大花黄牡丹的开花特性与繁育系统
2021-06-22陈庭巧
唐 英,袁 涛,陈庭巧
(北京林业大学 园林学院,国家花卉工程技术研究中心,城乡生态环境北京实验室,林木花卉遗传育种教育部重点实验室,花卉种质创新与分子育种北京市重点实验室,北京 100083)
大花黄牡丹(Paeonialudlowii),芍药科芍药属牡丹组落叶灌木,主要分布于西藏林芝市巴宜区至米林县海拔2 900~3 500 m的雅鲁藏布江河谷及山坡林缘的狭长地带,为中国西藏特有[1-2],自然分布区内仅依赖种子繁殖后代,被列为极危(CR A2c)等级[3],其株形高大、花色金黄且一枝多花,是牡丹新品种培育主要的目标性状。
植物的开花特性可以从花部构成和花的开放式样两个层次来认识[4]。花部构成是指包括花的结构、颜色、气味和蜜汁产量等单个花的所有特征;花的开放式样是指花在某一时间开放的数量和在花序上的空间排列,可看作花在群体水平上表现出的特征[5]。繁育系统通常的定义是代表直接影响后代遗传组成的所有有性特征[6],是植物内部的遗传机制和外部环境相互作用的一种表现形式,它们结合传粉者和传粉行为是影响生殖后代遗传组成和适合度的主要因素,在决定植物的进化路线和表征变异上起着重要作用[7-8]。物种濒危其实就是该种生活史中的一个或几个环节受内外因素的影响出现危机而使生物个体数量下降的现象[7],濒危物种繁育系统的研究可探究其生殖过程中可能存在的致濒原因,为濒危植物的保护和利用提供参考。
芍药属牡丹组所有野生种均已列入《中国物种红色名录(第一卷)》,濒危等级各异。大花黄牡丹是牡丹组中濒危等级最高的种之一[3]。但仅四川牡丹(P.decomposita)、滇牡丹(P.delavayi)、‘凤丹’(P.ostii‘Fengdan’)和矮牡丹(P.suffruticosasubsp.spontanea),可见有关繁育系统类型或开花授粉特性的报道,四川牡丹和‘凤丹’为混合交配系统,滇牡丹和矮牡丹为专性异交[9-12]。本研究观测大花黄牡丹开花过程并进行人工授粉控制实验,分析大花黄牡丹的开花特性和繁育系统类型,为解析可能影响其开花结实的因子,探讨其原产地条件下濒危的原因和资源开发利用提供参考。
1 材料和方法
1.1 时间和地点
2019-2020年5~9月于河南省洛阳市栾川县三川镇(33.94°N,111.40°E;海拔1 300 m)进行。2019年10月至西藏自治区林芝市巴宜区米瑞乡曲尼贡嘎村(29°29′59″N,94°37′01″E;海拔3 044 m)采收野生大花黄牡丹果实和种子。
1.2 材 料
试验地内大花黄牡丹开花株共计666株,为采自西藏自治区林芝市米林县南伊沟(29°19′10″N,94°20′25″E)的野生大花黄牡丹种子育成,2002年秋季播种,次年少量萌发成苗,2008年首次开花。2015年起疏于管理,每年仅夏季清除一次地面杂草。至2020年试验地内大花黄牡丹植株最大株龄17年,平均株龄约10年,稳定开花。
1.3 方 法
1.3.1 花部特征与开花物候盛花期选取不同植株10朵花,测量每朵花的直径,花瓣、苞片和萼片的长、宽及数量,雌雄蕊数量以及心皮长、宽、厚。随机标记不同植株的顶蕾和侧蕾各10个,观测记录露瓣、展瓣和落瓣的日期和天数。自试验地第1朵花开放日起,每天记录当天正值开放状态的花朵数。
1.3.2 花粉-胚珠比(Pollen-ovule ratio,P/O) 在不同植株上选取即将开放的20个花蕾,每个花蕾取3枚花药于开盖离心管中自然散粉后,滴入蒸馏水制成4 mL花粉溶液,取1 μL花粉溶液于载玻片,显微镜下统计花粉粒数量,重复10次。最后计算每朵花的花粉数。解剖刀划开心皮,统计胚珠数。单花花粉量=视野平均花粉数×稀释倍数(4000)/3×花药总数。
1.3.3 杂交指数(Outcrossing index,OCI) 取盛花期10朵花,测花朵直径后观测柱头与花药的空间位置,以最高的花药与柱头的位置为判断依据,根据Dafni的标准[13],计算杂交指数并判断繁育系统类型。(1)花朵或花序直径:<1 mm记为0,1~2 mm记为1,2~6 mm记为2,>6 mm记为3。(2)观察花药开裂与柱头具可授性的时间先后:同时或雌蕊先熟记为0,雄蕊先熟记为1。(3)柱头与花药的空间位置:同一高度记为0,空间分离记为1。(1)-(3)累计得OCI 值。
