高温和强光复合胁迫对铁皮石斛光系统I和II活性的影响*
2021-06-25黄伟谭顺玲孙虎
黄伟,谭顺玲,孙虎
(1.中国科学院昆明植物研究所 资源植物与生物技术重点实验室,云南 昆明 650201;2.中国科学院昆明植物研究所 植物医生研发中心,云南 昆明 650201)
植物利用光能进行光合作用,通过光反应产生的ATP(腺嘌呤核苷三磷酸)和NADPH(还原型烟酰胺腺嘌呤二核苷酸磷酸)在暗反应中被用于固定二氧化碳(CO2)并转化为合成有机物。光反应中的光合电子传递分为线性电子传递、围绕光系统I(PSI)的环式电子传递、假环式电子传递等[1]。光合电子传递的运转形成跨类囊体膜质子驱动势,驱动ATP合成酶合成ATP[2]。线性电子传递产生的ATP/NADPH比值大约为1.29,暗反应所需的ATP/NADPH比值大概为1.6,所以植物需要其他的替代电子传递途径来补充ATP的合成[3]。环式电子传递产生的跨类囊体膜质子梯度不仅补充ATP的合成,并且通过调节光合电子传递链的氧化还原态保护PSI和光系统II(PSII)免受过剩光能的损伤[4]。非生物胁迫对PSII活性的影响主要集中在单因子胁迫(高温、低温、强光、干旱等)[5-8],但是目前关于植物对高温和强光复合胁迫的光合生理响应还了解较少。
中等程度高温会对PSII的放氧复合体活性造成损伤[9],进而引起PSII的光抑制[10]。与此同时,高温导致气孔关闭,抑制卡尔文循环,增加叶绿体内活性氧自由基的产生[11]。活性氧自由基会抑制D1蛋白的合成,进而影响PSII活性的快速修复[12-13]。所以,高温胁迫不仅会提高损伤速率,还会降低修复速率[14]。在强光条件下,过剩光能大部分通过非光化学淬灭以无害的形式耗散(热耗散),避免PSII的修复受到抑制[7,15-16]。热耗散的激发依赖于跨类囊体膜质子梯度(ΔpH)的建立,而ΔpH的建立又依赖线性电子传递和环式电子传递[17],光合电子传递的运转受PSI和PSII活性的控制[18-20]。所以,在高温和强光复合胁迫下,PSII的损伤有可能抑制线性电子传递并影响跨类囊体膜质子梯度的建立,从而加剧光抑制的发生。
高温胁迫也会对PSI的活性造成影响,研究表明42 ℃的中等程度高温和300 μmol/(m2·s1)光照处理10 h就会对小麦(Triticumaestivum)的PSI活性造成严重损伤,且损伤程度要高于PSII[5]。这主要是高温加剧了类囊体膜的质子渗漏,降低了ΔpH[21],进而造成PSI的电子载体处于过度还原态,引发PSI内活性氧自由基的产生,造成PSI受损。此外,42 ℃的中等程度高温会加剧波动光强下烟草(Nicotianatabacum)幼叶的PSI损伤程度[22]。但是,中等程度高温和持续恒定强光对PSI活性的影响还缺乏研究。
铁皮石斛(DendrobiumofficinaleKimura et Migo)是一种有重要药用价值的兰科(Orchidaceae)植物,在亚热带地区被广泛种植。但是,目前关于逆境胁迫对铁皮石斛光合机构及光合电子传递影响的研究还十分欠缺。由于高温和强光复合胁迫在铁皮石斛种植地区的夏季经常出现,因此本研究将以铁皮石斛“昆植1号”品种为研究对象,研究高温和强光复合胁迫对其PSI和PSII的影响及其光合生理机制,为铁皮石斛的种植策略提供参考。
1 材料与方法
1.1 研究材料
供试材料为种植在中国科学院昆明植物研究所温室内的铁皮石斛品种“昆植1号”。铁皮石斛“昆植1号”植株被栽培在直径为20 cm的透水塑料盆中,以树皮为基质。生长季昼夜温度为20/30 ℃,空气湿度维持在60%左右,光照条件为全光照的40%。
1.2 实验设计
