冰川退缩迹地植被原生演替初期关键种的碳氮磷生态化学计量特征
2021-06-25庄文化李明明史忠林
周 萍, 庄文化, 李明明, 龙 翼, 史忠林, 周 继
(1.中国科学院、水利部 成都山地灾害与环境研究所, 山地表生过程与生态调控重点实验室, 四川 成都 610041; 2.四川大学 水利水电学院, 四川 成都 610065; 3.四川大学水力学与山区河流开发保护国家重点实验室, 四川 成都 610065; 4.中国科学院大学, 北京 100049)
生态化学计量学是研究生态系统营养元素分配、循环过程和限制性元素判断的一种新兴手段[1],主要用于研究不同生态系统、不同组织水平、不同有机体内部能量平衡、养分循环和化学元素(主要是C,N,P)的动态平衡[1]。生态化学计量特征不仅反映了养分状况,同时还能揭示养分的可获得性及其限制因子[2],这对探讨植物养分循环与分配的调控机制、揭示元素平衡、分析生态要素交互作用具有重要意义[1]。高山或极地冰川消退区所形成的冰川裸地作为研究植被原生演替的理想场所,引起了生态学家的广泛关注[3]。贡嘎山海螺沟地区第四纪冰川广泛发育,下伸到谷地,现代冰川相当发育[4]。松散固体物质储量巨大,受全球、局地气候变化(温度升高,暴雨增多或减少)、地震等,冰川末端逐渐退缩,冰川退缩后形成了大量原生裸地。由于此处受到的人为干扰少,随着时间的推移,在原来冰川退缩迹地上重新开始了植物的原生演替,演替序列具有连续性和完整性,有利于研究冰川退缩区植被演替动态[5]。冰川退缩形成的原生裸地在第4 a就有被子植物生长发育,在第5—6 a的冰川退缩迹地就会形成以柳叶菜(Epilobiumhirsutum)和马河山黄芪(Astragalusmahoschanicus)为关键种的群落,最初群落的植物比较稀疏,随着马河山黄芪固氮作用的加强,改善了局部生境,在第6—8 a,马河山黄芪取代柳叶菜成为建群种,再经过3~8 a,沙棘(Hippophaerhamnoides)取代马河山黄芪成为建群种,冰川退缩迹地朝着草本向灌丛的演替进行。植物对生境的适应是其存活和定居的关键,碳氮磷元素是植物生存与生长发育必需的结构元素[6]。植物不同器官的碳氮磷及其生态化学计量特征的研究有助于解释植物碳氮磷元素在不同器官内的分配规律及植物为适应环境对自身养分的调控能力[7]。近年来,国内外学者对生态系统的化学计量特征研究主要集中在沙地[8]、湿地[9]、荒漠等[10]不同生态系统,但涉及冰川退缩迹地植物C,N,P生态化学计量及植物各器官碳氮磷化学计量耦合等的研究较缺乏[11]。为此,本研究基于对贡嘎山海螺沟冰川退缩迹地植被演替序列初期的两种关键种的调查,选取了演替初期的柳叶菜和马河山黄芪2个关键种,分别代表冰川退缩迹地植被演替初期第5 a和第8 a的演替群落,探讨了演替初期植被不同器官的C,N,P分配特征及化学计量比的变化。并从生态化学计量特征的角度分析不同器官间C,N,P之间的相关关系,这对探索冰川退缩迹地植被演替初期关键物种的个体生长、种群动态和限制元素的判断,预测指示植被的演替进程及从一定程度上揭示演替早期关键种的侵入和维持机制提供科学依据和数据支持。
1 研究区概况与研究方法
1.1 研究区概况
