滆湖入湖河口区叶绿素a时空变化特征及相关环境因子分析
2021-07-01张毅敏高月香张志伟朱月明钱文瀚
刘 钰,杨 飞,张毅敏①,高月香,张志伟,朱月明,孔 明,赵 远,钱文瀚
(1.常州大学环境与安全工程学院,江苏 常州 213164;2.生态环境部南京环境科学研究所,江苏 南京 210042)
滆湖是中国苏南地区的第二大淡水湖。全湖大部分的水面区域位于江苏省常州市武进区境内(占武进区总面积71.4%),多年平均最大水位深度位于吴淞基面,约为3.30 m,相应的平均水深为1.25 m,面积约为166.7 km2,容积量约为2.05×108m3[1]。2018—2020年监测数据初步显示,目前滆湖的ρ(TP)为0.218 mg·L-1,ρ(TN)为3.79 mg·L-1,已超GB 3838—2002《地表水环境质量标准》劣V类水质的污染程度和富营养水平。滆湖周边地区的快速发展使河道内的氮、磷等营养物质逐年增多,滆湖的水草资源受到一定程度的破坏,藻类生物量逐年升高,加重了滆湖的面源污染[2]。入湖河道的水质在很大程度上影响了滆湖的水质。因此,对滆湖入湖河口区域的污染防治研究已刻不容缓。
叶绿素a(Chl-a)是浮游植物生物体的重要组成成分之一,其浓度水平反映了浮游植物现存量的高低,与水环境质量密切相关,是水体理化性质动态变化的综合反映指标[3-4],测定ρ(Chl-a)能够在一定程度上反映水质状况。湖泊的入湖河口区是特殊区域,兼有河流和湖泊的特征。滆湖作为太湖上游的主要湖荡,是太湖的前置库,是太湖流域重要的行蓄洪湖泊,也是区域供水、生态调节的重要水域,其污染控制和水质净化对太湖水环境改善具有非常重要的意义。同时,滆湖为引江济太(新孟河)工程河湖相连的重要节点,经太滆运河、漕桥河入太湖。滆湖治理和修复模式对太湖有借鉴意义。鉴于此,为探明滆湖入湖河口水域中ρ(Chl-a)的时空变化及其影响因素,以13个滆湖入湖河口水域为研究对象,于2018年1月—2020年1月对各河口水质指标进行监测,探讨其对于ρ(Chl-a)分布的影响。重点分析了环境因子(如水温、N/P比、CODMn、营养盐等)对河口ρ(Chl-a)时空动态的影响,并探讨其内在关联,为筛选和识别影响入湖水体ρ(Chl-a)变动的主要环境因子提供科学资料,同时也为ρ(Chl-a)的预测及滆湖蓝藻水华预警等水环境保护工作提供一定的理论依据。
1 材料与方法
1.1 采样时间与样点设置
滆湖入湖河流有北干河、中干河、夏溪河等,出湖河流有太滆运河。在沿岸河道共设13个监测点位(图1):武南河入湖(点位1)、孟津(点位2)、夏溪(点位3)、塘门沟1#(点位4)、塘门沟2#(点位5)、 湟里1#(点位6)、湟里2#(点位7)、北干(点位8)、中干(点位9)、渎新(点位10)、灌渎(点位11)、亳渎(点位12)、太滆运河(点位13)。监测时间为2018年1月—2020年1月,每月采样1次,每次采样于09:00—12:00完成,各点位采样顺序保持一致。用上、下底均有阀门的有机玻璃采水器采集水下0.5 m深处的水样1 L,于当日在实验室进行各项指标的测定。
1.2 测定方法
监测项目主要包括水温(T)、pH值、DO浓度、Chl-a浓度、TN浓度、TP浓度、NH4+-N浓度、NO3--N浓度、PO43--P浓度、CODMn、溶解性有机碳(DOC)浓度。其中T、pH值、DO浓度使用YSI多参数水质检测仪在采样现场测定,其他指标采样后带回实验室分析测定。TN浓度采用过硫酸钾氧化紫外分光光度法,TP浓度采用钼锑抗分光光度法比色法测定。DOC浓度使用总有机碳分析仪ASI-L(日本岛津)测定。Chl-a浓度使用分光光度法中的乙醇法测定[5]。
