Mechlppdk基因在木薯中的表达分析及RNA干扰载体构建和遗传转化
2021-07-09王海燕周新成王文泉
王海燕,陈 新,周新成,沈 旭,孔 华,王文泉
(1中国热带农业科学院热带生物技术研究所,海口 571101;2南京农业大学生命科学院,南京 210095;3海南大学热带作物学院,海口 570228)
0 引言
木薯是具有高生物量的重要热带植物,起源于南美洲,是全球三大块根作物之一,第六大粮食作物和新兴能源作物,具有高效累积光合产物和抗旱、耐瘠薄等特性[1],广泛种植于全球热带地区和中国两广,海南地区。栽培木薯是典型的C3-C4型热带植物,具有高光效高淀粉累积的特点。木薯块根中积累贮藏的同化物仅仅是地上合成同化物的1/5,所以木薯叶片具有超强的光合碳固定能力。但是C4型碳固定对木薯叶片的这种超强光合碳固定的作用效率尚未见报道。
栽培木薯叶片PEPC酶的活性从8.3~80 mmol/(kg(Chl)·s),约是玉米和高粱的 15%~25%[2]。栽培木薯PEPCase活性远远高于田间大豆等C3植物的活性,与C3-C4中间型黄花菊属几个种的活性相当,比包含类似花环结构、C3-C4型黍属的Panicum milioides中的活性高2~3倍[3],C4PEPC的存在通过免疫学方法得到进一步确证。利用来自玉米的PEPC、苹果酸酶和苹果酸脱氢酶的探针均能在栽培木薯叶片中检测到苹果酸酶和苹果酸脱氢酶的存在。当前对栽培木薯光合作用属于C3-C4中间型的认识仅限于栽培木薯叶片中有C4酶的存在,但是其中C4光合碳同化的效率,及对木薯生长发育的影响迄今尚未有报道。
丙酮酸磷酸双激酶(PPDK)是C4光合作用的关键酶,催化C4光合作用原初CO2固定的受体磷酸烯醇式丙酮酸(PEP)的再生,继而由磷酸烯醇式丙酮酸羧化酶(PEPC)催化PEP接受二氧化碳,合成草酰乙酸(OAA),至此完成二氧化碳的固定。玉米中C4循环是由叶肉细胞和维管束鞘细胞协作完成。同时发现玉米PPDK是C4循环的限速酶,一旦PPDK的活性被干扰,C4途径就会被影响[4]。PPDK首先是在C4植物中发现[5],后面相继在C3植物,原生动物和细菌都有发现。PPDK在C3和C4植物中是以同型四聚体形式存在,在细菌和原生动物是以二聚体形式存在[6],但他们的初级结构非常相似,表明ppdk基因在原核生物和真核生物分化前就存在了。迄今,科学家们已在多种植物中克隆到 ppdk基因,如小麦[7]、玉米[8]、水稻[9]、拟南芥[10]、甘蔗[11]、高粱[12]、籽粒苋[13]、家稗[14]等。
RNA干扰(RNA interference,RNAi)是指细胞中导入与内源性mRNA编码区某段序列同源的双链RNA(double-stranded RNA,dsRNA),可致该mRNA发生特异性降解从而导致基因表达沉默的现象[15]。该技术能够干扰特定基因,高效、特异地阻断体内基因表达,诱使细胞表现出特定基因缺失的表型,具有高效性、扩大性、特异性及可遗传性等特点。经过多年实践,RNA干扰技术日趋成熟,现已广泛应用于植物基因功能研究[16-17]。本研究拟利用QPCR确定Mechlppdk在木薯不同组织器官的表达谱,构建木薯Mechlppdk的RNA干扰载体,并转化木薯,获得Mechlppdk的干扰突变体,为下一步木薯丙酮酸磷酸双激酶基因Mechlppdk功能的研究提供研究材料。
1 材料与方法
1.1 实验材料和试剂