1.3.4 花粉粒活力在不同植株上选取同日开放的5朵花并标记,从标记当天至花瓣落尽期间,每日9点分别采集花粉粒后立即检测。将花粉粒均匀散布于载玻片,滴1滴0.5% TTC溶液,盖上盖玻片置于40 ℃恒温处理10 min后,显微镜(N-117M型)观察,每个载玻片取3个40倍的视野计数,红色花粉粒为有活力,未染色的花粉粒为无活力。花粉活力=红色花粉数/总花粉数×100%。
1.3.5 柱头可授性每天随机选取3朵当日开放的花朵标记并套袋,直到第1天标记套袋的花朵柱头完全变黑,最后1天剪取所有标记花朵的柱头和3个露瓣透色花朵的柱头。柱头取回后,装入离心管并按日期顺序分3组。每个离心管依次倒入1 mL现场配制的联苯胺-过氧化氢溶液(联苯胺∶3%过氧化氢∶蒸馏水=4∶11∶22),5 min后记录显色和气泡变化。
1.3.6 人工控制授粉实验2019年,随机选取不同株的开花枝,设置8组处理,每组10朵:(1)自然授粉(对照): 随机选取并标记10个透色顶花蕾,不做任何处理;(2)自然授粉:随机选取并标记10个透色侧花蕾,不做任何处理;(3)直接套袋:随机选取10个透色顶花蕾,套硫酸纸袋并标记,(4)去雄套袋:随机选取10个透色顶花蕾,去雄后套上硫酸纸袋并标记;(5)直接套网袋:随机选取10个透色顶花蕾,不去除雄蕊,直接套防虫网袋并标记;(6)同株异花授粉:随机选取10个透色顶花蕾,去雄后套上硫酸纸袋并标记;3 d后授以所在植株上其他花朵的花粉;(7)异株异花授粉:随机选取10个透色顶花蕾,去雄后套上硫酸纸袋并标记;3 d后授以不同植株的花粉;(8)花头朝向处理:随机选取10个透色顶花蕾,用小木棍支撑花茎使自然下垂的花朵直立向上,并标记。
2020年较2019年增加如下处理组,每组随机选取15个透色顶花蕾:(1)仅去除雄蕊并标记;(2)去除雄蕊,套防虫网袋并标记;(3)仅去除花冠并标记;(4)去雄蕊+去花冠并标记。
2019年9月、2020年9月蓇葖果变色时采收,计算结果率和结实率。结果率=(结实花朵数/处理花朵数)×100%,结实率=(饱满种子/胚珠数)×100%。
1.3.7 访花昆虫随机选取3朵花,从开花第1天至花瓣落尽期间,每天6:00-7:00、8:00-9:00、10:00-11:00、12:00-13:00、14:00-15:00和16:00-17:00拍摄、记录访花昆虫的种类、访花行为和访花时长,捕捉访花昆虫制作标本用于鉴定。
2 结果与分析
2.1 开花特性和泌蜜现象
2.1.1 开花特性大花黄牡丹每个开花枝着花1~4朵,生于枝顶(顶花)和叶腋(侧花)。单花直径78.65~106.84 mm,花瓣10~13枚,苞片2~6枚,萼片4~6枚,心皮1~2个(表1)。通常顶花最先开放,随后同枝的侧花从下往上依次开放(图1,A)。花朵昼夜开放程度不一,可见昼开夜合现象(图2)。花头通常在花冠闭合时倾斜朝上,随着花冠展开而缓慢下垂,然后在花冠逐渐闭合的同时又慢慢回升(图3);下雨时光线变暗花冠也会适度闭合且下垂。群体花期后期开花畸形多,少数无雌蕊或3心皮,雄蕊、花瓣易脱落,有的花冠成杯状不能完全展开。
2.1.2 泌蜜现象花朵开放前花蕾表面无可见花蜜,开放后有淡香。花丝、花药、雌蕊子房与花瓣之间部位可见花蜜(图1,B、C),后者花蜜量较大,花丝和花药上的花蜜则呈细小水珠状零星分布。前1天傍晚去除雄蕊套上硫酸纸袋,第2天早晨在子房与花瓣之间仍可见花蜜,用移液枪吸取花蜜后再次套硫酸纸袋以排除访花昆虫吸食花蜜和花蜜蒸发,次日仅少量花朵在同一位置仍有花蜜,大部分花朵已无可见花蜜。观察结果显示大花黄牡丹具花内蜜腺,泌蜜部位应主要在子房与花瓣之间。并非所有花朵都可见花蜜,花梗较短、叶片颜色偏黄的花朵较难观察到花蜜,长势较弱、花期短的花朵花蜜量明显少于正常花朵。
2.2 开花物候
2.2.1 群体花期2019年试验地大花黄牡丹群体花期5月17日开始,6月19日结束,持续34 d(图4),为集中开花模式。其中5月24日~5月30日,每日正值盛开的花朵数均超过200,5月27日达到峰值250朵。
2.2.2 单花花期参考杨翔[14]的方法,根据花朵开放过程中的形态变化,将单花开放过程(单花花期)划分为露瓣期、展瓣期、落瓣期3个时期(图5,表2)。2019年,平均单花花期为(8.95±1.28) d,其中顶花单花花期(8.70±0.95) d,侧花(9.20±1.55) d。侧花的展瓣期与顶花相同,为5.4 d,侧花的露瓣期和落瓣期都较顶花长,故侧花单花花期略长于顶花。