1.3 数据分析
利用IBM SPSS Statistics 25软件对滆湖入湖口13个采样点Chl-a浓度与环境因子进行相关性分析以及水环境因子的单因素方差分析,对每个采样点3组平行的季度均值进行均一化处理,对环境因子进行对数转化,以减少不同量纲的影响;对Chl-a浓度与其关系密切的环境因子进行多元线性回归分析及F检验,验证方程有效性。
2 结果与讨论
2.1 入湖河口水文与水质时空分布特征
2.1.1入湖河口水文分布特征
根据常州市水环境监测中心以及武进区水利局监测资料,收集到常州滆湖入湖13个河口的流量情况。由表1可知,不同点位河口的平均流量差异很大,从0.89~12.46 m3·s-1变化不等。其中武南河入口、孟津、北干、中干和太滆运河的流量较大,渎新、亳渎和灌渎的流量居中,夏溪、溏门沟1#、溏门沟2#、湟里1#、湟里2#流量较小。滆湖入湖河口的流量自上游至下游大致呈现从高到低的变化特征。
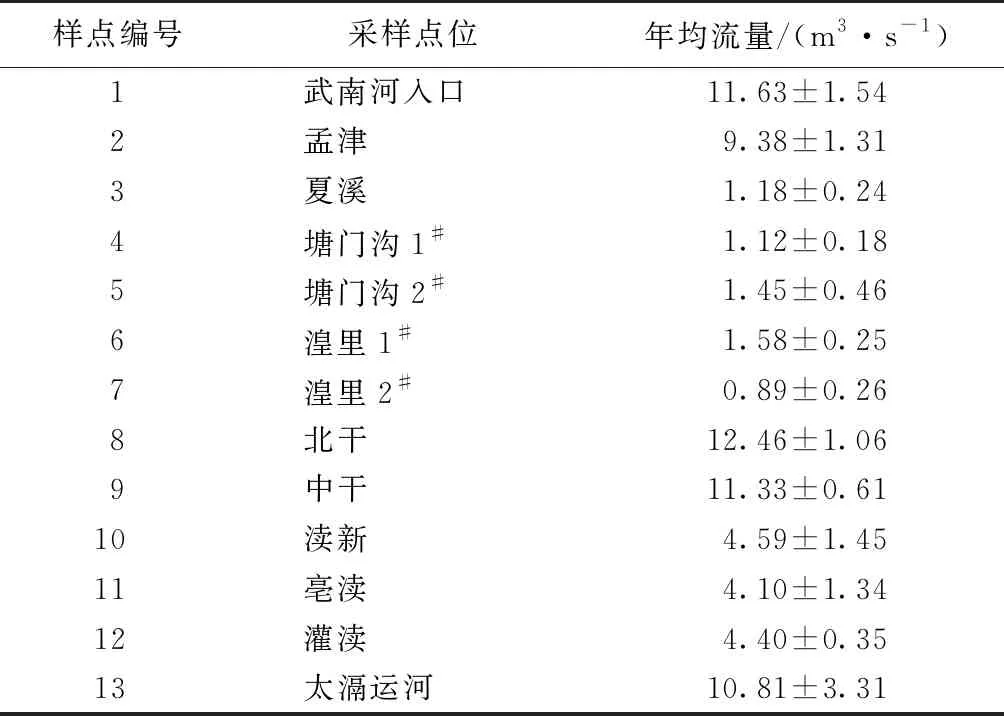
表1 滆湖入湖河口2018—2020年年均流量
2.1.2入湖河口水质时空分布特征
各入湖河口水质总体呈弱碱性。CODMn和ρ(TP)均值范围分别为5.26~6.94和0.24~0.55 mg·L-1,均属于Ⅳ类水;ρ(TN)均值范围为3.48~5.33 mg·L-1,属于劣V类水。由图2可知,入湖河口TN浓度季节变化规律大体表现为秋季>夏季>春冬季。其中,武南河入湖口、夏溪、中干、亳渎、太滆运河点位浓度较高。入湖河口ρ(TP)季节变化规律主要表现为春季最大,夏、秋季次之,冬季最小。空间上污染最为严重的区域主要集中于常州市武进区周边水系,与ρ(TN)的分布特征相似。CODMn总体呈现出冬季最高、夏秋季次之、春季较低的特征。
2.2 ρ(Chl-a)的时空变化特征
图3为各个季节各入湖口点位水体ρ(Chl-a)时空变化特征。整体来看,夏季湖泊水体ρ(Chl-a)比秋、春、冬季高,呈现较为明显的季节性变化。其平均浓度分别为17.14、10.61、7.80、4.21 μg·L-1(图3)。ρ(Chl-a)高值区主要分布于武南河入口、孟津、塘门沟1#、湟里1#、渎新。废水排放等原因影响了ρ(Chl-a)的空间分布特征[6],城镇集中且工业发达的塘门沟1#、湟里1#、渎新水质指标浓度较高,ρ(Chl-a)也较高。滆湖入湖河口ρ(Chl-a)总体上表现为夏季>秋季>春季>冬季。
综合来看,各入湖河口ρ(Chl-a)整体分布规律与水质时空分布规律较为接近,呈现较明显的季节变化和西高东低、北高南低的空间分布趋势。环境因子变化对于滆湖入湖河口ρ(Chl-a)的时空分布具有重要影响。
2.3 滆湖入湖口水体ρ(Chl-a)与流量的关系
河流流量作为水动力的重要指标,是影响河口区的大小、水质等的重要因素。于兰芳等[7]研究发现,ρ(Chl-a)与长江径流量存在较好的线性关系。笪文怡等[8]研究表明,千岛湖入湖流量与磷营养盐浓度存在显著相关关系,而营养盐的增加反过来促进了ρ(Chl-a)的升高。结合表1及图4可以发现,北干、中干和太滆运河流量较大,ρ(Chl-a)较低,这可能是因为水量的稀释效应。
仅武南河入口、北干、中干、灌渎和太滆运河ρ(Chl-a)表现出了明显的稀释效应,但对于其他河道来说,流量的影响并不明显。将各个点位的年均流量与ρ(Chl-a)进行相关分析,结果为不显著(r=0.061,P>0.05)。这可能是因为不同土地利用类型污染物的排放特征存在差别,例如北干、中干均属村镇及水产养殖密集的河道,水量交换缓慢,导致局部ρ(Chl-a)较高。将滆湖各入湖河口年均流量与ρ(Chl-a)进行线性拟合,得到年均流量(y)与ρ(Chl-a)(x)关系为:y=0.499x+10.718(r=0.061)。