ppdk基因表达谱分析木薯材料叶片,茎秆,块根均采自生物所试验地田间种植180天木薯植株(Mahnihot esculenta var.Arg7)。木薯遗传转化用材料是模式木薯TMS60444。大肠杆菌(Escherichia coli)DH5α系本实验室留存,农杆菌(Agrobacterium tumefaciens)LBA4404是上海生命科学研究院张鹏研究员馈赠,RNA干扰中间载体pHANNIBAL和pART27表达载体是本研究所郭安平课题组馈赠;Taq DNA polymerase、pMD18-T、T4 DNA连接酶购自大连宝生物公司,DNA凝胶回收试剂盒购自OMEGA公司,其他生化试剂为进口或国产分析纯试剂。引物合成和DNA测序是由生工生物工程(上海)有限公司完成。
1.2 方法
1.2.1 木薯叶片RNA提取和cDNA的合成 参照李正绪方法进行[18]。
1.2.2 Mechlppdk QPCR引物设计 通过比对GeneBank上公布的木薯Mechlppdk的cDNA序列,在转运肽区设计定量引物,如下QF:5’-GCAGGGGTATAGAGGAT CGC-3’;QR:5’-GGCGCCGTCCTAATTAACAT-3’,木薯的β-actin基因作为管家基因,actinF:5’-CAAGGGCAACATATGCAAGC-3’,actinR:5’-CCTT CGTCTGGACCTTGCTG-3’。热循环仪是Rotor-gene 6000,扩增程序如下:95℃预变性 1 min;95℃ 10 s,59℃ 15 s,72℃ 20 s,40个循环。每个样品设置3个重复,取其平均值用于表达分析。采用2-ΔΔCT进行数据分析,并使用IBM SPSS Statistics 19.0进行差异显著性分析。
1.2.3 Mechlppdk RNAi的引物设计 通过找到Mechlppdk的特异区段,分别设计2对引物,即在引物序列前分别加两个不同的酶切位点,形成带有不同酶切位点的两个反向互补片段,具体引物序列如下:senseF:5’-GAATTCATGTCGTCGGCG-3’;senseR:5’-AGGAGCAAGAGACTCGAG-3’;antisenseF:5’-ATC GATTCTCTTGCTCCT-3’;antisenseR:5’-CGCCG ACGACATTCTAGA-3’。同时把中间载体的内含子序列加到反向互补片段之间,预测其二级结构。
1.2.4 木薯Mechlppdk RNAi片段的PCR扩增及RNA干扰中间载体的构建 以1.2.2中合成的cDNA为模板,利用上述设计的引物,扩增Mechlppdk的特异片段,电泳,切胶、回收后连接PMD18-T载体,转化感受态细胞(E.coli DH5α)后,提取质粒并测序。根据测序结果选取与所设计干扰片段序列一致的两个质粒分别命名为p Mechlppdki-XE和p Mechlppdki-CX,分别用Xho I/EcoR I和Cla I/Xba I进行双酶切,电泳,切胶回收目的片段分别命名为Mechlppdki-XE和Mechlppdki-CX。同时用Xho I/EcoR I双酶切中间载体p HANNIBAL,胶回收大片段后用T4DNA连接酶将其与回收的Mechlppdki-EX连接,转化感受态细胞(E.coli DH5α)后用氨苄(终浓度为50 μg/L)筛选阳性克隆,提取质粒并酶切检测Mechlppdki-XE是否与pHANNIBAL载体连接,连接上Mechlppdki-XE的pHANNIBAL质粒命名为pHANNIBAL-MechlppdkiXE,将pHANNIBALMechlppdkiXE进行Cla I/Xba I的双酶切,切胶,回收目的片段后用T4DNA连接酶将其与回收的Mechlppdki-CX连接,转化感受态细胞(E.coli DH5α)后用氨苄(终浓度为50 μg/L)筛选阳性克隆,提取质粒并酶切鉴定,获得阳性克隆,命名为pHANNIBAL-MechlppdkiXECX。