海螺沟是贡嘎山东坡最主要冰川河,也是我国最具代表性的季风海洋性山谷冰川之一,水热条件好,冰川消融速度快,自小冰期开始退缩[12],近百年来没有冰进过程,土壤有连续成土过程。20世纪30年代退缩加速形成了贡嘎山东坡海螺沟冰川退缩区(29°3 421″N,102°5 942″E),在长达2 km的范围内形成了一个完整的从裸地到先锋群落再到顶级群落的连续植被原生演替序列。该区主要受东亚季风影响,该地区气候湿冷,属于山地寒温带气候类型,年平均气温3.8 ℃,1月平均气温最低(-4.38 ℃),7月平均气温最高(11.9 ℃),年降水日数在260 d以上,年平均降水量约1 960 mm,年平均相对湿度约90%。
1.2 研究方法
1.2.1 样品采集 本研究以120多年形成的贡嘎山海螺沟冰川退缩区的末端作为研究区,基于前人对海螺沟冰川退缩区植被演替不同阶段的划分[11],采用以空间代时间的方法,选取了冰川退缩迹地植被原生演替第5年的柳叶菜群落和第8年的马河山黄芪群落2个典型样地为主要研究区域,于2019年6—10月在2个样地分别随机布设5个1 m×1 m样方,调查样方内所有草本和地被物,记录其种类、数量等,并采集植物根系、茎、叶片、果实样品带回实验室进行预处理。
1.2.2 植物不同器官CNP测定 样方内选取生长健壮、长势良好的柳叶菜和马河山黄芪植株,从上中下3个层次分别采集每株植物成熟且无病虫害叶片、茎样品混匀后装入信封中。在成熟季节分别采集柳叶菜和马河山黄芪的果实样品,并挖取植株完整根系冲洗后装入信封,做好标记并带回实验室。将带回实验室的植物样品置于恒温干燥箱中在105 ℃下杀青0.5 h,然后在80 ℃下烘干到质量恒定,再粉碎并过100目(孔径为0.15 mm)筛后保存用于CNP测定。叶片、茎、果实、根系有机碳采用重铬酸钾外加热法测定,TN采用凯氏定氮法测定,TP采用高氯酸—硫酸消解钼锑抗比色法测定[13]。
1.2.3 数据处理 使用SPSS 18.0软件对数据进行统计分析,采用单因素方差分析法(one-way ANOVA)分析冰川退缩迹地演替初期柳叶菜和马河山黄芪不同器官的C,N,P含量以及元素生态化学计量比(C/N,C /P,N/P)特征,并运用Excel作图。
2 结果与分析
2.1 冰川退缩迹地植被演替初期两种关键种不同器官碳氮磷含量特征
碳(C)、氮(N)、磷(P)作为组成植物体的基本元素,在植物生长发育及其各项生理活动协调中发挥着重要作用。植物不同器官的碳氮磷存在相关关系,C是植物体干物质的组成元素[14],N是蛋白质的重要组成部分[15],P与细胞结构和DNA,RNA的组成密切相关[1]。冰川退缩迹地演替初期两种关键种柳叶菜和马河山黄芪不同器官C,N,P含量可反映出该植物对其生长环境的适应特征。柳叶菜根系有机碳含量变化范围为436~445 g/kg,较马河山黄芪根系有机碳含量(416~423 g/kg)高(图1)。叶片的C含量主要来源于土壤、根系和地上部分器官的输移能力[16],冰川退缩迹地演替初期,退缩迹地土壤养分贫瘠,适宜物种较少,每年的地表生物量及枯落物归还土壤的量均较少。柳叶菜和马河山黄芪根系全氮含量表现出与有机碳相反趋势,由于马河山黄芪为豆科固氮植物,促进了根系和地上部分对N素的吸收利用[16],因此马河山黄芪根系全氮含量显著高于柳叶菜根系全氮含量(p<0.05)。柳叶菜和马河山黄芪根系全磷含量较低,且未达到显著性水平,这主要是由于土壤中的P是一种沉积性元素,受成土母质、气候等影响,主要来源于岩石风化[17],冰川退缩迹地演替初期岩石风化程度低。柳叶菜和马河山黄芪茎的有机碳含量表现为:柳叶菜>马河山黄芪,且差异显著(p<0.05)。