A. 开花顺序为1→2→3(1为顶花,2和3为侧花);B、C. 花蜜位置(红圈示花蜜及位置)图1 大花黄牡丹花朵开放次序(A)以及花蜜位置(B、C)A. The order is 1→2→3 (1 is top flower. 2 and 3 are side flowers); B and C. Nectar location of P. ludlowii (Red circles indicate nectar and location)Fig.1 Flowering order on a branch and nectar location of P. ludlowii flowerlet

A、C、E和G依次为开花第1天、第2天、第3天和第4天7:00;B为开花第1天17:00;D、F为开花第2天、 第3天12:00;H为开花第4天的11:00图2 大花黄牡丹昼夜开花动态A, C, E and G are 7:00 O’clock on the first, second, third and fourth day in florescence, respectively; B is 17:00 O’clock on the first day of flowering; D and F are 12:00 O’clock on the first and second day in florescence, respectively; H is 11:00 O’clock on the fourth day in florescenceFig.2 P. ludlowii flowers bloom day and night dynamic

A-H依次为开花第2天7:00(花朵直立向上),8:00(花朵微倾斜朝上),9:00(花朵倾斜朝上),10:00(花朵朝上,倾斜程度增大), 12:00(花朵倾斜朝下),14:00(花朵朝下,倾斜程度增加),15:00(花朵朝下,倾斜程度减小)和17:00(花朵倾斜朝上)图3 大花黄牡丹同一天内花朵朝向变化A-H are 7:00 O’clock (Flower stands upright), 8:00 O’clock (flower tilts slightly upward), 9:00 O’clock (flower tilts upward), 10:00 O’clock (flower tilts upward, increasing in inclination), 12:00 O’clock (flower tilts down), 14:00 O’clock (flower tilts down, increasing in inclination), 15:00 O’clock (flower tilts down, decreasing in inclination) and 17:00 O’clock (flower tilts upward) on the second day, respectivelyFig.3 Changes in P. ludlowii flowers facing-orientation within a day

