低温胁迫下4份烟草品种苗期叶绿素及渗透调节物质含量的变化
2021-06-28杨超群高梓峰娄凤菊杜鑫宇朴世领
李 莹, 杨超群, 高梓峰, 娄凤菊, 杜鑫宇, 朴世领*
(1.延边大学农学院,吉林 延吉 13302;2.通化农业学校,吉林 通化 134001 )
低温是造成早花的主要原因,而早花是影响烟株品质的重要因素[1]。当烟草遭遇低温胁迫时,会由营养生长迅速转向生殖生长,进而导致叶片数减少,且叶片偏小、品质不佳,造成产量和品质降低[2]。低温胁迫下,烟草的理化性质会发生变化:1) 低温胁迫会影响植物光合作用和呼吸速率,从而抑制水分和养分的吸收,生物膜结构被破坏,最终致使细胞死亡[3]。2) 低温胁迫能降低叶绿体合成,相关酶活性抑制叶绿素的合成,造成植物光抑制现象产生,叶绿素含量的下降限制了植物光合速率,从而影响植物正常光合产物的合成[4]。
植物可以调节渗透来维持细胞内外平衡,调节细胞内pH值维持正常代谢从而维持反应平衡响应低温胁迫。植物体内游离脯氨酸在正常生长条件下含量很低,但当植物受到逆境胁迫后,游离脯氨酸大量积累[5]。有关研究指出[6-8],在低温胁迫下植物脯氨酸含量增加的幅度越大,其抗寒性越强。植物细胞中存在的可溶性糖与植物抗寒性密切相关[9],受低温的作用后可溶性糖含量会升高,它不仅是细胞代谢过程中的产物,也是调节植物生长发育的信号,其信号通路与胁迫通路相互作用形成复杂的网络,调节植物的代谢反应。可溶性蛋白能与水结合,可在低温胁迫下维持细胞内部的水分,此外,可溶性蛋白也可以通过影响基因的表达来提高植物的耐寒性[10]。
植物的耐寒性一直是植物抗逆研究的热点,温度是限制我国北方烟草生长最主要的影响因子,因早春低温导致幼苗长势差,诱导早花,严重影响烟叶的产量与质量。该文旨在探讨低温胁迫下渗透调节物质的变化规律及筛选耐寒性强的烟草品种,为烟叶生产提供理论依据。
1 材料与方法
1.1 材料及仪器
以烤烟品种龙江981、NC89及晒烟品种大兰花、延晒9号为试材,通过人工气候箱对4份烟草品种进行低温处理。