将铁皮石斛植株在常温下暗适应30 min后,放入42 ℃温室中继续暗适应5 min,然后利用Dual-PAM 100双通道叶绿素荧光仪(德国Walz公司)测定叶绿素荧光快速动力学曲线(300 ms)和Pm值,分析处理前叶片的PSII和PSI活性。随后暗适应5 min,然后测定强光〔1 809 μmol/(m2· s)〕处理过程中PSI和PSII参数的变化。处理60 min后,再暗适应30 min测定叶绿素荧光快速动力学曲线和Pm值,分析处理后叶片的PSII和PSI活性。通过比较处理前后PSII和PSI活性的差异,判断高温强光处理对PSII和PSI活性的损伤程度。
1.3 指标测定
PSI和PSII参数的测定都利用Dual-PAM 100双通道叶绿素荧光仪(德国Walz公司)完成。叶绿素荧光快速动力学曲线的测定使用20 000 μmol/(m2· s)的强光脉冲(300 ms),叶绿素相对荧光强度的快速动力学曲线利用最大荧光强度进行标准化。PSII的参数计算如下[23]:
Y(II)=(Fm′ -Fs)/Fm′;NPQ=(Fm-Fm′)/Fm′;Y(NO)=Fs/Fm。
式中:Fm代表暗适应后的最大叶绿素荧光强度;Fm′代表光适应后的最大叶绿素荧光强度;Fs代表光适应后的稳态荧光强度;Y(II)表示PSII的有效光化学量子产额;NPQ表示PSII的非光化学淬灭能力;Y(NO)表示PSII的非调节性热耗散量子产额。PSI的参数计算如下[22]:
Y(I)=(Pm′-P)/Pm;Y(ND)=P/Pm;Y(NA)=(Pm-Pm′)/Pm。
式中:Y(I)代表光PSI的光化学量子产额,Y(ND)代表PSI反应中心P700的氧化态比例,Y(NA)代表PSI反应中心P700的还原态比例。光合电子传递速率的计算如下:
ETRI = PPFD×Y(I)×0.84×0.5;ETRII = PPFD×Y(II)×0.84×0.5。
式中:ETRI和ETRII分别代表通过PSI和PSII的电子传递速率,ETRI-ETRII代表环式电子传递速率[24],PPFD代表光合有效辐射强度。
1.4 数据处理
本研究的每组实验测定包括5个重复,数据为平均值±标准误。采用SPSS 16.0软件对数据进行统计分析。处理前后的数据比较采用独立样本T检验进行。
2 结果与分析
2.1 高温强光胁迫对叶绿素快速动力学曲线的影响
高温强光胁迫对铁皮石斛叶绿素影响见图1。

图1 高温强光处理对铁皮石斛叶绿素荧光快速动力学曲线的影响
在42 ℃的中等高温胁迫下对铁皮石斛叶片进行强光〔1 809 μmol/(m2· s〕处理60 min后,测定其叶绿素荧光快速动力学曲线(OJIP曲线),发现J点、I点和P点的叶绿素荧光强度都发生大幅度下降(图1A),表明PSII活性受到了严重损伤。此外,在300 μs时的相对叶绿素荧光强度(K点)发生了显著上升(图1B),表明PSII的电子供体即放氧复合体活性受到了明显的损伤。
2.2 高温强光胁迫对PSII光量子产额的影响
高温强光胁迫对铁皮石斛PSII光量子产额的影响见图2。

图2 高温强光处理对铁皮石斛光系统II量子产额的影响
在暗适应叶片突然曝光的前1 min内,铁皮石斛的PSII光化学量子产额[Y(II)]维持在很高水平(图2A),这主要和水-水循环介导的PSII电子传递相关。在高温和强光处理过程中,Y(II)在前6 min内大幅度下降(图2A),说明铁皮石斛的卡尔文循环受到了高温胁迫的强烈抑制,导致线性电子传递产生的ATP和NADPH不能被及时消耗,NADP+/NADPH的比例下降,影响了电子从PSII经过PSI再传递给NADP+的过程。在高温和强光处理60 min后,铁皮石斛的NPQ从2.4下降到0.43,下降幅度达到了82%(图2B)。NPQ的激发不仅依赖跨类囊体膜质子梯度,同时还受PSII复合体核心活性的影响,前者和后者对NPQ的贡献分别大概为60%和40%。所以,铁皮石斛经高温和强光处理后NPQ的大幅度下降不仅和跨类囊体膜质子梯度的不足有关,同时还涉及PSII反应中心的受损。当NPQ能力不足时,PSII吸收的大部分光能就变成了过剩光能,引起Y(NO)的大幅度上升(图2C)。Y(NO)的大幅度上升会引起活性氧自由基在PSII复合体处大量产生,进而影响D1蛋白的合成和PSII的修复过程。
