低氮胁迫下不同氮效率玉米品种的氮代谢与物质生产差异
2021-06-28袁继超
李 强,任 云,邹 勇,刘 静,袁继超
(1.重庆文理学院 园林与生命科学院,特色植物研究院,重庆经济植物生物技术重点实验室,重庆市特种植物协调创新中心,重庆 402160;2.四川农业大学 农学院,农业部西南作物生理生态与耕作重点实验室,成都温江 611130)
玉米是世界上第一大粮食作物,全世界玉米总产量已超过10×1012kg,占世界粮食总产量的41%,玉米生产对全球粮食安全起着举足轻重的作用[1-3]。氮是玉米生长发育所必需的大量营养元素,也是限制作物生长和产量形成的首要因素[4-5]。增施氮肥是提高玉米产量最简单有效的方式,因此在玉米生产过程中常见过量施用氮肥现象[6-7]。氮肥的过量施用不仅会使玉米产量和品质显著下降,更会造成氮肥利用率降低,生产成本增加及环境污染等一系列问题[8-9]。在不增加或减少氮肥施用的前提下,进一步提高玉米的单产,对于保障全球粮食安全和缓解环境污染具有重要意义[10]。氮高效玉米品种的选用是提高玉米单产,解决氮肥过度施用最重要的途径之一[11-12]。
为了提高氮素利用效率,植物体不仅需要高效的氮素吸收机制,还需要将积累的氮素在体内高效转运和利用[13-15]。植物吸收氮素后,通过硝酸还原酶(NR)、亚硝酸还原酶(NiR)、谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)以及一些氨基转移酶、尿素酶等的作用转运氮素到储藏器官中供植物生长发育所需[16-17]。牛巧龙等[18]指出,玉米硝酸还原酶活性随施氮量增加呈单峰曲线变化,施氮量为180kg·hm-2时,叶片NR活性最高。Bertrand等[19]研究认为,NR活性高的玉米品种ATP含量也高,叶片中NR活性可以作为氮代谢、子粒产量和蛋白质含量的选种指标。李文龙等[17]研究表明,氮高效品种‘郑单958’的硝酸还原酶、亚硝酸还原酶、谷氨酸合成酶和谷氨酰胺合成酶的活性均显著高于氮低效品种‘绥玉7号’,从而提高其氮素利用效率,并获得高产。因此,不同氮效率玉米品种氮素的吸收利用差异与氮代谢酶活性的差异密切相关。
前人对不同氮效率玉米品种在氮素的吸收、同化、积累与分配等方面已进行大量研究[15,19],但对不同氮效率玉米品种氮代谢的报道多集中在不同氮肥水平下酶活性的差异,而关于不同氮效率玉米品种响应低氮胁迫的氮代谢与物质生产差异鲜见报道。本研究以前期试验筛选的氮高效品种‘正红311’(ZH311)和氮低效品种‘先玉508’(XY508)为试验材料,研究低氮胁迫下不同氮效率玉米品种氮代谢酶活性、干物质生产及氮素积累的差异,以丰富玉米苗期氮素营养理论,同时为玉米科学施肥提供理论指导与技术支撑。
1 材料与方法
1.1 试验设计