而茎全氮和全磷含量均表现为:柳叶菜<马河山黄芪,且差异显著(p<0.05)。叶片是植物代谢最活跃的器官,它的养分特征对环境响应的敏感性较高。柳叶菜和马河山黄芪叶片有机碳含量(311~394 g/kg)和全磷含量(1.49~1.78 g/kg)间的差异均未达到显著水平。柳叶菜叶片全氮含量显著低于马河山黄芪叶片的全氮含量(p<0.05)。柳叶菜和马河山黄芪果实的有机碳含量显著高于马河山黄芪果实的有机碳含量(p<0.05),果实的全氮含量则表现为柳叶菜<马河山黄芪,且达到显著性差异(p<0.05)。两个关键种果实的全磷差异未达到显著性水平。
柳叶菜不同器官的有机碳含量变化为:根系>果实>叶片>茎,柳叶菜叶片的全氮含量最高,其次为根系、果实和茎,柳叶菜全磷含量表现为:果实>叶片>根系>茎。马河山黄芪不同器官的有机碳含量变化为:黄芪根系>果实>茎>叶片。马河山黄芪全氮含量最高的器官为叶片,其次为根系、果实和茎,全磷含量变化为:叶片>果实>根系>茎。柳叶菜和马河山黄芪不同器官的C,N,P平均值均低于全球平均水平和我国平均水平[18],但2个关键种不同器官的CNP分配策略相似,均优先保障果实和叶片的NPK含量以繁衍后代和维持自身生长代谢,不断推动冰川退缩迹地植被演替的进行。

注:不同小写字母表示在0.05水平差异显著。下同。
2.2 冰川退缩迹地植被演替初期两种关键种不同器官生态化学计量比特征
冰川退缩迹地植被演替初期的柳叶菜和马河山黄芪两种关键种的根系C/N,叶片C/N和果实C/N之间存在显著差异(p<0.05),2种关键种的茎C/N之间差异不显著(图2)。柳叶菜不同器官的C/N高于马河山黄芪,C/N高说明植物固碳效率较高,柳叶菜较马河山黄芪处于演替的更早期,因此柳叶菜不同器官对碳的固定效率提高。柳叶菜和马河山黄芪两种关键种不同器官的N/P变化范围为5.94~17.82,也表现出根系N/P,叶片N/P和果实N/P之间存在显著差异(p<0.05),柳叶菜和马河山黄芪的茎N/P之间差异不显著。C/P可表征植物磷素的利用效率与碳同化能力,柳叶菜和马河山黄芪根系C/P和茎C/P之间差异不显著,而两种关键种的叶片和果实C/P比存在显著差异(p<0.05)。

图2 柳叶菜和马合山黄芪不同器官生态化学计量比特征
2.3 冰川退缩迹地植被演替初期两种关键种不同器官碳氮磷及生态化学计量比间相关性分析
冰川退缩迹地植被演替初期柳叶菜根系C含量与茎N含量、果实C含量之间呈现极显著正相关(p<0.01)(表1),柳叶菜根系P含量与根系C含量、叶片P含量及果实P含量间显著正相关(p<0.05),柳叶菜茎C含量与叶片P含量间极显著正相关(p<0.01),柳叶菜茎N含量与果实C含量间极显著负相关(p<0.01),柳叶菜叶片C含量与叶片N含量间极显著正相关(p<0.01)。

表1 柳叶菜不同器官C,N,P间相关性
冰川退缩迹地植被演替初期柳叶菜根系C/N与茎C/N、叶片C/N间存在显著正相关关系(p<0.05)(表2),柳叶菜根系N/P与叶片N/P间,根系C/P与茎C/P间存在显著负相关关系(p<0.05),根系C/P与叶片C/P间存在极显著正相关关系(p<0.01),根系C/P与果实N/P间显著正相关(p<0.05)。柳叶菜茎N/P与茎C/P间极显著正相关(p<0.01),柳叶菜茎N/P与叶C/P,茎N/P与果实N/P间显著负相关(p<0.05)。柳叶菜茎C/P与叶片C/P间显著负相关,茎C/P与果实N/P间极显著负相关(p<0.01)。柳叶菜果实N/P与叶片N/P,果实N/P与果实C/P间存在显著正相关关系(p<0.01)。