2.3 高温强光胁迫对PSI量子产额的影响
高温强光胁迫对铁皮石斛PSI光量子产额的影响见图3。

图3 高温强光处理对铁皮石斛光系统I量子产额的影响
与Y(II)类似,在暗适应叶片突然曝光的前1 min内,铁皮石斛的PSI光化学量子产额[Y(I)]维持在很高水平(图3A),这主要和水-水循环及环式电子传递的高度激发有关。在高温和强光处理过程中,Y(I)在前6 min内大幅度下降,随后缓慢下降(图3A)。PSI的反应中心在暗适应突然曝光后的20 s就高度氧化(图3B),说明水-水循环在高温处理的初期耗散了积累在PSI的过剩活化电子,避免其出现过度还原态(图3C)。但是,在随后的高温强光处理过程中,PSI的氧化态比例逐渐下降(图3B),过度还原态比例逐渐上升(图3C),这主要和PSI的逐渐受损相关。
2.4 高温强光胁迫对光合电子传递的影响
高温强光胁迫对铁皮石斛光合电子传递的影响见图4。
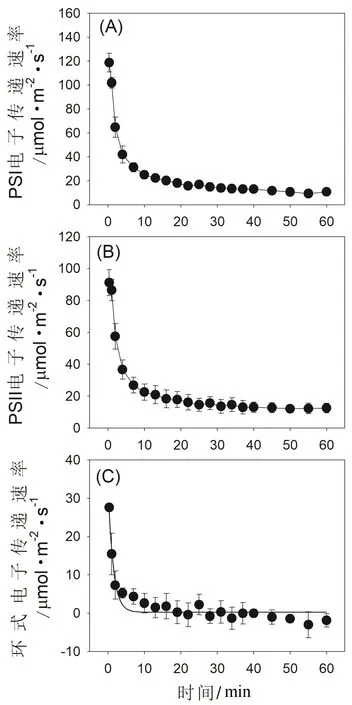
图4 高温强光胁迫对铁皮石斛光合电子传递速率的影响
在暗适应叶片突然曝光的前1 min内,PSI电子传递速率(ETRI)维持在很高水平(图4A),这主要是因为水-水循环和环式电子传递都高度激发。在随后的处理过程中,水-水循环介导的PSII电子传递大幅度下降(图4B),并且环式电子传递的激发受到抑制(图4C),导致ETRI快速下降。值得注意的是,在高温强光处理5 min以后,环式电子传递的活性基本丧失(图4C),这会严重影响跨类囊体膜质子梯度的构建,进而抑制NPQ的激发。
2.5 高温强光胁迫对PSI和PSII活性的影响
经过60 min高温和强光处理后,最大叶绿素荧光强度(Fm)下降了78%,表明PSII活性受到严重损伤。与此同时,最大可氧化的P700值(Pm)下降了31%,说明PSI活性也出现了中等程度损伤。相比较PSII活性而言,PSI活性的受损程度较低,这说明PSII是高温强光胁迫对铁皮石斛造成光损伤的主要靶点。

图5 高温强光胁迫对铁皮石斛光系统I和II活性的影响
3 讨论与结论
3.1 讨论
自然条件下,高温和强光胁迫是植物在夏季经常遭遇的非生物逆境胁迫,但这种复合胁迫对植物叶片PSI和PSII的影响还有待揭示。有研究表明,42 ℃和中等光强复合胁迫对小麦PSI活性的影响大于其对PSII活性的影响。本研究发现,在42 ℃下强光胁迫处理60 min后铁皮石斛的最大叶绿素荧光值(Fm)下降了78%,表明其PSII活性受到严重损伤[20]。与此同时,其最大可氧化的P700值(Pm)下降了31%,表明PSI活性也有显著的下降[22,25-26]。所以,高温和强光胁迫对铁皮石斛PSII活性的影响大于其对PSI活性的影响。此外,通过PSI和PSII的电子传递速率分别下降91%和86%,说明PSI和PSII的损伤对整个光合电子传递链的功能造成了严重影响。