1.2.5 RNA干扰表达载体的构建 根据实验室干扰载体pART27的质粒图谱,使用Not I对pHANNIBALMechlppdkiXE-CX和pART27分别进行Not I酶切,电泳,切胶回收,将回收后的MechlppdkiXE-CX大片段和pART27的大片段进行连接,转化感受态细胞(E.coli DH5α)后用卡那霉素(终浓度为50 μg/L)筛选阳性克隆Mechlppdk RNAi表达载体,具体流程详见图1。
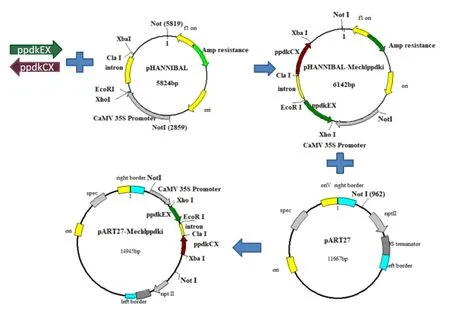
图1 Mechlppdk RNAi干扰载体构建流程图
1.2.6 RNA干扰载体转化木薯及nptII的PCR验证及QPCR检测 将1.2.4中获得的干扰载体通过电击转化的方法转化农杆菌LBA4404,获得含有干扰载体的农杆菌菌株,然后侵染木薯TMS60444脆性胚愈伤,通过共培养,洗菌,诱导,最后获得具有卡那霉素抗性的转基因木薯苗。具体方案参照文献[19-20]。将生根筛选出的转基因木薯苗提取基因组DNA,通过PCR的方法进行nptII的鉴定。利用Mechlppdk QPCR引物对获得的转基因株系进行Mechlppdk的表达分析。
2 结果与分析
2.1 Mechlppdk在木薯不同组织器官的表达分析
如图2,通过对180天的木薯根、茎和叶3个器官中Mechlppdk基因的表达分析,发现Mechlppdk在叶片中表达量最高,达到内参基因的50%,茎中只有可见的少量表达,根中没有表达。说明Mechlppdk主要在叶片中起作用。
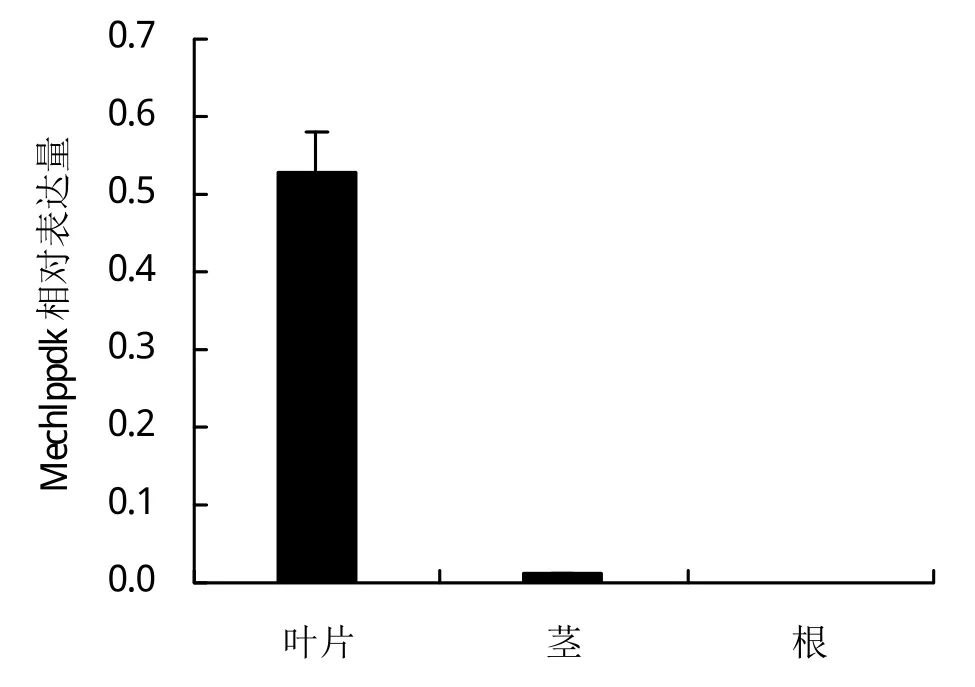
图2 Mechlppdk在根、茎、叶中的相对表达量
2.2 Mechlppdk干扰载体特异序列的选择
将JGI网站公布的Mechlppdk(Manes.03G188300),经BLAST,选择转运肽区一段保守序列,自第1个碱基至165个碱基,共165个碱基。具体序列信息如图3A。将正反序列中间连接pHANNIBAL的内含子,预测二级结构,如图3B,形成典型的发夹结构。

图3 木薯Mechlppdk siRNA设计
2.3 Mechlppdk RNA干扰片段的克隆
通过RT-PCR扩增得到175 bp左右的目的片段(包含酶切位点序列),见图4A,将目的片段连接至pMD-18T克隆载体上,通过化学转化法转至大肠杆菌DH5α中,经氨苄筛选获得阳性菌落,通过PCR鉴定,结果显示得到的小片段与预期大小一致(图4 B)。将阳性克隆送测序,选择与Mechlppdk选定序列完全一致的克隆,提取质粒,分别命名为pMechlppdkiXE和pMechlppdkiCX,用于下步构建。
2.4 Mechlppdk RNA干扰中间载体的构建
首先用EcoR I/Xho I分别酶切pMechlppdkiXE和pHANNIBAL,电泳,分别胶回收小,大片段,并用T4DNA连接酶将目的片段和载体连接,转化大肠杆菌DH5α,在带有氨苄的培养基上筛选阳性克隆,提取质粒酶切鉴定,结果见图5A,经EcoR I/Xho I双酶切后,获得与图4中PCR扩增大小一致的条带,将该阳性质粒命名为pHANNIBAL-MechlPPDKiXE。继而用Cla I/XbaI双酶切pHANNIBAL-MechlppdkiXE和pMechlppdkiCX,电泳,胶回收目的片段,用T4DNA连接酶连接载体和小片段,转化大肠杆菌DH5α,在筛选培养基上筛选阳性克隆,PCR扩增正确的克隆进行质粒提取和酶切鉴定,结果见图5B,经Cla I/Xba I双酶切,在100 bp和250 bp的条带之间有一条约170 bp的条带,与图4中PCR扩增的条带大小一致,酶切正确的阳性质粒命名为pHANNIBAL-MechlppdkiXE-CX.