2.4 滆湖入湖口水体ρ(Chl-a)与环境因子的相关性分析
ρ(Chl-a)在一定程度上反映了湖泊中浮游植物的生长状况,而浮游植物的生长又受到多种环境因子的影响和制约[9]。滆湖水体ρ(Chl-a)和环境因子之间的相关系数及检验结果(表2)表明,ρ(Chl-a)与水温、pH值、CODMn呈正相关,与ρ(DO)呈负相关,与ρ(TP)、ρ(TN)、ρ(NH4+-N)、ρ(NO3--N)、ρ(PO43--P)和ρ(DOC)无显著相关性;ρ(Chl-a)与环境因子之间的相关系数在0.157~0.637之间,与单个因子之间的相关系数均较低。以上结果表明,滆湖入湖口水体的ρ(Chl-a)变化受多个因子共同作用影响。

表2 滆湖入湖口各监测点位ρ(Chl-a)与环境因子之间的相关系数
2.4.1ρ(Chl-a)与理化环境因子的相关分析
由表2可知,ρ(Chl-a)与水温、pH值和CODMn呈显著正相关、与ρ(DO)呈显著负相关。葛优等[10]对阳澄西湖的研究表明,随水温的升高,ρ(Chl-a)逐渐增大,并在冬季达最小值。吕焕春等[11]对千岛湖水体为期1 a的监测结果表明,监测点位ρ(Chl-a)与水温呈显著正相关。滆湖13个入湖口ρ(Chl-a)皆与水温呈显著正相关,说明水温对浮游植物的生长代谢具有促进作用。浮游植物光合作用与呼吸代谢速率受水温的控制,从而影响ρ(Chl-a)[12-13]。滆湖入湖水体ρ(Chl-a)在全年监测中受季节变化的影响非常明显,水温的升高既促进了ρ(Chl-a)的上升,也有利于蓝藻形成优势[14-15]。
ρ(Chl-a)与pH值呈正相关。一般浮游植物生长茂盛的水体pH值较高[16-17]。这主要是由于浮游植物在光合作用时会吸收水中的CO2,同时释放出O2[18-21]。滆湖入湖口水体pH值在7.5~8.5之间,pH值不是影响ρ(Chl-a)变化的主要原因,只是ρ(Chl-a)变化的被动因子,随其含量增加而增加。
一般情况下,浮游植物的光合作用受水中ρ(Chl-a)影响,随ρ(Chl-a)上升,水体ρ(DO)也会增加[22-24]。但表2分析显示,ρ(Chl-a)与ρ(DO)呈负相关。出现这一现象可能是由于2个方面的原因:一方面,采样点位设置在较发达城镇的河道,养殖活动、生物及非生物好氧、残饵及排泄废物、污水排放等都会引起水体无机磷、硫化物、氨氮等浓度的升高,直接导致水体中DO浓度的降低;另一方面,河道出现藻类堆积现象,藻类腐烂需要消耗一定的DO,藻类大量漂浮聚集在水面,阻碍了覆氧,进而造成ρ(Chl-a)显著增高,同时使得水中的DO被消耗。尽管ρ(Chl-a)与ρ(DO)呈负相关,但是ρ(DO)也不是影响ρ(Chl-a)变化的主要原因。
ρ(Chl-a)与CODMn呈极显著正相关。当水体中ρ(Chl-a)升高时,浮游植物通过光合作用产生大量有机物,导致CODMn升高[25]。滆湖入湖口水体冬季CODMn为2.93~7.46 mg·L-1,夏季为4.05~9.36 mg·L-1。水体CODMn夏季略高于冬季,这可能是由于夏季水体中浮游植物大量繁殖,浮游植物在光合作用过程中会产生大量有机体,进而使水体的CODMn明显上升[26]。CODMn是ρ(Chl-a)变化的主要影响因子。
2.4.2Chl-a与营养盐环境因子的相关分析
