施氮对干旱条件下晚稻产量与生理的影响
2021-07-01黄慧娴沈天花傅军如周大虎贺浩华陈小荣
黄慧娴,沈天花,钟 蕾,杜 杰,傅军如,周大虎,贺浩华,陈小荣
(江西农业大学 农学院,作物生理生态与遗传育种教育部重点实验室,江西省超级稻工程技术中心,双季稻现代化生产协同中心,江西 南昌 330045)
作为重要的粮食类作物,水稻生长发育受内因与外因双重影响[1],水分和养分(氮)分别是农作物生长发育和产量形成的主要调节因子[2]。氮素与水分相互独立又相互影响,是一对既互补又相互制约的因子。农作物在发生水分胁迫的时候,会发生相应的生理生化反应和形态结构上的变化,这些生理和生化响应构成了对水分的适应机制,调节土壤养分的吸收、同化、利用和转运等[3-5]。
在我国,干旱等气候灾害是影响粮食安全的重要因子,近年来农业发展上为应对气候变化的挑战变革逐渐增多[6]。我国南方种植的多为耗水量较大的农作物如水稻等,季节性干旱常导致南方农作物处于水分亏缺胁迫下。另外,由于雨量分布不均,双季稻区在生长季节经常会遭遇长短不同的水分亏缺,造成不同程度的旱害和歉收,特别是在气候变暖已成为全球变化的必然趋势下[7],在我国长江以南的地区旱灾发生的频率增加。江西伏旱与秋旱发生时双季稻正处于幼穗分化期,已知水稻该时期作为穗生长敏感期,极易受到环境影响[8-9]。前人研究发现,幼穗分化期至齐穗期干旱胁迫后光合效率下降、脯氨酸与可溶性糖等渗透调节物含量增加,丙二醛等膜脂代谢物含量增加、籽粒灌浆速率减慢,有效穗数、总粒数、实粒数均较正常水分供应下降低,空粒数增加,对结实率影响最大,导致总产量降低[8-10]。
超级稻品种在南方稻区的推广推动了增产增收及品种的更新换代,新品种的干物质产能及产量潜力水平均得到提高,但生育期间对水肥需求更大,对水、氮亏缺与调控更为敏感[11]。同时有研究认为施氮将提高作物对水分亏缺的敏感性,使作物感受胁迫后响应更为剧烈[12],而这些研究是否同样适用于双季稻还未明确。有推测认为,水分调控与氮素运筹与作物类型、水分胁迫时期和程度及施氮量不同等相关[13-14],虽然人们对水稻水分与氮素互作及其耦合效应进行了不懈探讨,但具体关系尚不十分明确,双季稻水氮互作效应的机理也缺乏深入探讨。研究发现,氮肥、磷肥及钾肥的合理施用对于农作物抗、耐旱性均有一定的作用,而氮素使用运筹对植物抗、耐旱的效应问题在双季超级稻上较少有专门报道。同时,以往对植物干旱条件下的水氮耦合效应的研究通常是将氮素设置成不同水平,有关于双季稻同一氮素水平不同运筹对干旱的缓解效应的研究报道较少。
综上,开展氮素运筹对双季超级稻干旱条件下的影响及其有关生理机制的研究十分必要。本研究设置不同氮素运筹,从光合、氮素和干物质积累与分配,氮代谢有关酶活性、渗透调节物质等变化、产量等探讨旱前施氮及旱后补氮对超级杂交晚稻的产量形成及其有关生理指标的影响,为农作物生产应对干旱条件下合理的施肥(氮)策略提供理论依据。
1 材料和方法
1.1 试验材料
