太岳山油松林火烧迹地恢复初期植物群落结构特征
2021-07-12武帅楷
金 山, 武帅楷
1 长治学院生物科学与技术系, 长治 046011
2 太行山生态与环境研究所, 长治 046011
森林火灾极大地影响了森林生态系统的土壤理化性质及生物群落的多样性、稳定性和动态变化[1],是世界性的重大灾害,而且中国是世界上森林火灾多发的国家之一[2]。我国森林火灾分布区域主要为西南、华中南、东南以及东北地区[3]。生物群落结构是生态系统研究中的基础且重要的一部分,在生物群落中,各种群协同进化,占据不同空间,使群落具有一定结构,包括垂直结构、水平结构和时间结构[4]。种类组成、物种多样性、群落类型、稳定性等用来表征群落结构特征,尤其是物种多样性,其既能反映群落的内部结构特征,也可以表现出群落的演替阶段及稳定程度,同时也是反映群落功能的重要指标,一直都是群落生态学和恢复生态学领域的研究热点[5]。火灾后,森林生态系统遭到不同程度破坏,开始新一轮演替,研究群落结构变化可以揭示火烧干扰后森林生态系统恢复的程度。国内外学者对火烧迹地的研究主要包括:森林火灾风险的预测及评估[6]、运用遥感等技术手段研究火烧迹地的时间和空间特征[7]、利用光谱分析法研究火灾严重程度[8]和火灾后植被盖度恢复[9]、火烧迹地土壤的斥水性和渗透性特征[10]、土壤温室气体特征[11]、土壤节肢动物[12]、马尾松(Pinusmassoniana)林和油松(Pinustabuliformis)林火烧迹地的土壤营养元素特征[13-15]、高山栎(Quercussemecarpifolia)林火烧迹地的土壤呼吸[16-17]、落叶松(Larixgmelinii)林火烧迹地地表CO2通量[18]、火烧迹地植物群落多样性[19]、群落演替[20]以及火烧对当地植被的影响[21]等。可见,目前学者们对火烧迹地的研究主要集中在土壤理化性质、土壤微生物[22]和植物群落等方面,但是已有的研究缺乏对不同火烧强度和不同恢复时间植物群落动态变化规律的系统研究,而且现阶段对火烧迹地植被恢复程度评价的研究也存在一定的局限性[23]。
山西太岳山林区在冬、春季容易出现干旱现象,极易发生森林火灾[24],但对该区域火烧迹地植物群落动态变化的相关研究很少。基于此,本研究应用群落相似性指数、多样性指数、稳定性系数、TWINSPAN数量分类和DCA排序等方法研究太岳山油松林火烧迹地植物群落天然恢复初期的物种组成、物种多样性、群落类型和稳定性,了解太岳山区油松林火烧恢复初期植物群落结构特征,揭示火烧后林下植被恢复规律,以期为太岳山火烧迹地植被的恢复和重建以及太岳山区油松林生态系统的科学管理提供一定的理论依据。
1 研究区概况
研究区(图1)位于太岳山北麓,地理坐标:112°11′—112°22′ E,36°43′—36°51′ N。区内沟壑纵横、地貌复杂,以黄土丘陵和土石山地为主,平均海拔约1500 m。气候属暖温带半湿润大陆性季风气候,年平均气温9℃,年平均降水量约650 mm,无霜期约125—130 d。主要植被类型为油松林和蒙古栎(Quercusmongolica)林,灌丛主要有虎榛子(Ostryopsisdavidiana)灌丛、黄刺玫(Rosaxanthina)灌丛、沙棘(Hippophaerhamnoides)灌丛等。研究区于2019年3月经历森林大火,过火面积达6.67 km2,火烧后植物的地上部分全部死亡,只剩油松枯立木,林下腐殖质层全部烧尽,但土壤结构比较完整,植物地下繁殖体受损较小。火烧迹地外围几乎未受干扰,仍为油松林。