湖泊中营养盐直接或间接影响其中浮游植物的生物量。SAKAMOTO[27]研究显示,高营养盐浓度导致浮游植物生物量增加,影响浮游植物群落分布。ρ(Chl-a)与氮、磷浓度的关系也较复杂,如齐凌艳等[28]对洪泽湖的研究发现,ρ(Chl-a)与N/P比呈正相关。张智渊等[29]基于GAM模型研究表明,ρ(Chl-a)与ρ(TP)、ρ(TN)无显著相关性。杨威等[30]对洱海的研究发现,ρ(Chl-a)与ρ(TN)呈极显著正相关,而与N/P比无显著相关性。
对滆湖入湖口水体ρ(Chl-a)、ρ(TN)、ρ(TP)及N/P比取对数后进一步分析发现ρ(Chl-a)的对数与ρ(TP)的对数呈显著正相关(r=0.339,P<0.01), 与N/P比的对数呈显著负相关(r=-0.401,P<0.01), 而与ρ(TN)的对数无显著相关(图5)。LÜ等[31]对武汉的15个城市浅水型富营养化湖泊的研究表明ρ(TP)与ρ(Chl-a)的相关性比ρ(TN)高,与笔者研究结论与其一致。
另外,入湖水体ρ(Chl-a)与ρ(NH4+-N)、ρ(NO3--N)无明显相关关系。在水体的自净过程中,由于浮游植物的消耗,NH4+-N会不断转化为NO3--N。然而ρ(NH4+-N)并没有因为浮游植物的消耗而减少,分析原因可能是水体不断受到NH4+-N污染,使得ρ(NH4+-N)并不随着浮游植物的增加而明显减少。
2.5 滆湖入湖口ρ(Chl-a)与主要环境因子的多元线性回归分析
前述分析可知,ρ(Chl-a)与水温、pH值和CODMn、ρ(TP)、ρ(DO)、N/P比之间关系较为密切。表2显示,ρ(Chl-a)与pH值呈显著正相关,与ρ(DO)浓度呈显著负相关,前者是因,后者是果,pH值、ρ(DO)不适宜作为自变量与因变量ρ(Chl-a)建立线性方程。因此在相关分析的基础上,选择对ρ(Chl-a)变化相对重要的因子N/P比、水温、ρ(TP)、CODMn建立和ρ(Chl-a)的多元线性回归方程,并进行显著性检验。
经F检验,R2=0.576,方差分析F值的显著水平P=0.000,表明回归显著有效。ρ(Chl-a)与原始变量的多元线性回归方程为ρ(Chl-a)=7.291-0.025×ρ(TN)/ρ(TP)+0.907×ρ(TP)+0.314×T+0.253×CODMn。
综合滆湖入湖口水体2018年1月至2020年1月期间各点位均值来看,入湖口Chl-a与筛选出的4个主要影响因子N/P比、T、ρ(TP)以及CODMn密切相关。将利用拟合公式计算的入湖河口ρ(Chl-a)和实测结果进行比较,计算值(y)与实测值(x)的线性回归方程为y=4.761 1+0.697 0x(r=0.853 8),通过回归方程计算的ρ(Chl-a)在实测误差范围内,说明回归方程得出的ρ(Chl-a)可以接受。
3 结论
以滆湖环湖13个入湖河流为研究对象,采用多元统计分析方法分析滆湖入湖ρ(Chl-a)的时空分异特征,识别出特征影响因子为TP、T、CODMn、N/P比,冬季和夏季湖泊水体ρ(Chl-a)平均值分别为6.66和17.10 μg·L-1,冬季湖泊水体ρ(Chl-a)空间分布与夏季具有一定的相似性,整个区域呈现较明显的西高东低、北高南低的分布趋势。
河口区水体ρ(Chl-a)与T、pH值、CODMn呈显著正相关,与ρ(DO)呈负相关。ρ(Chl-a)的对数与ρ(TP)的对数呈显著正相关,与N/P比的对数呈显著负相关,而与ρ(TN)的对数无显著相关。各监测点位ρ(Chl-a)随时间变化明显,1—4月较低,5—9月出现峰值,总体上看夏季最高,秋冬季次之,春季最低。