表2 柳叶菜不同器官生态化学计量比间相关性
冰川退缩迹地植被演替初期马河山黄芪不同器官的C,N,P含量相关性分析结果表明(表3),马河山黄芪不同器官的C,N,P含量间相关性较低,马河山黄芪茎P含量与叶片P含量间存在极显著正相关关系(p<0.01),叶片P含量与果实N含量间存在显著正相关关系(p<0.05)。
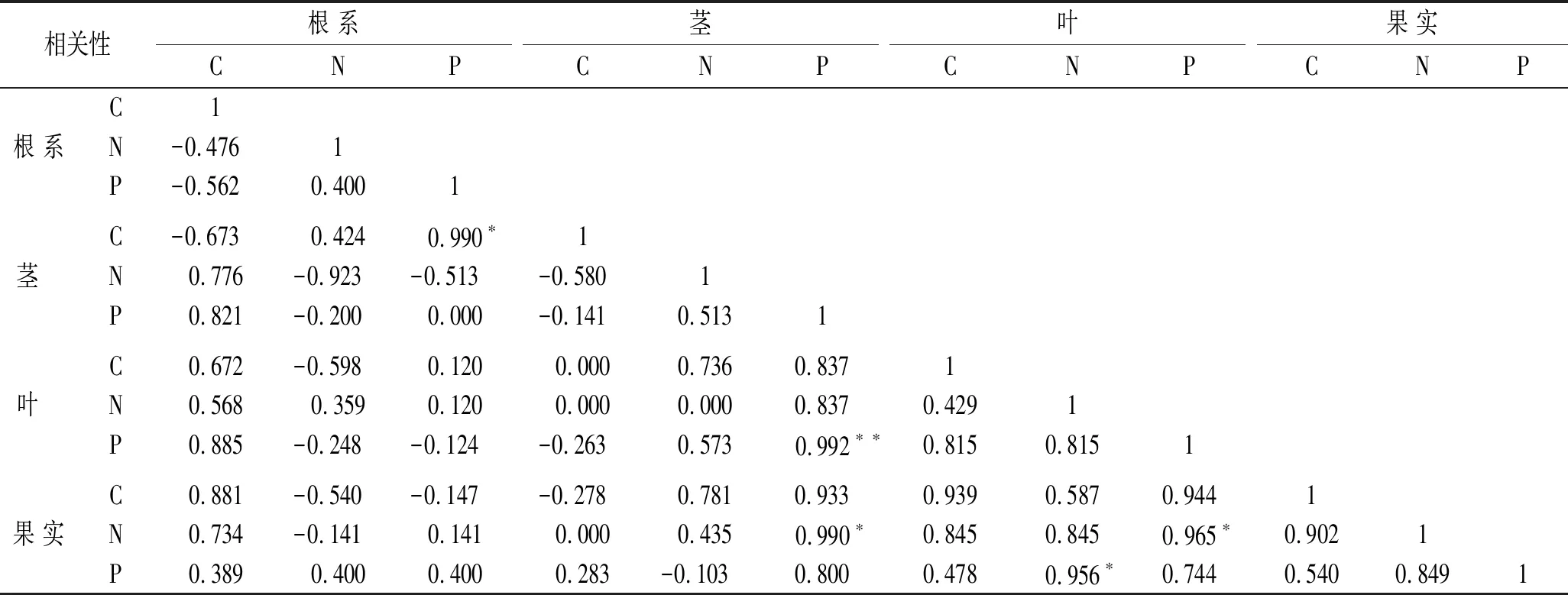
表3 马河山黄芪不同器官C,N,P间相关性
冰川退缩迹地植被演替初期马河山黄芪根系C/N与叶片C/P间存在显著正相关关系(p<0.05)(表4),茎N/P与茎C/P间显著负相关(p<0.05),茎N/P与茎C/N间极显著正相关(p<0.01),茎N/P与叶N/P间存在显著正相关关系(p<0.05)。马河山黄芪茎C/P与茎C/N,茎C/P与果实C/N间存在显著负相关关系(p<0.05),叶片C/N与叶片N/P间存在显著正相关关系(p<0.05)。

表4 马河山黄芪不同器官生态化学计量比间相关性分析
3 讨 论
3.1 冰川退缩迹地植被演替初期关键种的C,N,P元素特征
海螺沟冰川属于季风海洋性冰川,与欧洲、北美等处于高纬度的冰川相比,海螺沟冰川退缩区具有气温高、降水量大的特点,水热条件较好,为早期先锋植物的定居提供了良好的环境基础。Chapin等[19]通过对阿拉斯加Glacier Bay冰川原生演替研究发现,先锋植物在冰川退缩第10 a后才开始侵入原生裸地,且最初形成的先锋群落没有乔木幼苗成分,到15—20 a才有柳树和赤杨(Alnusrubra)进入群落。而贡嘎山海螺沟冰川退缩迹地,在第4 a就有种子植物黄芪、柳叶菜等生长发育,随着演替的进行有乔灌木幼苗伴生其中,植被演替速度加快。
冰川退缩迹地植被演替初期,柳叶菜和马河山黄芪常作为先锋物种快速侵入,随着演替的推进,马河山黄芪的固氮作用不断得到发挥,在演替到第6年,柳叶菜群落优势地位逐渐被马河山黄芪群落替代。植物不同器官的C,N,P含量及差异反映了植物对元素的吸收、运输和利用特征[20]。叶片与根系是植物地上部分和地下部分重要的两个营养器官,叶片是植物进行光合作用和制造有机物的重要器官[21],根系是植物体吸收、存储、运输养分水分[22]的主要器官。对比分析植物不同器官的C,N,P含量及其相关性有助于深入理解植物不同器官对C,N,P等营养物质的利用与分配规律[23]。冰川退缩迹地演替初期的柳叶菜和马河山黄芪两种关键种均为多年生草本植物,相比于一年生草本植物,多年生草本植物通常具有较长的叶寿命和较高的养分含量[24]。本研究的柳叶菜和马河山黄芪不同器官C含量平均值(409.63 g/kg)均低于全球植物C平均值[25],表明该区域植物不同器官C含量较低。柳叶菜不同器官N含量值变化为7.48~12.98 g/kg,表现为:叶片>果实>根系>茎,低于我国[19]和全球[26]植物叶片N含量。这可能是由于柳叶菜处于冰川退缩迹地演替初期,退缩迹地土壤养分较贫瘠有关。柳叶菜不同器官P含量均值为0.81~3.52 g/kg,与我国和全球尺度植物叶片P含量的研究较为接近[19]。这可能是由于柳叶菜不同器官的P元素主要来源于退缩迹地的母岩风化,由于冰川退缩迹地成土母质风化速度慢,使得冰川退缩迹地表层土壤P含量相对较低且变幅小(表5)。且2个关键种不同器官的CNP分配策略相似,均优先保障果实和叶片的NPK含量以繁衍后代和维持自身生长代谢,不断推动冰川退缩迹地植被演替的进行。植被演替年限稍长于柳叶菜的马河山黄芪茎P含量与叶片P含量,叶片P含量与果实N含量间存在显著正相关关系,这有助于深入理解不同植物的养分利用及其对环境变化的响应。