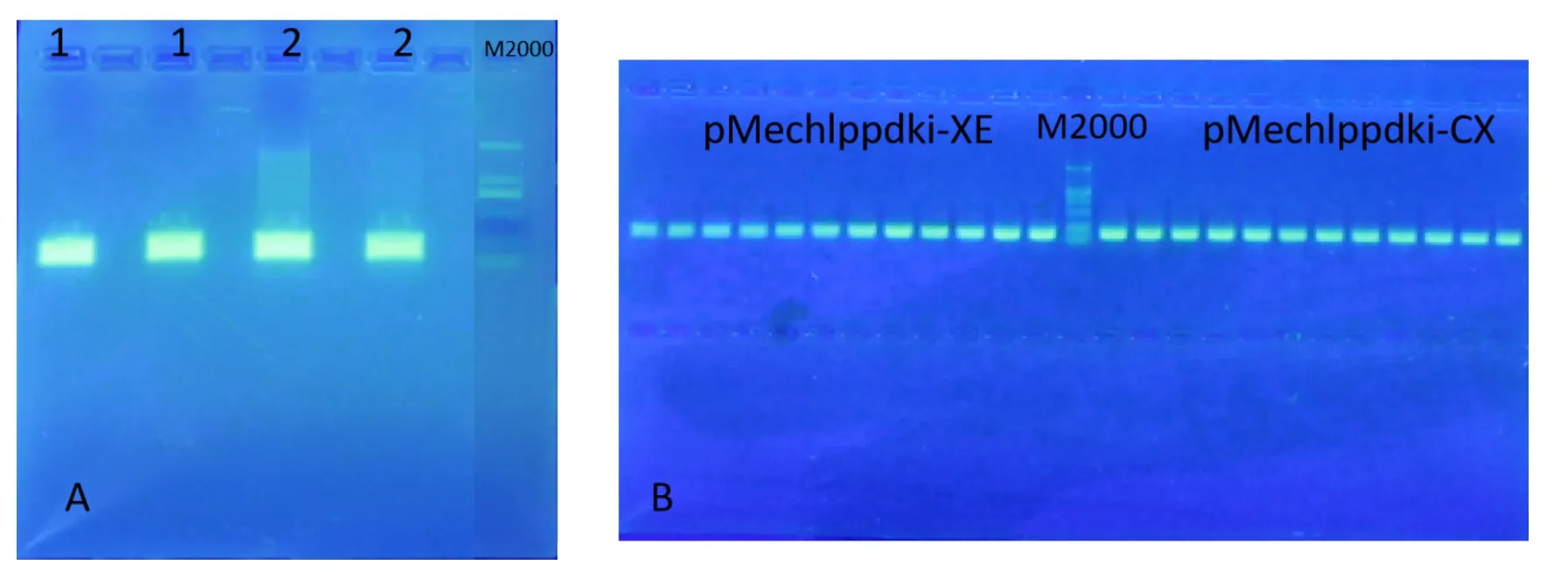
图4 Mechlppdk RNA干扰片段的扩增及克隆鉴定
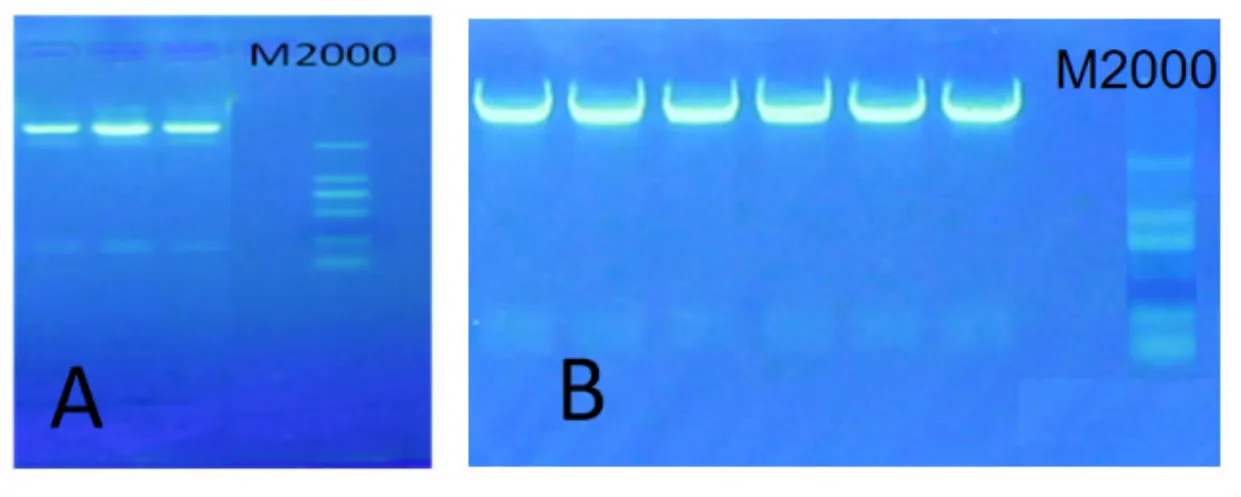
图5 pHANNIBAL-MechlppdkiXE-CX中间载体酶切鉴定
2.5 Mechlppdk RNA干扰表达载体的构建并转化农杆菌LBA4404
用Not I分别酶切pART27和pHANNIBALMechlppdkiXE-CX,电泳,胶回收目的片段和线性载体,用T4DNA连接酶将载体和目的片段连接,转化大肠杆菌DH5α,在带有卡那霉素的LB培养基上筛选阳性克隆,经菌液PCR(图6A),将菌液PCR阳性的克隆进一步提取质粒,Not I酶切,有8个克隆可切出与预期大小一致条带,结果见图6B,此时即获得pART27-MechlppdkRNAi。将 pART27-MechlppdkRNAi转化农杆菌LBA4404的感受态细胞,筛选培养基上获得阳性克隆,并经PCR和质粒酶切检测为阳性。
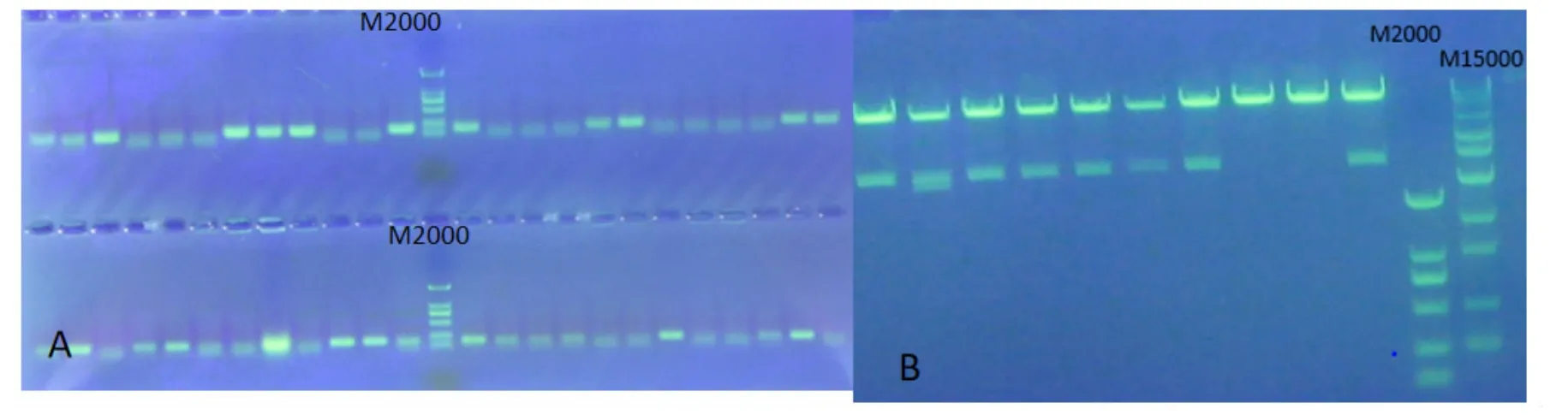
图6 pART27-MechlppdkRNAi的分子鉴定
2.6 pART27-MechlppdkRNAi转基因木薯苗的获得
将获得的带有干扰载体的LBA4404侵染木薯品种TMS60444的脆性胚愈伤,经过诱导,筛选,获得具有卡那霉素抗性苗。转基因结果如图7。抗性苗放置在添加有50 μg/L卡那霉素的MS培养基上进行生根筛选,如图7D,7~9天部分转基因植株在抗性培养基上生根,野生型没有生根。继而选择生根的株系进行卡那霉素Npt II的PCR鉴定,成功扩增到目的条带,如图7E。最终获得7个Mechlppdk的干扰株系。