试验于2018年3-5月在重庆市永川区重庆文理学院试验基地进行,采用两因素完全随机设计,不同氮效率玉米品种:氮高效品种‘正红311’和氮低效品种‘先玉508’;不同氮水平:正常氮处理(CK)—Hoagland完全营养液,氮为56 mg·kg-1和低氮处理(LN)—低氮Hoagland营养液,氮为0.7 mg·kg-1。氮以Ca(NO3)2·4H2O供给,并同时以CaCl2·2H2O补充不足的Ca2+。选取大小均匀、籽粒饱满的玉米种子用体积分数为10%的H2O2溶液消毒40 min,用蒸馏水清洗3遍后浸泡在蒸馏水中12 h。然后将种子放在垫有湿润滤纸的培养皿中,在室温黑暗条件下催芽,待种子根长到约1 cm时,将幼苗移到珍珠岩中继续培养。玉米幼苗两叶一心时,去除胚乳移入装有10 L培养液的塑料盆(长50 cm+宽35 cm+高18 cm)中培养(每盆20株),每处理30盆,共120盆。人工气候室昼夜时长为 14 h/10 h,温度为28 ℃/22 ℃。光照时玉米冠层高度的光量子通量密度为250~300 μmol·m-2·s-1。幼苗去除胚乳后在蒸馏水中培养1 d,再转入不同氮质量浓度的营养液中培养。营养液每3 d更换1次,并调节pH为 6.0±0.1,用充气泵每天通气6 h。样品随机摆放,每次更换营养液时相互交换位置。
1.2 指标测定
1.2.1 干物质与氮素积累 于处理后第9 天取样,每处理3次重复,每重复10株,按根系和地上部分开,于105 ℃下杀青30 min,再经80 ℃烘干至恒量后称质量,计算干物质积累与分配情况。称量后将样品粉碎过60目筛,采用凯氏定氮法测定样品氮含量,并计算氮素积累与分配情况。
1.2.2 生理指标测定 于处理后0 h、4 h、12 h、24 h、3 d、6 d和9 d分别取玉米幼苗的根系和叶片,每处理3次重复,每重复5株,分别测定氮代谢酶活性和可溶性蛋白含量。硝酸还原酶(NR)活性采用活体法测定[20],谷氨酰胺合成酶(GS)参照刘淑云等[21]的方法测定,谷氨酸脱氢酶(GDH)参照吕鹏等[22]的方法测定,可溶性蛋白含量采用考马斯亮蓝法[22]测定。
1.2.3 相关计算公式[23-24]根系氮吸收效率(mg·g-1)= 单株氮积累量(mg)/根系干物质(g);根系氮物质生产能力(g·mg-1)= 单株干物质(g)/根系氮积累量(mg)。
1.3 数据分析
采用Excel 2016对数据进行整理并作图,用SPSS 21.0对各性状进行方差分析,方差分析采用最小显著性差异检验法。
2 结果与分析
2.1 低氮胁迫下玉米幼苗氮代谢酶活性差异
低氮胁迫下,玉米幼苗根系和叶片的硝酸还原酶活性均显著下降,各时间点平均,‘正红311’和‘先玉508’根系硝酸还原酶活性分别下降 55.43%和73.24%,叶片硝酸还原酶活性分别下降 64.44%和77.00%(图1)。‘正红311’根系和叶片硝酸还原酶活性降幅均明显低于‘先玉508’,表明氮高效品种‘正红311’较氮低效品种‘先玉508’对低氮胁迫具有更强的响应能力。与‘先玉508’相比,‘正红311’在CK和LN处理下根系和叶片均具有更高的硝酸还原酶活性,各时间点平均,CK处理根系和叶片分别高出16.67%和 8.58%,LN处理分别高出94.33%和 67.91%。低氮胁迫下,两品种根系和叶片的硝酸还原酶活性均在处理12 h时显著增加,但‘正红311’较‘先玉508’根系和叶片的硝酸还原酶活性分别高 143.17%和150.55%,表明氮高效品种‘正红311’根系和叶片的硝酸还原酶较氮低效品种‘先玉508’对低氮胁迫具有更强的响应能力,提高其对低氮环境的适应能力。