五丰优T025:江西省主栽的双季超级杂交晚稻(五丰A/昌恢T025,2010年农业部认定为超级稻),南昌地区生育期112.3 d左右。试验于2016,2017年在江西省南昌市江西农业大学科技园试验基地进行。
1.2 试验设计与材料种植
设置7个处理,采用桶栽方式。试验用桶参数分别为:24.0 cm(高)、29.0 cm(上部直径)、23.5 cm(桶底直径)。土壤取自稻田0~20 cm耕层,经自然晾晒风干,用FT-1000A土壤粉碎机(常州市伟嘉仪器制造有限公司)粉碎成0.15 mm的土壤颗粒,移栽前预淹水浸泡14 d。于6月24日播种,7月26日移栽,移栽选取长势良好、一致的秧苗移栽到桶内,每个处理各种植20桶,每桶3株(穴),各个处理共计140桶,移栽前在大田统一管理。正常水分不同氮素水平处理(T1/T2/T3)全生育期保持3~5 cm水层,干旱处理组(T4/T5/T6),模拟江西等气象干旱发生时期与历时,于9月20日(2016年)和9月13日(2017年)(幼穗长度约为1 mm时期)傍晚搬至玻璃房干旱控水,9月26日(2016年)和9月19日(2017年)移出复水,于11月1日(2016年)和10月28日(2017年)收获。本试验中氮素水平按180 kg/hm2氮肥折算,各处理氮肥分别按T1/T4=2∶6∶2、T2/T5=2∶2∶6、T3/T6=2∶4∶4(基肥∶分蘖肥∶穗肥)比例施肥,蘖肥于移栽后5 d施用,穗肥于复水后第2天施用(尿素用量分配具体见表1);磷肥全部在基肥时期施用,施肥水平均采用 P2O590 kg/hm2;钾肥施肥水平为 K2O 180 kg/hm2,采用基肥∶分蘖肥∶穗肥=5∶2∶3比例施肥,栽培与病虫害防治措施与大田相一致。产量及产量构成采用2 a数据,其余指标均使用2017年数据。前期土壤理化性质检测结果为:土壤pH值为5.44,有机质含量为23.1 mg/kg,全氮含量为1.12 g/kg,速效养分中N氮磷钾含量分别为101.00,8.85,90.80 mg/kg。

表1 氮肥(尿素)施用运筹Tab.1 Nitrogen fertilizer (urea) application ratio
1.3 测定指标及方法
1.3.1 产量及其构成 各处理于成熟期选取长势一致且未损伤水稻7株,将其茎、叶、穗分装,穗子部位于日光下自然晒干以备后期氮素测定。晒干后分别考察其单株产量和产量构成因素(总粒数、结实率、穗长、千粒质量、二次枝梗数等)。
1.3.2 干物质积累量测定 各处理于穗肥后即抽穗期(同旱后复水)和成熟期各选取5株长势一致的水稻,将茎、叶、穗分装以备后期研磨。茎叶穗放于烘箱105 ℃杀青后烘干,成熟期穗子部位于日光下自然晒干后统一称量计质量。
1.3.3 氮素含量测定 氮含量的测定依据凯氏定氮法[15],取样方法同1.3.2。