图7 转pART27-MechlppdkRNAi木薯苗的诱导及鉴定
对上述获得的7个转基因木薯株系考察Mechlppdk基因的表达情况,结果如图8。7个株系中有4个株系(1,2,3,7)的Mechlppdk的表达量大幅下调,其中最大的有7号株系,Mechlppdk的表达量下调到野生型的9.7%。有2个株系的Mechlppdk的表达量有轻微下调,表达量分别下调到野生型的87%和74%。
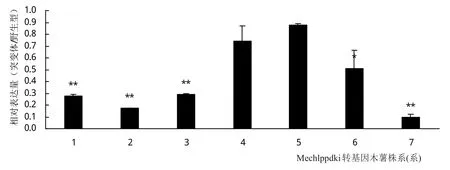
图8 Mechlppdk干扰株系Mechlppdk的相对表达分析
3 讨论与结论
RNAi是由双链RNA分子在细胞内干扰同源基因表达下调或沉默的技术[21]。RNA干扰最早追溯到1990年,Napoli等[22]在将查耳酮合酶基因(chs)转入牵牛花中意外发现的。RNAi能够定向关闭生物体内的某一基因,使其不发挥作用,同时不影响其他基因,方便人们对特定基因功能的研究[23]。当前广为应用的还有crisp技术,是在DNA水平敲除目的基因,效率高,但对于重要的功能基因,完全敲除后,往往具有植株致死性。而RNAi技术可以不同程度的干扰目的基因的表达,对于研究重要功能基因具有优势。但是RNAi的特异性是相对的,在某些情况下,siRNA能在翻译水平抑制与之不完全相关的基因表达即脱靶效应。为了应对此类脱靶现象,应该针对同一基因多设计几个靶点,以确保靶基因表达的有效抑制[24]。本研究共获得7个Mehlppdk的干扰株系,其中3个株系Mechlppdk的表达量显著下调,说明本次对Mechlppdk干扰操作有效。