图1 研究区位置
2 研究方法
2.1 采样方法
本文主要关注火烧迹地植物恢复生长一季后的草本层群落结构。于2019年10月初在研究区内选择7个样地进行调查(表1),样地平均坡度约30°,每个样地设置“100 m × 100 m”的调查区,定性记录调查区内出现的植物种名。在每个样地调查区内划分坡上、坡中、坡下3个坡位,在每个坡位分别设置3个“10 m × 10 m”的样格。在每个“10 m × 10 m”的样格中沿对角线设置3个“1 m × 1 m”的小样方,进行草本植物调查,记录每个小样方内草本植物的多度、高度、盖度等指标,同时记录经纬度、海拔、坡度、坡向等环境因子。

表1 样地信息表
2.2 计算公式
2.2.1群落相似性指数(Cj)
(1)
式中,a为样地A和B共有种数,b为样地B出现样地A没出现的种数,c为样地A出现样地B没出现的种数。Cj的值域为[0,1]。相似性等级(表2)一般划分为6级[25]:

表2 相似性指数(Cj)等级
2.2.2物种多样性指数选取Shannon-Wiener指数,其计算公式为
(2)
式中,H′为Shannon-Wiener指数;S为生境内物种数;Pi为物种i在群落中的相对重要值,其计算公式为:Pi=(相对多度+相对高度+相对盖度)/3[26]。
2.2.3物种Pielou均匀度指数(J)计算公式为
J=H′/lnS
(3)
2.2.4群落稳定性测定
采用Godron贡献定律法[27],利用群落中所有植物的种类和频度数据进行计算。以植物种类累积百分比为x轴,以累积相对频度为y轴,建立模糊散点平滑曲线,再求其与直线y=100-x的交点坐标,即为植物群落稳定点。交点坐标越接近(20,80),则群落稳定性越高[28]。平滑曲线模型为:
y=ax2+bx+c
(4)
2.3 数据处理与统计分析
采用EstimateS 9.0软件绘制各个样地植物累积丰度与样方数量的模拟曲线。应用PCORD 5软件中TWINSPAN分析对植物群落进行数量分类。利用Origin 2016软件绘制生活型组成、样方数量-植物累积丰度曲线和群落稳定性指数曲线。使用Rstudio软件对各样地的多样性指数进行多重比较分析以检验差异显著性大小,然后绘制箱型图,同时绘制优势种分布图及DCA排序图。
3 结果与分析
3.1 物种组成
3.1.1生活型组成
研究区7个样地共记录124种植物,分属于43科100属,其中以菊科植物最为丰富,有18属24种,其次是豆科(8属16种)、蔷薇科(12属15种),这3科植物种数占总物种数的44.35%。这124种植物中,木本植物31种,占植物总数的25.00%,其中乔木7种,占种总数的5.65%,灌木24种,占种总数的19.35%;草本植物93种,占种总数的75.00%。采用Raunkiner生活型系统来比较植物生活型组成(图2),发现火烧迹地恢复初期一年生植物只占植物总种数的12.10%,多年生草本植物占绝对优势,其中又以地面芽植物为主,占到植物总种数的42.74%。