表5 冰川退缩迹地植被演替初期柳叶菜和马河山黄芪不同器官N,P含量及N/P与其他研究结果比较
3.2 冰川退缩迹地植被演替初期关键种不同器官的化学计量特征
生态化学计量学是基于元素动态平衡原理和生长速率理论,可反映植物营养状况、养分需求、养分利用效率、内稳态特征以及养分限制等[1]。不同植物化学计量特征及养分分配策略存在差异,能够在一定程度上反映植物的适应能力[27]。C/N和C/P代表植物吸收养分且同化C的能力,即植物对养分的吸收利用率及固碳效率,N/P代表对生产力起限制作用的指示剂,能影响植物群落的结构和功能[28]。生态化学计量比能有有效反应植物养分循环与限制作用。可利用C/N,N/P,C/P生态化学计量分析揭示生态系统中养分的可获得性以及C,N,P元素的循环、平衡机制与相互制约关系。N,P作为陆地生态系统中植物生长的主要限制元素,N/P可反映生态系统中植物受N,P养分的供应与限制特征[29]。当植被叶片N/P<14时,主要是受到N的限制,当N/P>16时,主要是受到了P的限制,当14 冰川退缩迹地演替初期柳叶菜和马河山黄芪两种关键种不同器官C,N,P含量及其化学计量比存在相关关系。C,N表现为极显著正相关,说明地上器官叶片与地下器官细根养分分配具有一致性,功能联系密切。柳叶菜根系、茎、叶片C/N间表现为相互促进、协同增长的正相关关系,柳叶菜果实N/P与叶片N/P间存在显著正相关关系(p<0.01)。马河山黄芪茎P含量与叶片P含量间存在极显著正相关关系(p<0.01),叶片P含量与果实N含量间存在显著正相关关系(p<0.05),根系C/N与叶片C/P间,茎N/P与茎C/N间,茎N/P与叶N/P间,叶片C/N与叶片N/P间存在极显著正相关关系(p<0.01),马河山黄芪茎N/P与茎C/P间存在显著负相关关系(p<0.05),茎C/P与茎C/N,与果实C/N间存在显著负相关关系(p<0.05)。说明植物不同器官在进行C,N,P分配过程中,能量如果更多地用于生长,则会减少同化产物贮存。 (1) 柳叶菜根系有机碳含量较马河山黄芪根系有机碳含量高。马河山黄芪为豆科固氮植物,其根系全氮含量显著高于柳叶菜根系全氮含量。柳叶菜不同器官的全磷含量较马河山黄芪不同器官的全磷含量高。 (2) 柳叶菜不同器官的C,N,P含量均低于全球平均水平和我国平均水平,马河山黄芪不同器官的N含量略高于全球和我国平均水平,C,P含量均低于全球平均水平,2种关键种的各养分分配策略相似,均优先保障果实和叶片的N,P,K含量以繁衍后代和维持自身生长代谢推动冰川退缩迹地植被演替的进行。 (3) 冰川退缩迹地植被演替初期的柳叶菜不同器官的C/N高于马河山黄芪,高C/N值反映出柳叶菜具有较高的碳同化效率、较慢的生长速率及较低的营养需求,即资源保护策略。马河山黄芪高N/P和低C/P值反映了其具有高生长率或繁殖输出,即生长竞争策略,与柳叶菜相比,马河山黄芪具有快速生长的特性,以生长竞争优势即生长竞争策略战胜柳叶菜,从而促进冰川退缩迹地植被原生演替的向前推进。3.3 冰川退缩迹地植被演替初期关键种不同器官化学计量比的相关关系
4 结 论