1.3.4 光合作用 在抽穗期和开花期9:00-11:00使用CI-340便携式光合作用测定仪(CID Bio-Science,Camas,WA,USA)测定叶片净光合速率(Pn)、蒸腾速率(Tr)、叶片气孔导度(Gs)、细胞间CO2浓度(Ci),每个处理每个重复选定4~6株水稻的倒二叶进行测定,取平均值。
1.3.5 渗透调节物质的测定 于抽穗期和开花期,各处理选取长势均匀的水稻叶片,对其进行测定。测定有关渗透调节物质:可溶性糖测定采用蒽酮比色法,可溶性蛋白测定利用BCA法,丙二醛(MDA)测定硫代巴比妥酸(TBA),脯氨酸(Pro)的提取利用磺基水杨酸(SA)法[16-18](测试所用试剂盒均由苏州科铭生物技术公司提供,具体操作参照说明书)。
1.3.6 氮代谢酶的测定 于抽穗期和开花期,各处理选取长势均匀的叶片,测定谷氨酸脱氢酶(GDH)[6-8]、谷丙转氨酶(GPT)[19]、谷胺酰胺合成酶(GS)[20]和硝酸还原酶(NR)[21]4种氮代谢酶活性(测试所用试剂盒均由苏州科铭生物技术公司提供,具体操作参照说明书)。
1. 4 数据统计与分析
农艺性状与生理数据采用Microsoft Excel 2010软件整理数据,运用SPSS 19.0(v 20.0,SPSS Inc. Chicago,USA)软件下t检验进行方差分析,P<0.05表示差异达到显著水平。
2 结果与分析
2.1 不同蘖、穗氮肥处理水稻产量及其构成的差异
表2中2017年的结果表明,正常水分下3个氮肥运筹处理单株产量表现为T1>T3>T2>CK,两穗肥重施处理T2和T3较蘖肥重施T1产量分别降低了29.95%,28.46%,且T1与T2、T3间产量呈显著差异水平,T2与T3间无显著差异,显示出正常水分条件下超级杂交晚稻分蘖期施氮的重要性;干旱条件下,两穗肥重施处理T5和T6较蘖肥重施T4产量差异达显著水平,分别较后者低38.89%,33.26%,说明双季晚稻旱后(穗分化期)补氮恢复效果不及旱前(分蘖期)重施氮处理。从产量构成上看,正常水分条件下,蘖肥重施处理T1较2个穗肥重施处理T2和T3只有单株有效穗数呈现显著差异水平,T1较T2、T3分别多38.07%,30.54%;干旱条件下,蘖肥重施处理T4与2个穗肥重施处理T5、T6单株有效穗数和结实率均呈显著差异水平,其中单株有效穗数T4较T5、T6分别多26.92%,5.60%,结实率分别高24.14%,33.33%。另外,正常水分与干旱条件下,蘖肥重施处理与2个穗肥重施处理间穗长、二次枝梗数、穗粒数、千粒质量等指标间的差异各存在不同特点,但干旱条件下这些指标在蘖肥重施处理与穗肥重施处理间总体差异不明显。2016年的结果与2017年整体趋势较为一致。干旱条件下,T4较T5、T6的单株产量高35.83%,20.60%。
2.2 不同蘖、穗氮肥处理水稻干物质积累的差异