图2 火烧迹地植物生活型组成
3.1.2优势种组成
根据实际调查情况,将每个样地中平均重要值大于8%的物种确定为优势种,共9种(图3),分别为大披针薹草(Carexlanceolata)、多花胡枝子(Lespedezafloribunda)、荠苨(Adenophoratrachelioides)、米口袋(Gueldenstaedtiaverna)、二色棘豆(Oxytropisbicolor)、地榆(Sanguisorbaofficinalis)、白莲蒿(Artemisiastechmanniana)、牻牛儿苗(Erodiumstephanianum)和茜草(Rubiacordifolia)。样地1和样地2包括4个优势种,大披针薹草和多花胡枝子在4个样地中出现,茜草、地榆、牻牛儿苗、白莲蒿则只在1个样地中出现。可见,火烧迹地恢复初期各样地优势种不尽相同,但大披针薹草和多花胡枝子为绝对优势种。

图3 样地-优势种关系
3.2 群落分类
3.2.1TWINSPAN聚类
参照《中国植被》的分类系统,根据火烧迹地恢复初期植物群落调查样方数据,选择在样方中出现频率≥95%的31种植物重要值组成的植物群落矩阵,进行TWINSPAN分类。依据植物群落分类和命名原则,结合调查结果分析的指示种和优势种命名群落类型,将火烧迹地恢复初期植物群落在3级划分水平上分为7类(图4)。

图4 TWINSPAN分类树状图
I. 大披针薹草+南牡蒿+地榆群丛
包括1、2、19—22、24、45、53、54、67等11个样方,主要分布在样地3,该群丛平均盖度约10%,均高约0.15 m,优势种为大披针薹草、南牡蒿、地榆,主要伴生种有小红菊(Chrysanthemumchanetii)、苍术(Atractylodeslancea)、二色棘豆等。
II. 多花胡枝子+大披针薹草+二色棘豆群丛
包括10—13、23、29、34、37—41、43、44、46—48、50—52、56等19个样方,主要分布在样地5和样地6,该群丛平均盖度约10%,均高约0.10 m。优势种为多花胡枝子、大披针薹草,伴生种主要有白莲蒿、华北前胡(Peucedanumharry-smithii)、野韭(Alliumramosum)等。
III. 多花胡枝子+大披针薹草+米口袋群丛
包括3—5、7—9、14、25—27、30—33、35、36等16个样方,主要分布于样地1和样地4,该群丛平均盖度约7%,均高约0.05 m。优势种为多花胡枝子和大披针薹草,伴生种主要有苍术、地榆、荠苨等。
IV. 多花胡枝子+大披针薹草+荠苨群丛
包括6、11、12、15—18、28、42等9个样方,主要分布于样地2,该群丛平均盖度约7%,均高约0.05 m。优势种为多花胡枝子、大披针薹草和荠苨,伴生种主要有茜草、地榆、斑叶堇菜(Violavariegata)等。
V. 大披针薹草+白莲蒿+狗娃花群丛
包括55、58、59、63等4个样方,主要分布于样地7,该群丛平均盖度约30%,均高约0.25 m。优势种为大披针薹草、白莲蒿、狗娃花,伴生种主要有桃叶鸦葱(Scorzonerasinensis)、斑叶堇菜、狗尾草(Setariaviridis)等。
