典型草原啮齿动物群落对不同放牧干扰的响应
2021-07-09郭乾伟苏佳楼崔志刚付和平
马 久,袁 帅,郭乾伟,苏佳楼,崔志刚,金 国,张 恒,付和平,*
(1.内蒙古乌兰察布市察右后旗土牧尔台镇政府,土牧尔台 012406;2.内蒙古农业大学草原与资源环境学院/内蒙古农业大学啮齿动物研究中心/内蒙古农业大学草地资源教育部重点实验室/农业农村部饲草栽培、加工与高效利用重点实验室,呼和浩特 010018;3.内蒙古锡林郭勒盟草原工作站, 锡林浩特 013000;4.内蒙古锡林郭勒盟东乌珠穆沁旗草原工作站,乌里雅斯太 026300;5.内蒙古锡林郭勒盟正镶白旗草原工作站, 明安图 013800)
草地啮齿动物群落是群落生态学研究中不可或缺的重要内容,由于其与草地生态系统的密切互作关系,一直备受关注[1~2]。草地啮齿动物群落结构是指啮齿动物在草地环境中的分布及其与环境之间长期的相互作用所形成的结构, 主要包括群落的时间结构和空间结构。群落的时间结构和空间结构变化是群落演替动态特征的主要表现形式[3],是由自然环境因素的节律性改变所引起的啮齿动物群落在时间和空间结构上相应的周期性或非周期性变化[3~5]。而人为干扰活动,如放牧、休牧、禁牧等可直接或间接地改变啮齿动物群落成分间的相互关系,导致自然生境的相应变化, 从而对动植物群落之间的关系和系统功能产生重大影响[5~6]。人为干扰导致自然栖息地破碎化过程中生物群落格局的变化已有一些报道[2], 但主要集中在半荒漠及荒漠生境中的研究,关于典型草原区啮齿动物群落特征与放牧干扰关系的研究相对较少[7~9],而有关啮齿动物群落组成、结构及优势种动态的研究更是少见[10]。典型草原是我国北部典型的干旱生态系统, 生物群落结构并不复杂, 分布其中的啮齿动物占有重要地位,在典型草原生态系统中发挥着重要作用。因此,了解啮齿动物群落组成、结构及其优势种群在不同放牧干扰下的变动特征,是再认识典型草原生态系统动植物关系及系统功能干扰效应的重要内容, 也是探讨典型草原啮齿动物群落干扰生态学的前提,有助于深入了解典型草原生态系统的结构、功能以及草地退化的生态机制, 同时对典型草原生态系统的恢复与重建及鼠害的综合防控都具有重要意义。本研究于2014~2018年在锡林郭勒大针茅典型草原区设置过度放牧(OG)、季节轮牧(QG)和禁牧(GE,对照)3个处理区,对不同草地利用方式人为干扰下啮齿动物群落的演替和优势种群动态进行研究,以期探索由自然环境因素的节律性改变所引起的啮齿动物群落在时间结构上相应的变动特征,明确典型草原生态系统动植物关系及系统功能的干扰效应,为典型草原区鼠害防控提供基础依据。
1 研究区自然条件与研究方法
1.1 自然条件
研究地位于内蒙古锡林郭勒盟东乌珠穆沁旗境内典型草原区,地理坐标为44°42'~45°15'N、 115°30'~116°30'E,草地类型为典型草原,主要植被种类为大针茅(Stipagrandis)、羊草(Leymuschinensis)、糙隐子草(Cleistogenessquarrosa)、多根葱(Alliumpolyrhizum)、栉叶蒿(Neopallasiapectinata)等;气候属于典型的大陆性气候,冬季寒冷风大,夏季水热同期,年平均气温1.6℃,极端最高气温39.7℃,最低气温-40.7℃,无霜期120d。年降水量300mm左右,主要集中在6~8月份。
1.2 研究方法