低氮胁迫下,玉米幼苗根系和叶片的谷氨酰胺合成酶活性均明显下降,但根系和叶片的降幅存在明显差异(图2)。各时间点平均,‘正红311’和‘先玉508’根系的谷氨酰胺合成酶活性分别下降32.29%和43.33%,而两品种叶片的谷氨酰胺合成酶活性分别仅下降9.79%和18.61%,表明低氮胁迫对玉米幼苗根系谷氨酰胺合成酶活性的影响较叶片更大。氮处理后,两品种根系和叶片的谷氨酰胺合成酶活性均表现出明显的响应,但‘正红311’在CK和LN处理下根系和叶片的谷氨酰胺合成酶活性峰值均显著高于‘先玉508’,CK处理下根系和叶片分别高出17.88%和 62.61%,LN处理下根系和叶片分别高出 84.54%和 104.98%。两品种LN处理下根系和叶片谷氨酰胺合成酶活性明显高于CK处理,表明与氮低效品种‘先玉508’相比,氮高效品种‘正红311’在低氮胁迫下仍能保持较高的谷氨酰胺合成酶活性,提高其对低氮环境的适应能力。
低氮胁迫下,玉米幼苗根系和叶片的谷氨酸脱氢酶活性均显著下降(图3),各时间点平均,‘正红311’和‘先玉508’根系的谷氨酸脱氢酶活性分别下降30.31%和43.80%,叶片的谷氨酸脱氢酶活性分别下降40.05%和51.12%。‘正红311’根系和叶片的谷氨酸脱氢酶活性降幅均明显低于‘先玉508’,表明氮高效品种‘正红311’较氮低效品种‘先玉508’对低氮环境具有更高的适应性。各时间点平均,CK处理下‘正红311’根系和叶片的谷氨酸脱氢酶活性较‘先玉508’分别高出3.94%和5.01%,而LN处理下分别高出 28.88%和28.81%,表明‘正红311’较‘先玉508’的根系和叶片均具有更高的谷氨酸脱氢酶活性。低氮胁迫后,两品种根系和叶片的谷氨酸脱氢酶活性表现为快速增高,但‘正红311’根系和叶片的峰值均显著高于‘先玉508’,分别高出 36.58%和73.63%,表明氮高效品种‘正红311’较氮低效品种‘先玉508’不仅具有更强的氮吸收利用能力,还能快速响应低氮胁迫,提高谷氨酸脱氢酶活性,提高其对低氮环境的适应性。
2.2 低氮胁迫下玉米幼苗的渗透调节物质变化
玉米幼苗根系和叶片的可溶性蛋白含量差异明显,各处理平均,叶片可溶性蛋白较根系高出 175.06%(图4)。与‘先玉508’相比,各时间点平均,CK处理下‘正红311’根系和叶片的可溶性蛋白含量分别高出6.94%和5.46%,而LN处理下分别高出21.13%和25.99%,表明氮高效品种‘正红311’较氮低效品种‘先玉508’具有更强的渗透调节能力,抗逆性更强。