双子叶植物中,ppdk由单基因编码[25],5’有两个不同的启动子分别产生细胞质和质体定位的转录本[26]。木薯ppdk同样有2个启动子,分别产生叶绿体定位的Mechlppdk和细胞质定位的Mecyppdk。前期研究发现Mecyppdk完全嵌套在Mechlppdk里面,所不同的是Mechlppdk的5’多一转运肽序列,具有组织特异性,主要在叶绿体表达[27]。本研究为更准确的干扰Mechlppdk基因,选择了该基因的5’转运肽区进行干扰载体的构建,最大限度的减少对Mecyppdk基因的干扰。目前尚未看到关于木薯Mechlppdk基因功能的研究报道。Zhang等[28]获得了chlppdk功能缺失的玉米突变体,该材料的叶片在种植108 h后出现白化,且消耗完种子内存活的营养物质就会死亡。表明chlPPDK对于玉米的生存至关重要。接下来将利用获得的干扰突变体展开生理生化和分子生物学研究,以期阐明chlPPDK对木薯生长发育的影响。
本研究通过QPCR发现Mechlppdk在木薯叶片中具有较高的表达,达到内参基因的50%;通过构建木薯Mechlppdk的干扰载体转化木薯,获得7个阳性转基因株系,其中3个株系较大幅度的降低了Mechlppdk的表达,最低的仅是本底chlppdk的9%。这些转基因干扰株系将为Mechlppdk功能研究提供非常有利的实验材料,同时能够部分阐述光合C4固定对木薯生长发育的影响。