VI. 白莲蒿群丛
包括60这1个样方,分布于样地7的坡中,该群丛平均盖度约50%,均高约0.40 m。优势种为白莲蒿,为单优种群落,伴生种主要有大披针薹草、华北前胡、野艾蒿等。
VII. 大披针薹草+狗娃花+野艾蒿群丛
包括61、62这2个样方,主要分布于样地7的坡下,该群丛平均盖度约20%,均高0.25 m。优势种为大披针薹草、狗娃花、野艾蒿,伴生种主要有多花胡枝子、委陵菜(Potentillachinensis)、香青(Anaphalissinica)等。
3.2.2DCA排序
采用DCA对研究区内7个样地的63个样方进行排序(图5),前4个排序轴的特征值分别是0.4167、0.3665、0.2869、0.2485。DCA排序的第一、二轴特征值较大,能较准确反映出植物群落相互之间及其与环境因子之间的关系。群落类型在DCA二维排序图上的分布一定程度上反映了群落所处空间的变化及其相互关系。从排序轴看,第一轴基本反映了群落类型随样地的变化,从左到右,群落所在样地在逐渐变化,最左边是分布在样地1和样地4中的多花胡枝子+大披针薹草+米口袋群丛,最右边则是分布在样地7中的白莲蒿群丛。第二轴基本反映了坡位的变化,即沿第二轴从下往上,坡位由坡下到坡上变化,植物群落也相应的改变。从排序图可以看出,样地的DCA二维排序图反映的植物群落结构与生境的空间位置关系,样地3和样地5的样方分布较集中,可见这两个样地的植物群落总体上相似性较大,群落类型较少。样地2与样地5的样方分布较分散,可见这两个样地的群落类型较为多样,群落间差别较大。样地1、样地2、样地4和样地5的各个样方间重合较大,表现出这些样地间的植物群落较相似,差别较小。总之,除样地7外,其余各样地的样方混合排布,可见火烧迹地恢复初期的植物群落大致可分为两大类,即样地7的植物群落和其它样地的植物群落。DCA排序结果与TWINSPAN的分类结果相匹配,可反映出火烧迹地恢复初期的植物群落结构较为单一。

图5 火烧迹地各样方DCA二维排序
3.3 群落相似性
为了解火烧迹地恢复初期植物群落的相似性,对所调查的各个样地进行群落相似性分析,计算了Jaccard相似性系数(表3)。由表3可以看出,各样地间的Jaccard群落相似性系数最高达0.39,最低为0.19,样地1与其余各样地极不相似。对照表1,可发现分布在极不相似水平和轻度相似水平的相似性系数约各占50%。可见,火烧迹地恢复初期各个样地群落间整体的相似性系数均比较低。
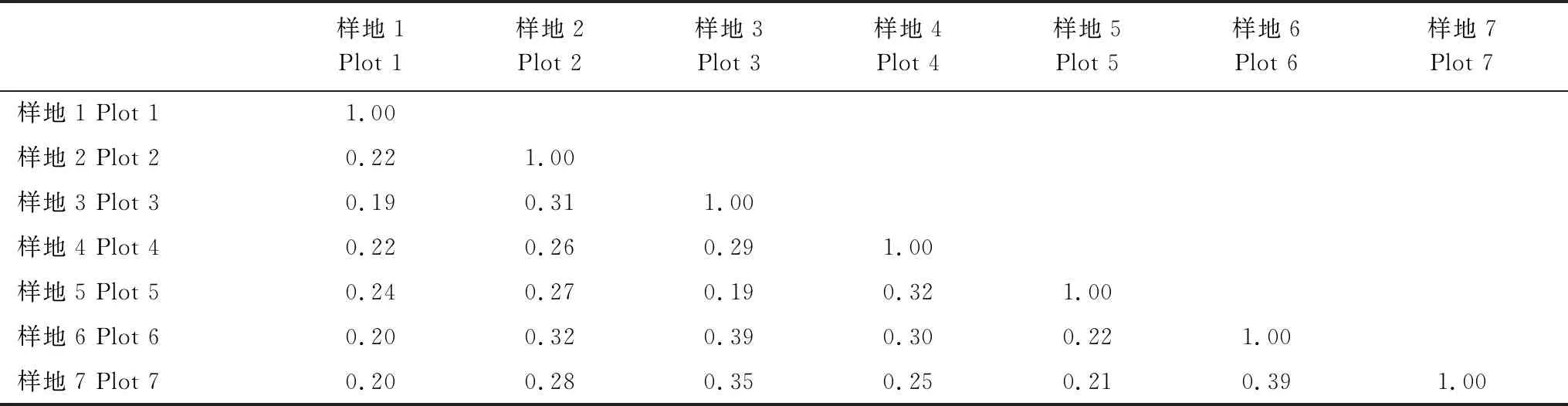
表3 不同样地植物相似性系数
3.4 物种多样性
3.4.1样方数量-物种丰度累积曲线
研究发现,在每个样地中随着样方数量的增加,植物物种数量在逐渐增加(图6)。各个样地所维持的物种数量均较小且存在差别,样地7的物种最丰富,样地5的植物种类最少,样地3和样地6的物种丰富度与变化趋势基本一致。可见在恢复初期,研究区内的植物物种丰富度还处在较低的水平。
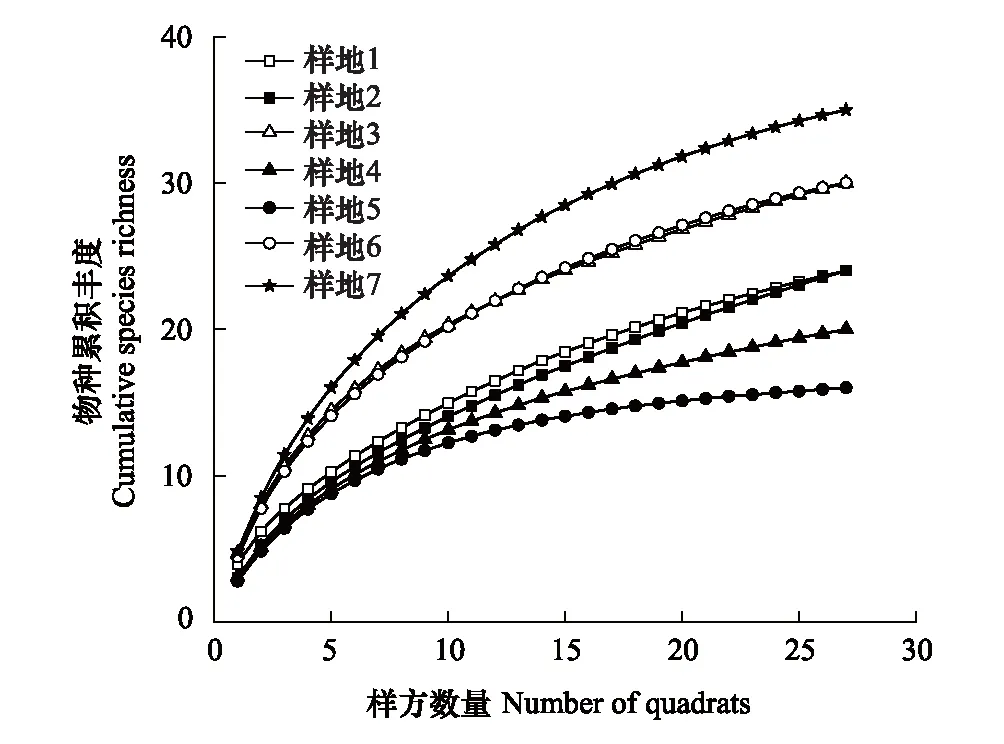
图6 样方数量-物种累积丰度关系