2014~2018年,在锡林郭勒大针茅草原区采用完全随机区组试验设计,将9个小区划分为3个区组,每个区组中设有3个处理,即禁牧区(GE,对照)、季节轮牧区(QG)和过度放牧区(OG)。每个小区面积分别为10hm2。禁牧作为对照不进行放牧;季节轮牧分别将样地划分为2个等面积小样区,在小样区间进行周期为2个月的轮流放牧;过度放牧为超过当地载畜量的自由放牧。每个样区设置3个啮齿动物调查样方,分别进行同心圆布笼取样,布笼方式如图1。鼠类调查采用标志重捕法,调查期间以新鲜花生米为诱饵,每年5月、7月和9月分别进行一次野外调查,每次连续4d,每日早、午、傍晚共查笼3次,并记录捕获鼠类的位置、性别、体重(g)及繁殖状况,使用PIT芯片对捕获的鼠类进行标记。
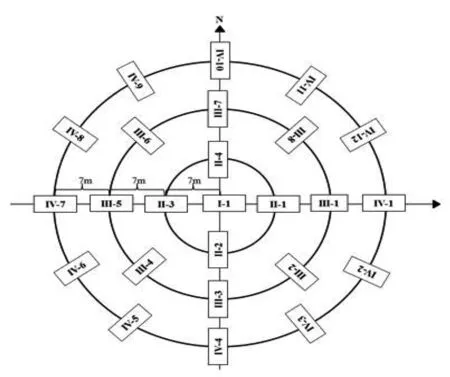
同心圆的半径由内向外分别为7m、14m和21m,每一个方框代表1个布笼位置,方框中的号码代表每一个笼子唯一的编号图1 同心圆布笼方式
2 数据处理
本文采用百笼捕获率表示啮齿动物种群密度,即以100笼所捕获的不重复啮齿动物数量来计算,公式为:
式中:P为捕获率;N为所捕获鼠的数量;H为总的布笼数;n为连续布笼天数。
采用啮齿动物捕获比例确定群落中优势种和次优势种,计算公式如下:
式中:B为捕获比例;A为某种鼠所捕获个体的数量;M为总的鼠的捕获数量;
本文啮齿动物群落中种群数量差异性分析采用SPSS12.0单因素方差分析(One-way ANOVA),显著性水平P<0.05。
3 结果与分析
3.1 不同放牧方式下啮齿动物群落组成
研究期间在3个处理样地共布放活捕笼13500笼日,捕获啮齿动物275只,隶属4科7种:松鼠科(Sciuridae)的草原黄鼠(Spermophilusdauricus);仓鼠科(Circetidae)的布氏田鼠(Lasiopodomysbrandtii)、黑线毛足鼠(Phodopussungorus)、黑线仓鼠(Cricetulusbarabensis)和长爪沙鼠(Merionesunguiculatus);跳鼠科(Dipodidae)的五趾跳鼠(Allactagasibirica);鼠兔科(Ochotonidae)的达乌尔鼠兔(Ochotonadaurica)。在3个处理样地,总体上布氏田鼠捕获比例均为最高,是啮齿动物群落的优势种。过度放牧区布氏田鼠捕获量比例为33.33%,其次为黑线仓鼠25.36%,黑线毛足鼠为24.18%,其余种类均小于10%,啮齿动物群落为布氏田鼠+黑线仓鼠+黑线毛足鼠群落;季节轮牧区布氏田鼠捕获比例51.20%,其次为黑线毛足鼠23.20%,黑线仓鼠为22.82%,其余种类均小于10%,啮齿动物群落为布氏田鼠+黑线毛足鼠+黑线仓鼠群落;禁牧区布氏田鼠捕获比例为64.67%,黑线仓鼠为25.30%,其余种类捕获比例均小于10%,啮齿动物群落为布氏田鼠+黑线仓鼠群落。
3.2 啮齿动物群落对不同放牧干扰的响应
3.2.1对过度放牧干扰的响应
过度放牧区啮齿动物群落组成为布氏田鼠、黑线毛足鼠、黑线仓鼠和草原黄鼠共4种,其种群数量的年际波动如图2,达乌尔鼠兔、五趾跳鼠和长爪沙鼠在过度放牧区5个年度中均未捕获到。从啮齿动物群落组成的年度变化看,2014年黑线仓鼠种群密度最大,捕获率为2.20%,其次为黑线毛足鼠1.60%,啮齿动物群落为黑线仓鼠+黑线毛足鼠群落。2015年黑线仓鼠种群密度仍为最大,捕获率为2.10%;其次为草原黄鼠,捕获率为0.20%,啮齿动物群落为黑线仓鼠+草原黄鼠群落。2016年草原黄鼠种群密度最高,捕获率为7.80%,种群数量显著增加;其次为黑线毛足鼠,捕获率5.60%,黑线仓鼠捕获率为2.10%,啮齿动物群落为草原黄鼠+黑线毛足鼠+黑线仓鼠群落,群落组成和结构较2014和2015年度发生了明显改变。