PSII的损伤程度由损伤速率和修复速率共同决定,当损伤速率大于修复速率时,PSII活性发生损伤[27]。反之,当修复速率大于损伤速率时,PSII活性得到修复[28]。在自然条件下,强光导致的PSII损伤会在弱光条件下修复[29]。PSII的损伤速率主要和光照强度相关,但修复速率会受到其他环境因素的影响,例如盐胁迫、温度胁迫等[10]。本研究发现,高温和强光复合胁迫处理对铁皮石斛的放氧复合体活性造成显著性损伤,这和PSII损伤的“两步模型”相吻合。在“两步模型”中,放氧复合体是PSII损伤的最初靶点,放氧复合体的受损随后引起PSII的损伤[30]。在逆境条件下,叶片的光能利用效率下降,导致活性氧自由基的含量上升。活性氧自由基会抑制PSII复合体关键亚基D1蛋白的重新合成,进而影响D1蛋白和受损PSII复合体的组装,限制PSII活性的修复。所以,环境胁迫对PSII修复速率的影响主要和活性氧自由基的产生密切相关[10]。当暗适应叶片在42 ℃下突然曝光后,其PSII电子传递速率(ETR II)先保持在较高水平,然后急速下降,这主要和水-水循环电子传递的短暂激发相关,此时水-水循环电子传递的运转避免了PSI处于过度还原态,也消耗了PSII的部分过剩光能。但是,在随后的高温强光处理过程中,ETR II逐渐下降并维持在较低的水平,说明铁皮石斛的暗反应受到限制。高温容易引起卡尔文循环的受限,导致NADPH的消耗速率变慢,引起NADP+的缺失,进而造成电子从PSI到NADP+的传递受阻,最终导致ETRII受限,引发活性氧自由基的大量产生,从而抑制PSII活性的修复。此外,非光化学淬灭(NPQ)作为PSII耗散过剩光能的主要保护机制,在高温强光胁迫处理60 min后大幅度下降,这也加剧了PSII内部活性氧自由基的产生。所以,在42 ℃和强光复合胁迫条件下,较高的PSII损伤速率和较低的PSII修复速率协同作用,导致铁皮石斛的PSII活性严重受损。
PSII活性虽然容易受损,但是在合适条件下只需要几个小时就能够基本修复[31]。而PSI活性则不同,一旦受损就需要一周甚至更长时间才能完全修复[18,32]。PSI的损伤主要是由于其反应中心附近的电子载体处于过度还原态,造成PSI复合体内部产生大量的活性氧自由基,进而引发氧化损伤[33]。在正常生长条件下,叶片产生较高的跨类囊体膜质子梯度(ΔpH),在Cyt b6f复合体处调节PSII到PSI的电子传递,保证PSI的电子载体在强光条件下都处于高度氧化态,进而避免过度还原引起的光损伤[34-35]。本研究发现,铁皮石斛的PSI电子载体在高温强光胁迫处理的初期并没有发生过度还原,但是在处理后期PSI电子载体出现过度还原并伴随着光损伤。ΔpH在调控PSI氧化还原态过程中发挥重要作用,例如在拟南芥(Arabidopsisthaliana)环式电子传递突变体pgr5中,强光条件下ΔpH的建立不足导致其PSI发生严重损伤,甚至造成植株死亡[36-39]。所以,环式电子传递是植物在强光条件下建立充分的ΔpH并保护PSI活性的重要替代电子传递途径[1,40]。本研究还发现,在高温强光胁迫处理的初期,铁皮石斛的环式电子传递被高度激发。但是在处理后期,环式电子传递的激发程度很低,这就导致ΔpH的建立不足。同时,由于NPQ的激发也依赖ΔpH的建立,因此高温强光处理后NPQ的下降也证实了ΔpH的建立受限。所以,长期的高温强光胁迫会导致环式电子传递失活,引起ΔpH的建立不足,进而引起PSI活性受损。
3.2 结论
高温胁迫对光系统II活性及光量子产额的影响是导致其光合作用机构受损的重要方面[41-44]。本研究发现,在高温强光复合胁迫处理60 min后,铁皮石斛的PSI和PSII活性都显著下降,且PSII的下降幅度更大,表明PSII活性的受损是铁皮石斛难以适应长期高温强光复合胁迫的重要原因。提高光能利用效率是维持植物生长和提高产量的基础,而负责光反应的PSI和PSII活性是决定光能利用效率的重要因素。鉴于高温强光胁迫会对铁皮石斛的PSI和PSII活性造成严重损伤,在铁皮石斛种植条件的选择和控制上,应尽量避免高温和强光复合胁迫。本研究的设计思路和实验结果对于其他农作物和经济作物种植策略的选择同样具有参考和借鉴意义。