1.2 试验设计
将4份烟草种子在温室进行播种,烟苗达到7~8叶龄时,放入人工气候箱。处理组设置昼夜温度均为9 ℃,对照组设置昼夜温度均为24 ℃,昼夜湿度均为60%,设置昼夜光照为7 500 lx/0 lx,持续时间为9 d,分别在1.5、3、4.5、6、7.5和9 d 6个低温胁迫时间进行取样。每个阶段取由上至下第3~6片叶作为试验材料,3次重复。4份烟草品种低温处理的设计方案见表1。

表1 试验设计方案
1.3 试验测定以及计算方法
1.3.1 干物重、叶绿素测定
每个处理阶段取长势一致的烟苗3株,洗净表面灰尘后同时放入DHG-9240A型电热恒温鼓风干燥箱中(65~80 ℃),烘至恒重,冷却后测定质量即为干物重。叶绿素含量通过叶绿素仪(SPAD-205)测定。
1.3.2 渗透调节物质测定
采用蒽酮法[11]测定烟叶可溶性糖含量。采用考马斯亮蓝法[11]测定可溶性蛋白含量。采用茚三酮比色法进行测定游离脯氨酸含量[11]。
1.4 数据分析
1.4.1 隶属函数法
采用隶属函数法[12],对不同品种低温处理9 d时生理指标数据进行计算,按照耐寒能力的强弱进行排序。用隶属函数进行综合评价,能对各品种的各项指标进行综合分析,从而对品种做出耐寒性评价。
1.4.2 数据分析
低温下所有品种的生理生化指标利用Excel进行简单处理,利用SPSS 20对试验数据进行统计分析,采用Duncan多重比较法进行差异显著性分析。
2 结果与分析
2.1 低温胁迫进程中4份烟草品种苗期干物重、叶绿素含量的变化
2.1.1 低温胁迫进程中烟草苗期干物重的变化
不同烟草品种在低温胁迫下干物质的变化规律分析结果表明(表2):与对照相比,低温处理品种干物重增长较慢,低温处理1.5 d时所有品种对照与相应处理之间均无显著性差异(P>0.05),低温处理3~9 d期间,除3 d时T1(龙江981)外,其他品种的对照与其相应处理间均存在显著性差异(P<0.05)。低温处理9 d时,干物重较高的为T1(龙江981)与T4(延晒9号),分别为0.86和0.84 g。