3.4.2多样性指数
对研究区各个样地的Shannon-Wiener指数、Pielou均匀度指数、物种丰富度和样方盖度进行分析,结果见图7。样地1、样地3、样地6和样地7的Shannon-Wiener指数相对较高,其余3个样地则相对较低,而且样地7的Shannon-Wiener指数均值最大;各个样地的Pielou均匀度指数总体上均较大,但存在差异。其中样地1、样地4、样地5和样地6相对较高,样地2和样地7的次之,样地3的均匀度指数平均值最小且显著低于除样地2外的其余各样地;各个样地的物种丰富度存在差异,样地7的物种丰富度相对较高,样地1、样地3和样地6次之,其余3个样地相对较低,且样地5的物种丰富度均值最小;各个样地的植物群落盖度总体上均较小,其中样地7的群落盖度均值最大且显著高于除样地6之外的其余各样地,样地1的群落盖度均值最小且显著低于除样地5外的其余各样地。综上可知,样地6和样地7植物群落各项指标均相对较高,可见这两个样地的植物群落恢复效果相对较好。

图7 不同样地植物群落多样性指数
3.5 群落稳定性
从图8可知,各个样地的累积相对频度-种类累积百分率曲线与直线的交点距离稳定点(20,80)都较远,稳定性大小排序为样地1>样地2>样地4>样地6>样地3>样地7>样地5,可见样地1的植物群落稳定性相对最高,样地5的稳定性最低。说明几个样地的群落均处在不稳定阶段,进而说明整个火烧迹地恢复初期植物群落处于不稳定的演替阶段。

图8 各样地植物群落稳定性
4 讨论
群落的结构特征是研究生态系统的动态变化规律及其生态功能形成与维持机制的基础。群落物种组成对群落动态变化规律、群落功能群划分以及植被恢复策略等方面的研究具有重要作用[29];TWINSPAN数量分类与DCA排序则可以客观地反映各植物群落间的相互关系;而群落相似性是群落结构分析的一个重要基础内容;同时群落多样性则可直观地反映出群落结构的变化;植物群落的稳定性一定程度上反映出各个种群间关联性,各种群间的变化关系又表现了植物群落的稳定程度,通过深入研究群落稳定性,可为植被保护与修复提供理论依据。所以本研究采用以上这些指标来进行探究火烧迹地恢复初期植物群落的结构及其维持机制。
4.1 群落物种组成特征
对太岳山油松林火烧迹地恢复初期的植物群落物种组成研究表明,研究区内菊科、豆科和蔷薇科3个科植物占绝对优势,另外40个科为少种和单种科,可见火烧迹地恢复初期植物一方面向大科集中,另一方面向少种科和单种科分散,与陈小雪等[30]的研究结果一致,这可能是由于火烧干扰形成多样化的生境为更多植物的生长提供了条件。童笑笑[31]对受水淹干扰的三峡水库消落带植物群落的研究也得出了类似的结果,可见这一现象可能是干扰条件下植物演替和适应的一种方向和方式;研究区内以多年生草本植物为主,其中又以地面芽植物所占比例最大;各个样地优势种不尽相同,但均为多年生植物,且均以大披针薹草和多花胡枝子为绝对优势种。李安定等[32]研究发现火烧迹地在自然恢复过程中,除种子萌发恢复外,其最主要方式取决于芽库数量。本研究无论从生活型组成还是从优势种组成均可看出,火烧迹地恢复初期以多年生植物为主,这些植物的地下繁殖体部分受火烧影响较小,在生长季仍然能够萌发生长,而一年生的先锋植物尚未大量入侵并定植,这可能是由于研究区周围都是油松林,一年生先锋植物种源较少,其次油松林的存在也可能在一定程度上阻碍了一年生植物种子的传播。