图1可知,无论是正常水分还是干旱条件下,水稻抽穗期各处理干物质积累量总体表现为氮素蘖肥重施处理高于穗肥重施处理,干物质积累量与前期分蘖肥呈现正相关,分蘖肥充足,穗部干物质积累快。成熟期正常水分处理组仍保持与抽穗期一样的趋势,干物质总积累量表现为T1>T3>T2,可见前期供氮充足有助于干物质积累和籽粒充实。幼穗分化期干旱条件下,成熟期干物质积累量显著低于正常水分处理,T4较T1、T5较T2、T6较T3分别下降19.2%,15.94%,22.31%;但不难发现,干旱条件下无论是总干物质积累量,还是穗部干物质积累量,均以T4处理最高。
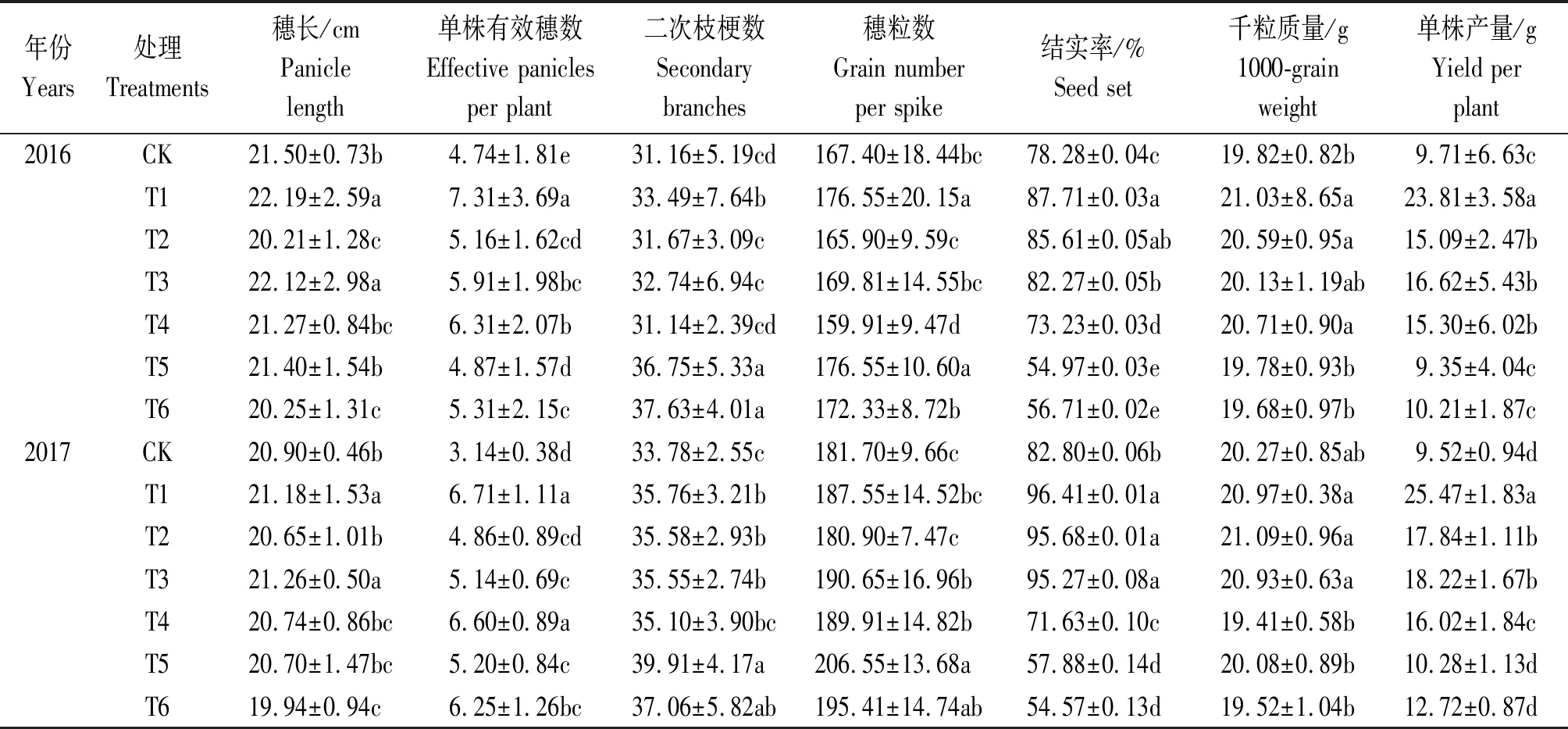
表2 水稻各处理下产量及产量构成Tab.2 Yield and its composition of rice for the different treatments
2.3 不同蘖、穗氮肥处理水稻氮含量的差异
图2表明,正常水分条件下,抽穗期不同蘖、穗氮肥处理以T1总氮含量最高,T1与T2、T3间差异显著,且T1氮含量大多集中在叶片部位;幼穗分化期干旱条件下,3个不同蘖、穗氮肥处理地上部氮含量无显著差异。无论是正常水分,还是幼穗分化期干旱,至成熟期稻株氮含量均以氮素蘖肥重施处理最低,氮素穗肥重施处理显著高于前者,尤其以茎、叶部差异明显,表明氮素穗肥重施处理将导致更多的氮素积累在茎、叶部,向穗部的转运相对较少。
2.4 不同蘖、穗氮肥处理水稻光合的差异
表3表明,无论是正常水分还是干旱条件下,水稻抽穗期、开花期各处理净光合速率(Pn)均是以氮素蘖肥重施处理T1与T4最低,显著低于氮素穗肥重施各处理;幼穗分化期干旱条件下,氮素蘖肥重施处理T4抽穗期和开花期的蒸腾速率(Tr)也显著低于氮素穗肥重施各处理,叶片气孔导度(Gs)、细胞间CO2浓度(Ci)也表现类似差异特点;另外,4个光合指标在幼穗分化期干旱条件下从抽穗期到开花期均不同程度上升。
2.5 不同蘖、穗氮肥处理水稻渗透调节的差异
图3表明,抽穗期和开花期各处理可溶性糖、可溶性蛋白、丙二醛与脯氨酸含量呈明显的差异。幼穗分化期干旱条件下,氮素蘖肥重施处理,抽穗期脯氨酸含量较高;开花期持有高水平的可溶性糖含量与较高的可溶性蛋白含量,虽然T4丙二醛含量高于T6,但明显低于T5。正常条件下,开花期T3处理的渗透调节物质及丙二醛含量最高;在抽穗期,可溶性糖和丙二醛含量在蘖肥重施处理中相对较高,可溶性蛋白和脯氨酸含量在蘖肥重施中相对较低。

表3 水稻各处理下光合指标Tab.3 Photosynthetic indexes of rice for the differentiation treatments
2.6 不同蘖、穗氮肥处理水稻氮代谢酶活性的差异
图4表明,各处理抽穗期和开花期的硝酸还原酶活性、谷氨酰胺合成酶、谷氨酸脱氢酶和谷丙转氨酶活性均表现出明显的差异,开花期较抽穗期硝酸还原酶活性和谷丙转氨酶活性均表现出规律性下降;其中相较于正常水分,幼穗分化期干旱条件下不同氮素蘖、穗肥处理抽穗期的硝酸还原酶活性和谷丙转氨酶活性更高;同时,硝酸还原酶和谷丙转氨酶活性在抽穗期T1>T2,T4
3 讨论与结论
3.1 幼穗分化期干旱与否条件下水稻产量及构成因素与不同蘖、穗氮肥处理的关系