表2 低温胁迫下不同烟草品种干物重的变化
不同烟草品种低温持续下干物重的增幅变化可知(图1),随着低温胁迫的延长,与对照相比所有品种干物重降幅随时间延长而不断增大。在胁迫1.5 d时,仅有龙江981干物重含量为增幅,其他均为降幅。在3~9 d时,龙江981与延晒9号干物重含量降幅始终较小,NC89与大兰花降幅始终最大,龙江981与延晒9号降幅之间无显著性差异(P>0.05),但龙江981与NC89、大兰花之间均存在显著性差异(P<0.05)。
2.1.2 低温胁迫进程中烟草苗期叶绿素含量的变化
不同烟草品种在低温胁迫下叶绿素变化规律分析结果表明(表3),当低温胁迫不断加剧时,4份处理的烟草品种叶绿素含量均呈下降趋势,而对照处理逐渐上升,低温胁迫的4.5~9 d时,T2(NC89)、T3(大兰花)与其相应对照之间存在显著性差异(P<0.05),而在7.5~9 d时,T1(龙江981)、T4(延晒9号)才与相应对照之间出现显著性差异(P<0.05),说明龙江981与延晒9号受低温胁迫后仍能较好的保持体内叶绿素的含量,而NC89、大兰花较差。
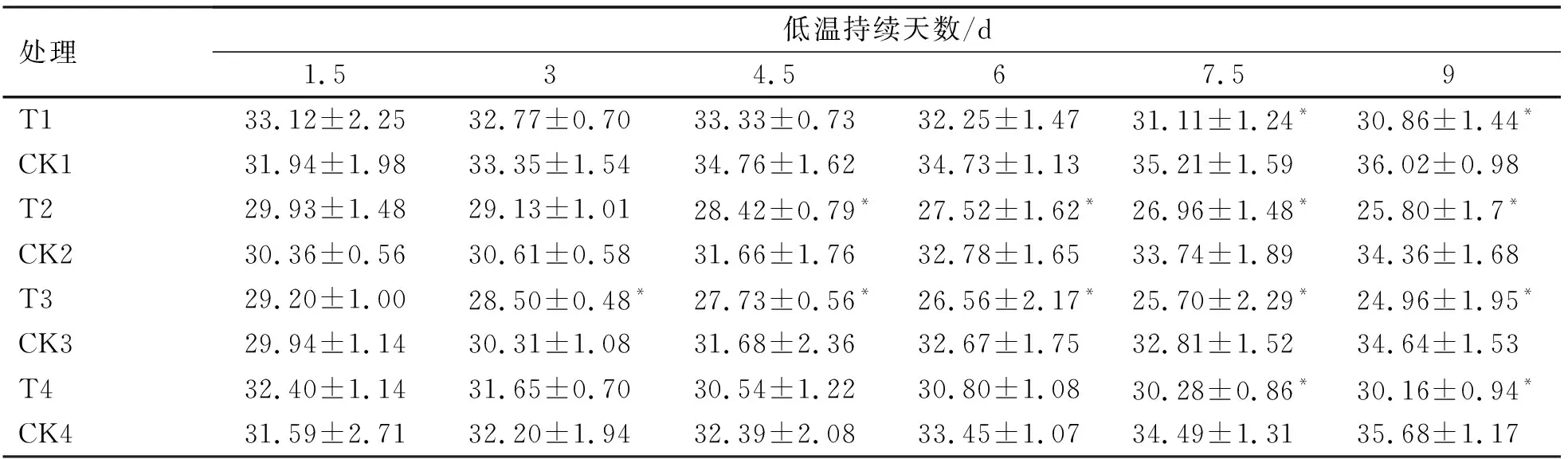
表3 低温胁迫下不同烟草品种叶绿素含量的变化
由表3和图2可知,低温胁迫下4份烟草品种叶绿素含量变化规律与干物重变化规律基本一致。在4.5~9 d期间,龙江981与NC89、大兰花叶绿素含量降幅之间始终存在显著性差异(P<0.05),而与延晒9号之间无显著性差异(P>0.05)。胁迫9 d时,所有品种降幅均达到最大,降幅由小到大依次为龙江981(-14.31%),延晒9号(-15.48%)、NC89(-24.93%)、大兰花(-27.95%)。
2.2 低温胁迫进程中4份烟草品种苗期渗透调节物质含量的变化
2.2.1 低温胁迫进程中烟草苗期Pro含量的变化