2017年草原黄鼠种群密度显著下降,捕获率为1.10%,黑线毛足鼠种群密度最高,捕获率4.00%,黑线仓鼠捕获率与草原黄鼠相同(1.10%),因此啮齿动物群落为黑线毛足鼠+草原黄鼠+黑线仓鼠群落,群落结构再一次出现改变。2018年布氏田鼠数量显著增加,捕获率上升到15.60%,与其他3个种群的密度差异显著(P<0.01),成为该啮齿动物群落的优势种;其次为黑线仓鼠,捕获率为2.20%,黑线毛足鼠捕获率为1.50%,啮齿动物群落为布氏田鼠+黑线仓鼠+黑线毛足鼠群落,群落组成和结构又一次发生改变。从上述结果可以看出,随着草地的过度放牧利用,啮齿动物群落组成和结构发生了明显的演替,群落优势种由最初的黑线仓鼠逐渐被草原黄鼠、黑线毛足鼠和布氏田鼠所替代。
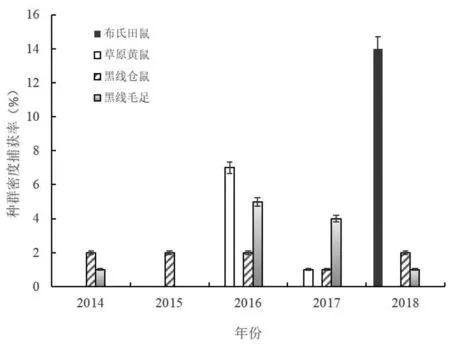
图2 过度放牧干扰下鼠类种群密度年度动态
3.2.2对季节轮牧干扰的响应
季节轮牧区啮齿动物群落组成为布氏田鼠、黑线毛足鼠、黑线仓鼠、草原黄鼠、达乌尔鼠兔和五趾跳鼠共6种,其种群数量的年际波动如图3,长爪沙鼠在季节轮牧区5个年度中均未捕获到。从啮齿动物群落组成的年度变化看,2014年黑线毛足鼠种群数量最高,捕获率为8.90%;其次为达乌尔鼠兔,捕获率2.10%; 黑线仓鼠捕获率为1.60%;啮齿动物群落为黑线毛足鼠+达乌尔鼠兔+黑线仓鼠群落。2015年黑线仓鼠种群密度最大,捕获率6.70%;其次为黑线毛足鼠,捕获率为1.20%;啮齿动物群落为黑线仓鼠+黑线毛足鼠群落。2016年黑线仓鼠种群密度仍为最高,捕获率5.60%;其次为黑线毛足鼠,捕获率为4.80%;草原黄鼠捕获率为4.40%;啮齿动物群落为黑线仓鼠+黑线毛足鼠+草原黄鼠群落,群落组成和结构发生了明显改变。2017年草原黄鼠种群密度继续上升,捕获率5.20%;其次为黑线毛足鼠和黑线仓鼠,捕获率均为4.80%;啮齿动物群落为草原黄鼠+黑线毛足鼠+黑线仓鼠群落,群落结构发生改变。2018年布氏田鼠种群数量极显著增加,捕获率达到41.60%,占季节轮牧干扰区啮齿动物群落中个体数量的90%以上,与其他5个种群的密度呈极显著差异(P<0.001),成为绝对的优势鼠种;其次为五趾跳鼠和草原黄鼠,捕获率均为3.60%;啮齿动物群落为布氏田鼠+草原黄鼠+五趾跳鼠群落。上述结果表明,草地季节轮牧利用方式也使得啮齿动物群落组成和结构发生了明显演替,群落优势种由最初的黑线毛足鼠逐渐被黑线仓鼠、草原黄鼠和布氏田鼠所替代。
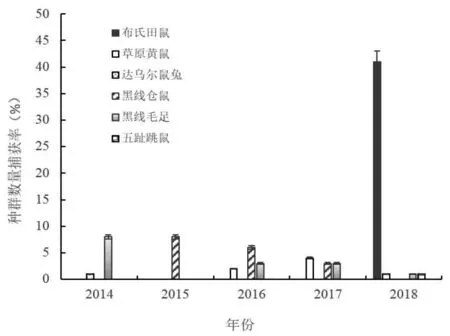
图3 季节轮牧干扰下鼠类种群密度年度动态
3.2.3对禁牧干扰的响应
禁牧区是本研究设置的对照区,啮齿动物群落组成为布氏田鼠、黑线毛足鼠、黑线仓鼠、达乌尔鼠兔和草原黄鼠共5种,其种群数量的年际波动如图4,长爪沙鼠在5个年度中均未捕获到。从啮齿动物群落组成的年度变化看,2014年只捕获到了黑线毛足鼠和黑线仓鼠,黑线毛足鼠捕获率虽然只有10%左右,但是占啮齿动物群落中个体总数的80%,是禁牧干扰区的绝对优势鼠种,因此群落为黑线毛足鼠+黑线仓鼠群落;由于黑线毛足鼠种群数量波动较大,2015年和2018年在禁牧区均未捕获到。2015年黑线仓鼠种群数量明显增加,捕获率近20%,个体数占整个群落的80%以上,是明显的优势种;其次为达乌尔鼠兔,捕获率为4.50%;啮齿动物群落为黑线仓鼠+达乌尔鼠兔群落。2016年黑线仓鼠种群数量仍然最高,捕获率15.60%,占啮齿动物群落中个体总数的80%以上,为明显的优势鼠种;其次为黑线毛足鼠,捕获率为6.80%;草原黄鼠捕获率为4.00%;啮齿动物群落为黑线仓鼠+黑线毛足鼠+草原黄鼠群落。