低氮胁迫前期(处理24 h内),两品种根系和叶片的可溶性蛋白含量均明显升高,‘正红311’根系和叶片分别升高 20.84%和12.35%,而‘先玉508’分别升高 12.83%和 3.63%;低氮胁迫后期(处理24 h后),两品种根系和叶片的可溶性蛋白含量均明显下降,‘正红311’根系和叶片分别下降23.00%和32.11%,而‘先玉508’分别下降32.02%和 43.17%。氮高效品种‘正红311’根系和叶片的可溶性蛋白含量在低氮胁迫前期增幅明显高于氮低效品种‘先玉508’,而低氮胁迫后期的降幅则明显低于‘先玉508’,使得‘正红311’根系和叶片的可溶性蛋白含量均明显高于‘先玉508’,表明氮高效品种‘正红311’较氮低效品种‘先玉508’具有更强的渗透调节能力,低氮胁迫下优势更 明显。
2.3 低氮胁迫下玉米幼苗的干物质与氮积累
低氮胁迫下,玉米地上部、根系和单株的干质量均显著降低,而根冠比显著升高(表1)。与CK相比,LN处理下‘正红311’和‘先玉508’地上部干质量分别下降51.85%和63.08%,根系干质量分别下降14.33%和13.62%,单株干质量分别下降41.42%和48.44%,根冠比则分别升高 78.05%和133.99%。两品种地上部分干质量降幅均显著高于根系,表明低氮胁迫下玉米幼苗优先满足根系生长,对地上部生长的影响更大。‘先玉508’地上部和单株干质量的降幅与根冠比的增幅均明显高于‘正红311’,表明氮高效品种‘正红311’较氮低效品种‘先玉508’对低氮胁迫具有更强的适应性,尤其是在维持地上部生长上优势更明显。
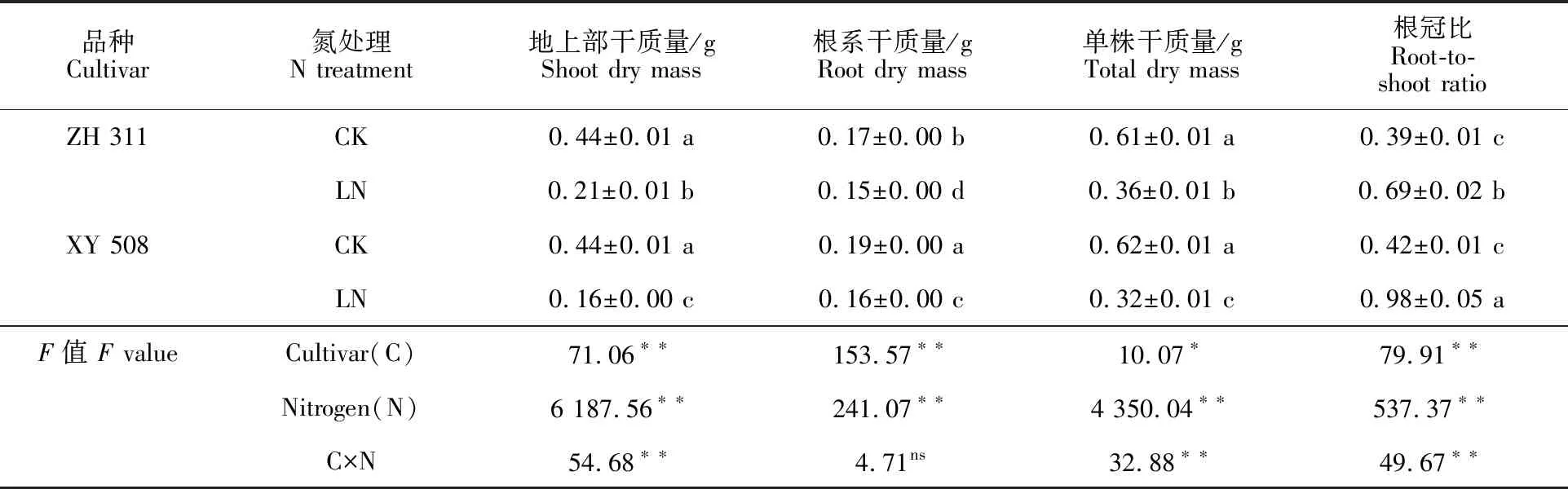
表1 低氮胁迫下不同氮效率玉米品种的干物质积累Table 1 Dry matter accumulation of maize cultivars with different N effiencies under low stress
低氮胁迫下,玉米幼苗地上部、根系和单株的氮积累量均显著降低,而根冠氮分配比显著升高,表明玉米幼苗在氮素受限时优先供给根系(表2)。低氮胁迫下,不同氮效率玉米品种氮素的积累与分配差异显著。‘正红311’地上部和单株的氮积累量较‘先玉508’分别高出41.61%和 21.59%,而根冠氮分配比较‘先玉508’低 31.75%,表明低氮胁迫下氮高效品种‘正红311’较氮低效品种‘先玉508’具有更强的氮素吸收能力,且能更好地协调根系与地上部的氮分配,维持地上部的正常生长,提高其对低氮环境的适应 能力。
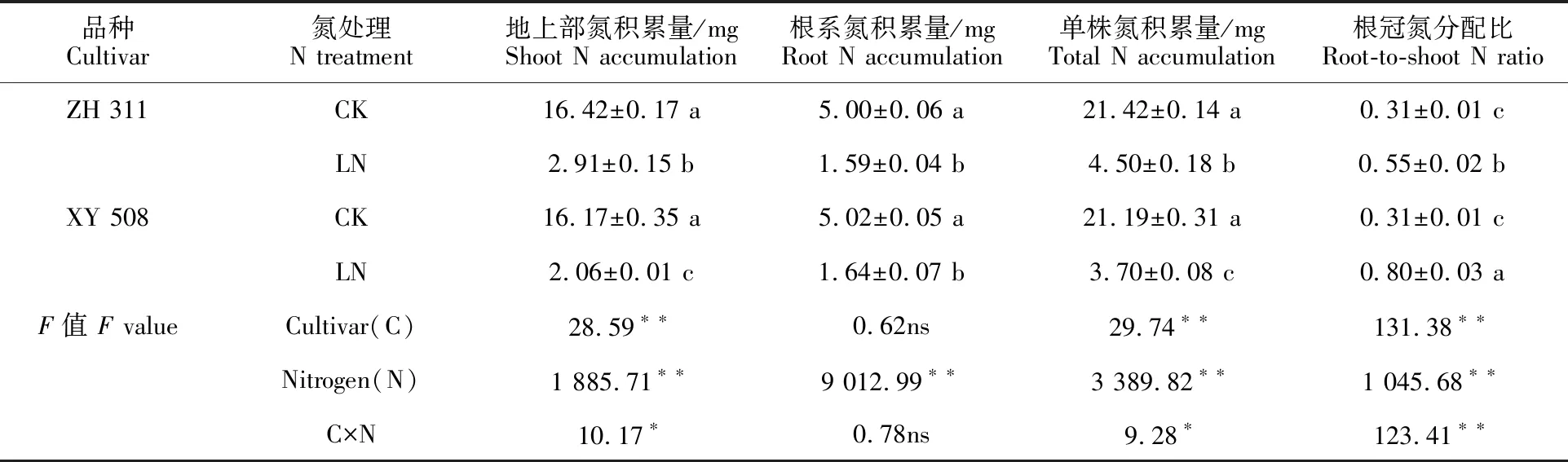
表2 低氮胁迫下不同氮效率玉米品种的氮积累Table 2 N accumulation of maize cultivars with different N effiencies under low stress
2.4 低氮胁迫下玉米幼苗的氮吸收利用差异