本研究发现,无论是哪种蘖、穗氮肥比例处理,幼穗分化期干旱后,水稻产量均大幅下降,与Ding等[23]研究结果一致,显示出双季晚稻幼穗分化期对水分亏缺的敏感性。穗分化期干旱或正常水分条件下,氮素蘖肥重施较氮素穗肥重施水稻产量均有明显的优势,与孙永健等[24]研究指出的随氮肥穗肥后移比例的增大将导致水稻产量的显著下降结果相一致,表明了双季晚稻氮素蘖肥的重要性,分析其原因,可能与双季晚稻由于受到生育期的限制,而氮素蘖肥适当重施条件下有利于分蘖早生快发,提高成穗率,从而增穗增产,进一步分析产量构成可发现,氮素蘖肥重施不仅有利于改善分蘖成穗,还将保障结实率的稳定;而幼穗分化期干旱条件下,随着氮素穗肥比例的增多,复水后新生蘖大幅增加,但由于随后生长期的有限性,部分新生蘖无法抽穗,形成无效蘖,耗费营养,还有部分虽然抽穗,但由于随着气温的下降,结实率大幅下降。因此,幼穗分化期干旱条件下更应保证氮素蘖肥的充分供应,充分利用“以氮调水”效应,保证该逆境胁迫下的产量。
3.2 幼穗分化期干旱与否条件下水稻干物质、氮素积累与不同蘖、穗氮肥处理的关系
干物质作为衡量产量的重要相关性指标,与产量呈显著正相关。本研究表明,抽穗期干物质积累量与氮素蘖肥呈正比,分蘖肥充足,穗部干物质积累快,分蘖成穗率高,这与前人的研究相一致[25];成熟期各处理生长中心及干物质积累均转向穗部,前期肥水充足处理表现出更为稳定的茎、叶物质向穗部转运趋势。幼穗分化期干旱条件下,各处理的穗部干物质积累相对正常水分处理减少,而氮素蘖肥重施处理的稻株穗部积累量显著高于穗肥重施处理。幼穗分化期干旱条件下,成熟期茎、叶氮素积累量占地上部总积累量的比例均以氮素穗肥重施处理高,这与Singh等[26]研究发现的旱后植株内滞留的氮素增加一致。总体看来,幼穗分化期干旱条件下,各蘖、穗氮肥处理的穗部干物质和氮素积累与魏征等[27]和汤国平等[28]提出的旱后补偿效应、氮素补偿机制存在一定的差异,这可能与水稻幼穗分化期氮素和水分亏缺较其余生育时期更为敏感与复杂有关,其机制值得深入探究。
3.3 幼穗分化期干旱与否条件下水稻光合与不同蘖、穗氮肥处理的关系
前人研究发现植物光合速率随氮浓度增加而提高,但随土壤含水率减少而下降。在很多作物中都观察到氮水互作对光合作用的影响[29]。本研究表明,正常水分条件下,随生育进程的推进,氮素穗肥的增加,净光合速率下降趋势将减缓,与Liu等[30]提出的氮素营养增加更能保持后期光合速率稳定相一致。水稻在幼穗分化期遭遇严重干旱后,通过限制气孔开度,抑制PSⅡ的光反应和电子传递过程,胞间二氧化碳浓度和净光合速率等参数降低。旱前重施氮肥提高了光合色素和二磷酸核酮糖羧化酶等相关蛋白的合成和表达,维持较高的电子传递速率,从而保护光合作用的生物化学过程,水稻对干旱胁迫的适应性。