脯氨酸可以使细胞水势增高保持细胞与外界压强的稳定[13]。从表4可以看出,不同烟草品种在低温胁迫下,所有低温处理脯氨酸含量均随着胁迫天数的增加而不断升高,而对照处理一直保持平稳的水平;1.5 d时,T2(NC89)、T3(大兰花)、T4(延晒9号)与其相应对照之间无显著性差异(P>0.05),而T1(龙江981)与其对照之间存在显著性差异(P<0.05);低温处理至9 d时,T1(龙江981)与T4(延晒9号)的脯氨酸含量较高,分别为46.93和45.30 U/g,而T3(大兰花)的含量最低,为37.19 U/g且与其对照之间存在显著性差异(P<0.05)。
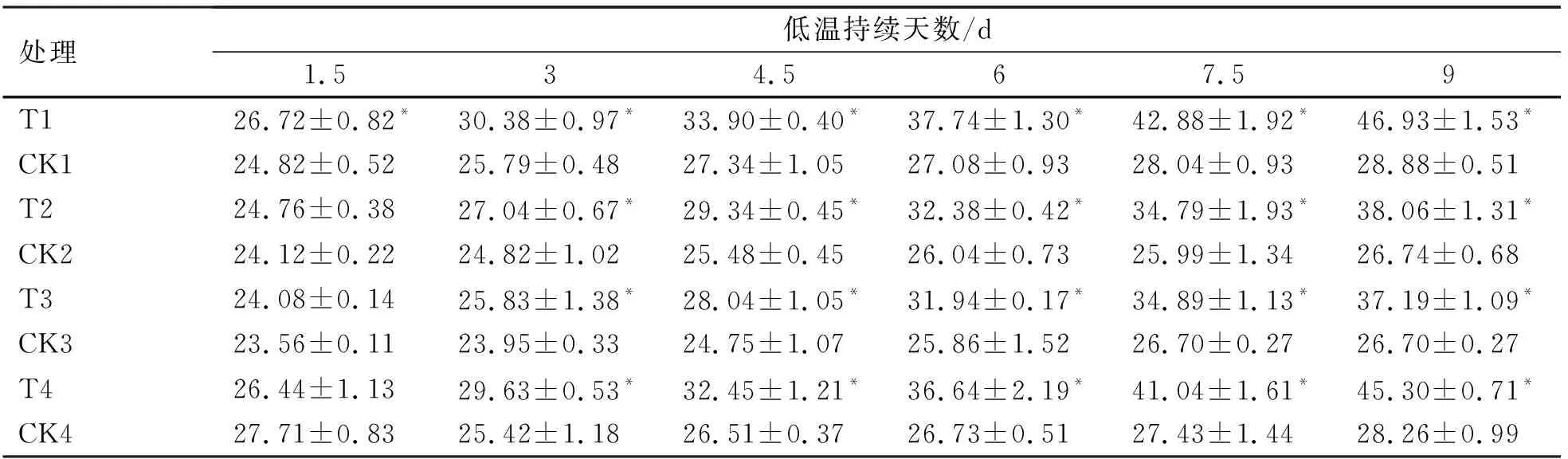
表4 低温胁迫下不同烟草品种Pro含量的变化
由图3可知,在低温胁迫的期间龙江981的脯氨酸含量增幅始终最高,而大兰花的增幅始终最低。在3~9 d期间,龙江981与NC89、大兰花增幅之间均有显著性差异(P<0.05)。由此可知,龙江981的耐寒性最强,而大兰花的耐寒能力最弱。
2.2.2 低温胁迫进程中烟草苗期可溶性糖(SS)含量的变化
低温胁迫下植物体内可溶性糖含量会不断积累[14-15]。4份烟草叶片中的可溶性糖含量在低温胁迫的环境下发生了显著的变化(表5),低温处理的烟苗可溶性糖含量变化呈逐渐上升的趋势,而对照处理缓慢上升。1.5~3 d期间,T2(NC89)、T3(大兰花)与其相应对照之间均无显著性差异(P>0.05),而T1(龙江981)、T4(延晒9号)在低温胁迫过程中始终与其相应对照有显著性差异(P<0.05);9天时,各个品种可溶性糖累积量均达到最高,T1(龙江981)与T4(延晒9号)可溶性糖累积量最高且显著高于其他低温处理,T3(大兰花)的累积量最低,且与其对照之间存在显著性差异(P<0.05)。
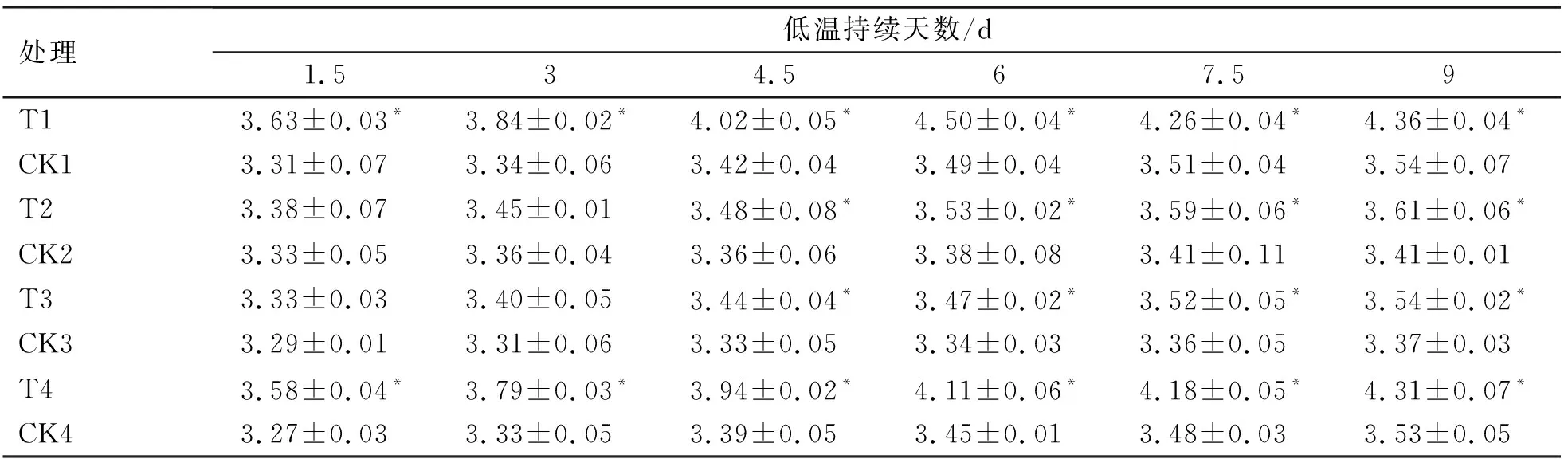
表5 低温胁迫下不同烟草品种可溶性糖含量的变化