4.2 群落TWINSPAN数量分类与DCA排序
已有研究发现火烧迹地恢复初期草本层植物恢复较快[26,33],本研究用TWINSPAN分类法将火烧迹地恢复初期草本层植物群落分为7种群落类型。大披针薹草出现在其中6个群落当中,也可以显示出火烧迹地恢复初期植物群落优势种较为单一,各样地间有相似的植物群落类型。样地7的群落类型和其余6个样地差异相对较大,单独包含“V、VI、VII”3个群落类型。DCA排序的第一轴和第二轴分别基本反映了群落类型随样地与坡位的变化,其结果和TWINSPAN分类结果相互印证。不同学者对DCA排序轴的生态解释有不同的看法,可见DCA排序轴的生态解释会因为研究地点、研究对象或研究尺度的不同而存在差异[34]。TWINSPAN分类和DCA排序结果都表明火烧迹地恢复初期各样地植物群落较为相似,优势种较为单一,这一结果可能同样是由于原生植物群落结构所决定的,因为只经过一个生长季的恢复,大部分物种还未来得及侵入,仅保留了一部分未被火烧淘汰的原有多年生植物。而样地7与其它样地的植物群落存在差别,这可能与样地7的油松林较稀疏,受火烧强度较小,很多原生植物的繁殖体未被烧死等因素有关。
4.3 群落相似性
群落相似性是指群落间或样地间植物种类组成的相似程度,常用Jaccard相似性系数来表示[35]。总体而言,研究区各样地之间的Jaccard相似性系数比较低,在0.19—0.39之间。这可能因为各样地之间油松林下的原生植物群落在物种组成、群落盖度等方面不相同,加之火烧淘汰了部分以种子繁殖和扎根较浅的物种,进而影响了各样地之间的群落相似性。样地1与其余样地之间均表现为极不相似,这可能由于样地1的坡度陡、土层薄且砂石较多、林下原生植被较少等原因所导致。另外,群落相似性和样地间的距离也有一定的关系,距离近的样地间相似性系数较大,例如样地6和样地7。坡向、坡度、土壤等生境因子也会影响样地间的群落相似性,例如样地3和样地6虽然相隔较远,但因其生境相似,群落相似性系数也较大。群落相似性指数计算结果与TWINSPAN数量分类和DCA排序结果存在一定差异,这是因为TWINSPAN数量分类和DCA排序所用数据为在样方中出现频率≥95%的物种。但这也说明了研究区植物群落的优势种群相似性较高,但由于各个样地都有不同少见种和偶见种的存在,导致群落物种的整体相似性较低。
4.4 群落多样性
植物群落多样性可能会受到温度、水分、地形、土壤和干扰等诸多因素的影响。研究区7个样地在坡度、坡向、土壤和微地貌等方面都存在差异,可以说每个样地的微生境均不同。已有相关研究表明,生境异质性对植物群落多样性维持具有积极的作用[36]。如图7,群落Shannon-Wiener多样性指数和丰富度指数均大致可分为两部分,即样地1、样地3、样地6和样地7的Shannon-Wiener多样性指数较大,而其余3个样地则相对较小,这可能与各样地原生植被及样地间的生境条件密切相关。