‘正红311’两个氮水平下根系的氮吸收效率与物质生产能力均高于‘先玉508’,表明氮高效品种‘正红311’较氮低效品种‘先玉508’具有更强的氮吸收利用能力(图5)。低氮胁迫下,玉米幼苗根系的氮吸收效率显著下降,而根系的氮物质生产能力显著升高。‘正红311’和‘先玉508’的根系氮吸收效率分别降低75.47%和79.76%,根系氮物质生产能力分别升高78.18%和 57.55%,氮高效品种‘正红311’根系氮吸收效率的降幅较氮低效品种‘先玉508’更低,而根系氮物质生产能力的增幅更高,使得‘正红311’根系氮吸收能力和物质生产能力均显著高于‘先玉508’,表明低氮胁迫下‘正红311’较‘先玉508’的氮吸收利用优势更明显。
3 讨 论
3.1 不同氮效率玉米品种氮代谢响应低氮胁迫的差异
氮代谢是植物生长发育过程中最重要的生理生化过程之一[25]。硝酸还原酶是植物同化硝态氮的限速酶,在玉米氮代谢中具有重要地位,谷氨酰胺合成酶和谷氨酸脱氢酶则是植物同化氨的关键酶[20,26]。与氮低效品种‘绥玉7号’相比,氮高效品种‘郑单958’具有更高的硝酸还原酶、亚硝酸还原酶、谷氨酸合成酶和谷氨酰胺合成酶活性,使得其氮素利用效率和产量均较‘绥玉7号’更高[17]。谢孟林等[27]发现,低氮胁迫下玉米根系硝酸还原酶、谷氨酰胺合成酶和谷氨酸脱氢酶活性均显著下降,但氮高效品种的降幅明显低于氮低效品种,使得氮高效品种硝酸还原酶、谷氨酸合成酶和谷氨酸脱氢酶活性高于氮低效品种,提高了氮高效品种对低氮胁迫的适应性。本研究结果表明,低氮胁迫下玉米幼苗根系和叶片硝酸还原酶、谷氨酰胺合成酶和谷氨酸脱氢酶活性均显著下降,但氮高效品种‘正红311’各指标的降幅均低于氮低效品种‘先玉508’,表明氮高效品种‘正红311’较氮低效品种‘先玉 508’对低氮环境具有更强的适应性,这与前人[15,27]研究结果一致。氮高效品种‘正红311’根系和叶片的NR、GS和GDH活性均高于氮低效品种‘先玉508’,且低氮处理下优势更突出,表明旺盛的氮代谢是‘正红311’氮高效与适应低氮胁迫的重要生理机制。两品种根系和叶片的氮代谢活性响应低氮胁迫均表现出大幅增加,但氮高效品种‘正红311’各指标的响应幅度均显著高于氮低效品种‘先玉508’,因此,氮高效品种‘正红311’氮代谢对低氮胁迫的高效响应是其较‘先玉508’更适应低氮胁迫的重要原因。
植物在遭受逆境胁迫时,体内会产生防御机制以适应胁迫环境[28]。可溶性蛋白等有机物,可以提高叶片的渗透调节能力,延缓叶片衰老,为植物有机物的合成提供碳源和氮源[22]。本试验结果表明,低氮胁迫下玉米根系和叶片的可溶性蛋白含量显著下降,但‘正红311’在CK和低氮处理下根系和叶片的可溶性蛋白含量均高于‘先玉508’。低氮胁迫下玉米活性氧清除能力下降,体内膜系统受到伤害,但细胞内保持较高的可溶性蛋白等渗透调节物质,在一定程度保证了细胞渗透势的平衡[22]。氮高效品种‘正红311’较氮低效品种‘先玉508’具有更强的渗透调节能力,是其适应低氮胁迫的重要生理机制之一。
3.2 低氮胁迫下不同氮效率玉米品种物质生产特性差异
