组合提取物进行筛选后发现,组合物对酵母细胞的时序寿命延长作用最为显著,在1 000 μg/mL 浓度作用下酵母细胞存活率可达(126.81±5.22)%,显著高于溶剂对照组和各单一原料提取物的存活率,说明组合物延衰效果优于组合物中任一单一原料的效果。 试验结果进一步表明,组合提取物可显著减少酵母细胞内ROS 水平和降低线粒体膜电位,并显著激活酵母细胞内与应激和碳水化合物积累密切相关的Msn2/4、Gis1 和Hsf1 转录因子。 通过试验结果分析可进一步推断,组合物不仅可通过激活细胞内的与抗氧化密切相关的转录因子Msn2/4、Gis1 和Hsf1 来清除细胞内的活性氧,起到抗氧化和延长酵母寿命的作用; 而且可通过激活转录因子Msn2/4、Gis1 和Hsf1 增加酵母细胞内的碳水化合物积累,降低线粒体膜电位,减少能量产生和释放,从而起到延长酵母时序寿命的作用。 这些结果表明该组方在应用于研发延衰、抗氧化、调节线粒体膜功能的食品、功能性食品以及药品中极具潜力。