在调查时也观察到该4个样地中植物种类相对较多且植物长势较好,同时与样地1相比,其余3个样地的生境条件较为优越,预测在今后演替过程中样地1的植物群落恢复会相对较慢;各个样地的Pielou均匀度指数均比较高,说明火烧迹地恢复初期样地中各群落间的植物种类较为相似,这可能和各样地原生植物种质资源有很大关系,同时各群落中植物的丰富度与生长状况相近,所以均匀度较高;样地7的群落盖度显著高于其他样地,可能因为样地7原生油松林密度较低,林下植物群落盖度较大,加之一定的放牧干扰,在恢复初期各群落盖度就相对较大。总之,各个样地的群落多样性指数不尽相同,样地6和样地7植物群落各项指标均相对较高,但是演替初期的植物群落十分脆弱,高水平的物种多样性也并不能说明群落具备了稳定性[37],只能说明这2个样地的植物群落恢复相对较快,这可能和各个样地的火烧强度不同有很大关系。总体而言,火烧迹地恢复初期草本层植物群落除Pielou均匀度指数外,各个多样性指数均较低,这与邓贤兰等[38]的研究结果一致。
4.5 群落稳定性
已有研究表明种间竞争、生境条件和人为干扰等因素均会影响植物群落稳定性[39]。本研究采用Godron稳定性分析方法,发现各个样地群落稳定比值远离(20,80),可见火烧迹地恢复初期植物群落各物种间尚未形成稳定共存的组合及适宜的种间关系,群落的结构还不稳定,物种组成及其数量结构处于动态变化中。曹慧等[24]研究也发现油松林火烧后物种多样性和群落的稳定性均长时间处于较低的状态。这可能是因为研究区油松林的群落结构简单,成层性较差,种源缺乏,恢复速度相对较慢。本研究还发现各个样地间群落稳定性存在差异,说明不同样地间微生境的不同是影响火烧迹地植物群落稳定程度的因素之一。不同样地的光照、坡度、土壤等生境条件以及人为干扰程度不同,导致各样地群落稳定性系数不同。样地1植物群落的稳定性系数最大,但其植物种类与多样性却较低,这可能是因为样地1的坡度较大,微地貌较为单一,植物群落类型也较少。样地5的植物群落稳定性系数最小,可能是因为样地5内存在沟谷、平地、坡地以及虎榛子灌丛等多种微生境,导致群落类型较为多样,物种间关系较复杂,群落稳定性较低。
4.6 对植被修复的建议
太岳山区纯林较多,林分结构简单,易燃树种比例大、分布广,特别是油松林分布广泛,而且林下松针累积较厚,很容易发生森林火灾[24]。在野外调查过程中观察到一些植物,如虎榛子、灰栒子(Cotoneasteracutifolius)和黄刺玫等灌木以及蒙古栎只是地上部分被烧毁,又从茎干基部萌发出了新枝。预测在接下来几年的演替过程当中,研究区的植物群落会从多年生草本植物群落到灌木和草本植物群落逐渐到蒙古栎群落演替。虽然蒙古栎和油松均为太岳山区现有森林群落的主要树种,但实际上该地区属于黄土丘陵区,水分条件和土壤条件均比较差,并不适宜种植高大乔木,加之乔木的生长周期较长,建议在该地区火烧迹地修复过程中不宜只飞播油松种子,而是要因地制宜,科学地进行植物配置。根据火烧迹地的火烧强度、地形地势、土壤和光照等条件,选择合适的物种搭配,以提高更新速度并减少成本。建议在坡度较陡、土层较薄的阳坡,主要栽植胡枝子、沙棘和黄刺玫等耐旱灌木以及大披针薹草等耐旱草本,在坡度较缓的阳坡可栽种蒙古栎,同时可以在阴坡合理种植油松和虎榛子,这样可以加快植被恢复进程,又可减少火烧迹地水土流失,并兼顾植物多样性保育。