物质生产是作物产量形成的基础,低氮胁迫影响植株根系对养分的吸收、转运、同化和利用,进而影响其物质生产,但受影响的程度存在明显的品种差异[25]。玉米在低氮胁迫下干物质积累量、产量与氮积累量均明显下降,但氮低效品种的降幅明显高于氮高效品种[29]。本研究结果表明,低氮胁迫下玉米幼苗根系、地上部和单株干质量均显著下降,而根冠比显著升高。地上部干质量降幅明显高于根系干质量的降幅,表明低氮胁迫对玉米幼苗地上部生长的影响较根系更大。这与卫晓轶等[30]的研究结果一致,低氮胁迫主要是影响玉米苗期地上部的生长。氮高效品种‘正红311’地上部和单株干质量的降幅与根冠比的增幅均明显低于氮低效品种‘先玉508’,这与吴雅薇等[31]的研究结果一致。根冠比显著升高,说明在植株物质积累受限的条件下,将地上部分合成的光合产物优先分配给根系,利于根系维持生长。‘先玉508’根冠比的增幅显著高于‘正红311’,表明氮低效品种‘先玉508’较氮高效品种‘正红311’对低氮胁迫更敏感,受胁迫程度更大,这与刘婷婷等[32]氮高效自交系自330根冠比在缺氮条件下明显高于氮低效自交系陈94-11的结果不一致,主要是因为本研究选取的氮高效品种‘正红311’对氮的高效吸收是通过单位根系更强的氮吸收能力实现的[11],而自330较陈94-11的氮高效是将较多的光合产物分配给根系,从而扩大根系氮吸收面积实现的。
低氮胁迫下,两品种根系的氮吸收效率均显著降低,而根系的氮物质生产效率均显著升高。‘正红311’根系的氮吸收效率在CK和低氮处理下均显著高于‘先玉508’,进一步说明氮高效品种‘正红311’较氮低效品种‘先玉508’的氮高效体现在单位根系的氮吸收能力上。低氮胁迫下,‘先玉311’根系氮物质生产能力显著高于‘先玉508’,表明氮高效品种‘正红311’较氮低效品种‘先玉508’在缺氮时能够更好地协调地上部与根系平衡,维持地上部的正产生长,保证光合产物的供给。与氮低效品种‘先玉 508’相比,氮高效品种‘正红311’保持较高的根系氮吸收效率与物质生产能力是其适应低氮胁迫的重要机制。
4 结 论
低氮胁迫下,玉米幼苗根系和叶片的硝酸还原酶、谷氨酸合成酶、谷氨酸脱氢酶活性,可溶性蛋白含量,地上部、根系和单株干质量与氮积累量及根系氮吸收效率均显著下降,但‘正红311’各指标降幅均低于‘先玉508’;根冠比、根冠氮分配比和根系氮物质生产力均显著升高,但‘正红311’根冠比、根冠氮分配比的增幅均低于‘先玉508’,而根系氮物质生产力的增幅较‘先玉508’更高。氮高效品种‘正红311’的硝酸还原酶、谷氨酸合成酶、谷氨酸脱氢酶活性,可溶性蛋白含量均高于氮低效品种‘先玉508’,且对低氮胁迫响应更高效,使得其根系的氮吸收效率和物质生产能力显著高于‘先玉508’,而根冠比和根冠氮分配比例显著低于‘先玉508’,有效地维持地上部与根系干物质和氮营养的协调分配,保证其物质生产与氮素积累,提高了其对低氮环境的适应能力。因此,肥沃地区种植氮高效品种可适当减少氮肥用量,充分发挥其氮高效潜力,而丘陵地区推广氮高效品种,则有利于保证玉米的高产稳产。