2017年和2018年两个年度中布氏田鼠种群密度明显增加,捕获率分别达到25.60%和75.80%,极显著高于其他种群数量(P<0.001),占群落中个体数均达到90%以上,成为禁牧区绝对优势种,啮齿动物群落为布氏田鼠+黑线仓鼠群落。因此,禁牧区虽然没有放牧干扰,但是5个年度的啮齿动物群落仍然出现了明显的动态演替,群落组成和结构明显改变,优势种由最初的黑线毛足鼠逐渐被黑线仓鼠和布氏田鼠所替代。
4 讨论
群落的时间格局是群落的动态演替特征之一,是由自然环境因素或者是人为干扰的时间节律动态所引起的群落各物种在时间上相应的过程变化。同一空间不同的时间尺度,物种的聚集是许多因子和生态过程作用的结果[11~13]。不同人为干扰下群落鼠类组成数量的变化,是生物物种适应不同干扰斑块并对相应干扰类型的综合反映。本研究中过度放牧、季节轮牧和禁牧3种不同干扰方式在5年的时间尺度中,啮齿动物群落组成中种类最多的为季节轮牧干扰,有6种啮齿动物,最少的是过度放牧干扰,有4种啮齿动物。究其原因,在于过度放牧区是啮齿动物生境破碎化严重的干扰区,而季节轮牧区具有较高的物种多样性,啮齿动物种类和数量趋于均衡发展,这符合群落的中度干扰假说(Intermediate disturbance hypothesis)[14],即群落的中度干扰增加异质性, 提高多样性和丰富度,严重干扰可能增大,但更可能是减小异质性。鼠类作为草地生态系统中必不可少的成员,在系统中不仅是消费者,还是物质和能量的天然传递者。因此,在草地利用中更应该注重系统的这种均衡,寻求“草—畜—鼠”之间的平衡,这样才有利于草地生态系统的健康发展。
当一种景观由单一、均质和连续的整体趋向于复杂、异质和不连续的斑块镶嵌体或嵌块的过程变化时,即出现破碎化[15]。干扰所形成的景观破碎化将直接影响到物种在生态系统中的生存和生物多样性维持[16~18]。景观破碎化将直接导致动物栖息生境的破碎化,也就是生境空间格局的改变[11, 19]。因此,放牧作为人为干扰活动可直接或间接地通过改变生境空间格局而改变生物群落间的相互关系,从而对生物群落和系统功能产生重大影响。在一定区域内,不同类型的斑块镶嵌形成了统一的平衡体,而人为干扰形成的斑块中的动植物往往适应了这种干扰,为此产生了有独特环境或资源的斑块,既与周边区域有不同的物种结构和成分,又与其保持相对平衡状态[19]。本研究3种不同干扰方式5年中啮齿动物群落均表现为组成啮齿动物群落的鼠类种群随着时间尺度发生了不同程度的演替,这种演替就是对干扰形成的不同生境斑块的适应过程以及对栖息地环境和资源保持相对平衡的状态。在这一过程中,不同的啮齿动物种群在群落中所起的作用并不完全不同,特别是典型草原的啮齿动物代表种布氏田鼠在2014~2017年均未在不同干扰样地中捕获到,而在2018年出现种群高峰,种群数量波动极大。这种由草地不同干扰引起生境变化导致啮齿动物群落发生演替及优势种出现替代的现象,实质上是生物群落的“上行调节”(Down-up regulation),这一调节过程主要有哪些环境因子或物种决定, 还有待于进一步对不同干扰斑块动态及群落响应机制进行研究。
5 结论
5.1典型草原草地的不同利用方式会直接影响啮齿动物群落的组成、结构和优势种,不同的草地利用方式造成啮齿动物群落不同的年度动态演替,出现群落组成、结构的明显改变和优势种替代。
5.2供试典型草原不同利用方式3个处理区组,经过4~5年草地啮齿动物群落的优势种均被布氏田鼠所替代。