各蘖、穗氮肥处理表现为复水后短期内净光合速率上升,与Zhou等[31]对水稻土壤水分亏缺后的研究相一致,推测认为受干旱胁迫的影响,旱后氮素吸收转运集中在叶片,叶绿素浓度升高,PSⅡ原始光能转换效率(Fv/Fm)和实际光合效率Y(Ⅱ)上升。旱后各氮素穗肥重施处理净光合速率、蒸腾速率、气孔导度在开花期上升,说明水稻幼穗分化期旱后复水后重施穗肥,在开花期逐渐表现出对水稻叶片光合一定的生理补偿效应,但不同氮肥处理的生理补偿效应的强度以及发生时间出现差异[31]。然而,氮素穗肥重施处理下光合作用出现一定补偿效应,但光合产物在茎、叶部的滞留不容忽视,相对于氮素蘖肥重施处理,产量还是受到更大的影响。
3.4 幼穗分化期干旱与否条件下水稻渗透调节与不同蘖、穗氮肥处理的关系
水分亏缺作为一种环境变化引起的胁迫,不同植物在应对时有不同的机制,前人研究表明,在失水细胞损伤后会产生一系列的过氧化产物如丙二醛等,但同时也会产生的一些具有脱水保护功能的可溶性蛋白和糖类以调节减缓损伤[32]。在水稻土壤水分下降到50%以后,稻株会为了减缓胁迫影响而合成许多新的糖类物质,这些可溶性糖一部分作为渗透调节物质起作用,一部分作为营养物质,同时影响蛋白的合成与积累,胁迫下植物还可以通过积累一定量的脯氨酸以降低水势,维持体内的水分平衡[33-34]。
本研究发现,各氮素处理穗分化期复水后初期叶片保护物质可溶性糖与脯氨酸含量上升,至开花期下降,与戴高兴等[33]研究结果一致。而丙二醛与可溶性蛋白在水稻穗分化期干旱胁迫后,随着生育进程的推进以及氮素的吸收利用,其含量反而逐渐增加,推测认为,水稻幼穗分化期干旱复水后由于膜脂伤害加剧,过氧化产物丙二醛增加,并与氮素吸收后产生的滞留氮呈正比,这可能是水分亏缺促进了氮素物质的再利用,加速最初功能叶片的衰老进程,促进了物质的运转,前人有关研究也出现类似推论[35]。氮素蘖肥重施中抗氧化酶活性提高有利于缓解叶片损伤,导致开花期丙二醛含量较穗肥重施处理显著下降,提高氮肥施用水平可有效增强植物的抗氧化能力[36]。同时,氮素蘖肥重施处理在开花期持有更高水平的可溶性糖含量,显示出不同氮素蘖、穗肥比例处理下稻株渗透调节的差异性,可能是其应对穗分化期干旱胁迫的重要生理机制之一。
3.5 幼穗分化期干旱与否条件下水稻氮代谢酶活性与不同蘖、穗氮肥处理的关系
水稻产量的形成离不开氮素的吸收与利用,后者又受到氮代谢酶的调控。本研究表明,干旱条件下抽穗期各氮代谢酶相对保持更高的活性。幼穗分化期干旱条件下,不同处理抽穗期硝酸还原酶与谷丙转氨酶活性在穗肥重施处理中上升,与孙永健等[37]结果相一致;谷氨酸脱氢酶活性变化则表现为在开花期升高,与Goel等[38]苗期干旱试验中活性升高相类似。说明各氮代谢酶活性与氮素施用有关,而其对产量的影响,则与氮素的转运与穗部的积累有关,较为复杂。

综上所述,双季晚稻幼穗分化期干旱条件下,干旱前(分蘖期)适当地重施氮肥较干旱后(穗分化期)重施氮肥可在一定程度保持较高的单株有效穗数和结实率,防止干旱条件下的稻株大幅减产,前者较后者存在较明显的“以氮调水”效应,原因主要是氮素蘖肥重施有利于生育后期氮素及干物质的转运及向穗部积累,虽然其生育后期净光合速率、氮代谢酶活性等低于氮素穗肥重施处理。