不同烟草品种在低温胁迫下可溶性糖的增幅变化规律如图4。可溶性糖的增幅随胁迫的延长逐渐上升。在1.5~9 d期间,龙江981与延晒9号增幅之间始终无显著性差异(P>0.05),并且显著高于其他品种(P<0.05),NC89和大兰花增幅之间始终无显著性差异(P>0.05)。9 d时,龙江981与延晒9号的增幅较高,说明其耐寒性较好,而NC89与大兰花其体内可溶性糖含量积累较少,耐寒性较弱。
2.2.3 低温胁迫进程中烟草苗期可溶性蛋白(SP)含量的变化
可溶性蛋白能与水结合,可在低温胁迫下维持细胞内部的水分,此外,可溶性蛋白也可以通过影响基因的表达来提高植物的耐寒性[16]。低温胁迫下4份烟草叶片可溶性蛋白含量发生了明显的变化(表6)。所有低温处理的可溶性蛋白含量不断下降,且与相应对照处理之间均有显著性差异(P<0.05)。低温胁迫至9 d时,T1(龙江981)与T4(延晒9号)可溶性蛋白的含量高于其它低温处理,其中T3(大兰花)的可溶性蛋白含量最低。
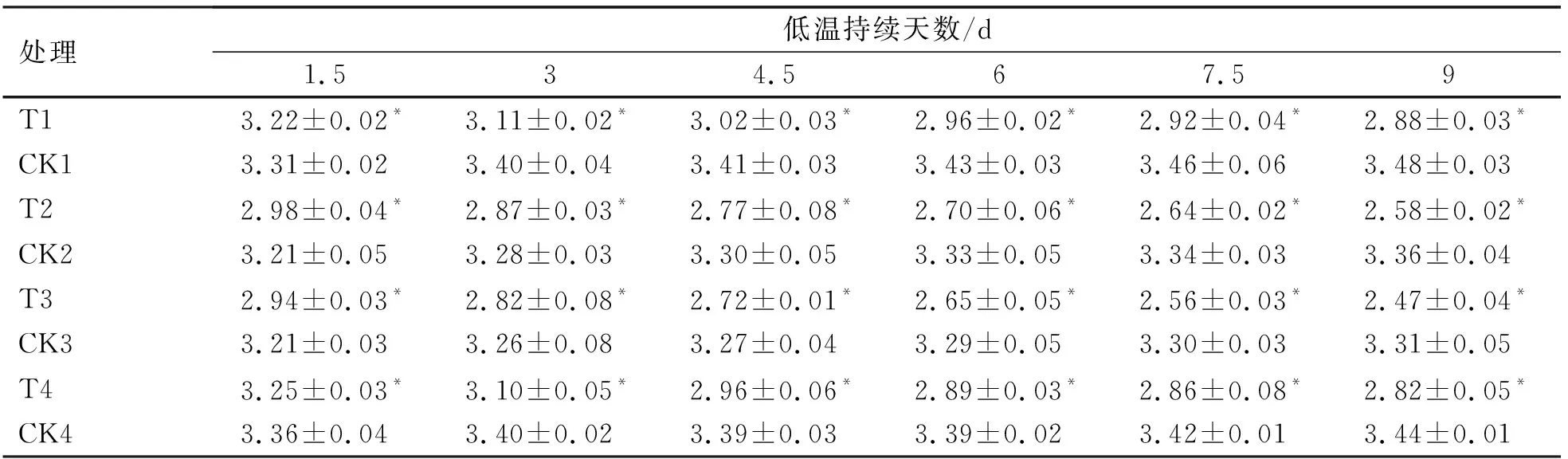
表6 低温胁迫下不同烟草品种可溶性蛋白含量的变化
由图5可知,随低温胁迫的延长可溶性蛋白含量降幅越来越大。在低温胁迫期间,龙江981与延晒9号之间始终无显著性差异(P>0.05)。低温胁迫至9 d时所有品种的降幅均达到最大。其中,龙江981的降幅最小(-17.12%),而大兰花的降幅最大(-25.56%)。
2.3 低温胁迫9 d时,隶属函数值及综合排序
4份烟草品种在低温胁迫9 d时的隶属函数值均采用模糊数学中隶属函数计算公式,先将9 d的生理生化指标测定值的变化幅度进行定量转换为隶属函数值,再将其隶属函数值取平均进行比较。由表7可知,低温胁迫9d后隶属函数值排序由大到小依次为龙江981、延晒9号、NC89、大兰花。龙江981的隶属函数值最高,为0.724,大兰花的隶属函数值最低,为0.436。

表7 低温胁迫9 d时隶属函数值及排序
3 讨论与结论
烟草在低温胁迫条件下其生长受到抑制,随着低温胁迫的延长植物生长愈加缓慢甚至停滞[17]。而叶绿素作为植物光合作用的重要物质,其含量的多少直接影响植物光合能力的强弱。该试验结果表明,随着低温持续天数的延长,4份烟草品种干物重及叶绿素含量均呈下降趋势,其降幅由小到大依次均为龙江981、延晒9号、NC89和大兰花。说明植物体受低温胁迫程度越大,其体内叶绿素含量越低,导致植物光合能力减弱,影响干物质的积累减少。
渗透调节是植物在逆境胁迫时出现的一种调节方式,由细胞生物合成和吸收积累某些物质来完成其调节过程。渗透调节物质主要包括可溶性糖、可溶性蛋白、脯氨酸和甜菜碱等。植物细胞中存在的可溶性糖不仅是细胞代谢过程中的产物,也是调节植物生长发育的信号[18],在低温胁迫的条件下会不断的积累有利于提高植物的抗逆性[19],同时,可溶性糖可以为植物代谢提供大量的碳源,能够在逆境下为各种代谢提供基础,从而增强植物的耐寒性[20]。脯氨酸作为渗透调节物质能保持环境渗透平衡。游离脯氨酸能促进蛋白质的水合作用,蛋白质胶体亲水面积增大,保护酶的空间结构为生化反应提供足够的生理活性物质,从而对细胞起到一定的保护作用。李建设等[21]以茄子为材料的试验中得出,茄子在低温胁迫后,脯氨酸的含量会大幅度上升,这说明脯氨酸与耐寒性密切相关,可以作为烟草耐寒性评价的指标之一。该试验结果表明,随低温胁迫时间的延长,4份烟草品种的可溶性糖与脯氨酸含量均逐渐增加。其中,龙江981可溶性糖与脯氨酸含量增幅在各个时期均最大,而大兰花增幅始终最小。可溶性蛋白的亲水性较强,能增加细胞的保水能力,从而提高植物的抗寒性。抗寒性强的品种可溶性蛋白含量高,反之则低。该试验结果表明,随低温胁迫的延长,可溶性蛋白含量呈下降趋势,降幅由大到小依次为大兰花、NC89、延晒9号和龙江981,与陈卫国[22]的研究结果相近。
该试验采用隶属函数法对4份烟草品种的耐寒性进行排序。隶属函数是采用模糊数学理论来解决问题,是模糊集合的特征函数。隶属函数分析提供了一条在多指标测定基础上,对各植物特性进行综合评价的途径[23]。该试验基于隶属函数的耐寒性综合评价结果为4份烟草品种耐寒性由强到弱依次为龙江981、延晒9号、NC89和大兰花。
综上所述,龙江981烟草品种耐低温能力最强,而大兰花对低温敏感且耐寒性最差。