表1 大花黄牡丹花部形态指标

图4 试验地大花黄牡丹的群体花期(2019年)Fig.4 The groups florescence of P. ludlowii in Luanchuan, Henan province in 2019
2.3 花粉活力与柱头可授性检测
表3显示,大花黄牡丹开花第1天时花粉粒不具活力,第3天花粉活力最强,为(93.86±3.10)%;第4天花粉活力仍较强,为(91.37±1.66)%;第7天花瓣开始脱离,花药由鲜黄色变深并逐渐干瘪,花粉粒不再具有活力。柱头可授性检测结果(表4)发现,开花前1天,花朵呈透色状态,此时已具可授性,开花第3天(展瓣期)和第4天(展瓣期)可授性最强,至第12天(花瓣落尽)起柱头可授性消失。大花黄牡丹花粉活力最好的时期与柱头可授性最强的时期高度重合,皆在开花第3天和第4天,故本研究认为此时是人工杂交授粉最佳时期。
2.4 繁育系统检测
2.4.1 花粉-胚珠比按照Cruden的标准[15]:P/O为2.7~5.4时,为闭花受精;P/O为18.1~39.0时,为专性自交;P/O为31.9~396.0时,为兼性自交;P/O比为244.7~2 588.0时,为兼性异交;P/O比为2 108.0~195 525. 0时,为专性异交。
由表5可知,大花黄牡丹单花平均292个花药(最多399个、最少211个),6 770 333粒花粉(最多10 237 333粒,最少3 461 333粒),20个胚珠(最多29个,最少9个)。P/O为119 356.31~731 238.07,平均P/O有375 502.28,20组单花P/O中仅编号15和17在2 108.0~195 525. 0之间,其余组和平均值均超出Cruden的标准范围。

A.未露瓣花蕾;B、C.露瓣期;D-I.展瓣期;J-L.落瓣期图5 大花黄牡丹单花开放进程A. Flower buds whose petals are not exposed; B, C. Petal-appearing period; D-I. Opening petal-opening period; J-L. Falling petal-falling periodFig.5 The single flower blooming process of P. ludlowii
2.4.2 杂交指数根据Dafni[13]的标准:OCI = 0为闭花受精;OCI = 1为专性自交;OCI = 2为兼性自交;OCI = 3为自交亲和,有时需要传粉者;OCI = 4为异交、部分自交亲和、需要传粉者。表6显示,大花黄牡丹OCI值为4,初步判断其繁育系统类型为异交、部分自交亲和、需要传粉者。
2.4.3 人工控制授粉2020年,试验地内大花黄牡丹开花数量较2019年多,但部分花蕾不展开,雄蕊脱落,结果率与结实率显著低于2019年(表7),同一授粉处理的结实情况也差异较大(表8)。
2019年试验地自然授粉的结实率为(32.50±8.29)%,略高于同年西藏林芝巴宜区米瑞乡(29.01±15.57)%,但2020年试验地顶花结果率和结实率分别为46.67%、(14.33±6.26)%,较 2019年的100%、(33.75±9.10)%下降明显。2020年侧花结果率100%,侧花结实率(19.02±9.54)%,侧花结实率虽比2019年(31.25±7.67)%低,但仍高于2020年顶花的结实率(14.33±6.26)%,且连续两年所有侧花均全部结果;说明侧花的开花结实比顶花稳定,杂交时可优先给侧花授粉。
去雄套袋组2019年既结实又有正常种子,结实率(31.48±5.56)%,而2020年结实为0,说明大花黄牡丹可能存在不稳定的无融合生殖。但牡丹组中仅见四川牡丹具微弱无融合生殖的报道[9],其无融合生殖需要进一步确认。

表2 大花黄牡丹单花花期(2019年)

表3 大花黄牡丹花粉粒活力日变化
直接套袋组和同株异花授粉组2019年与2020年皆有结实,自交结实率分别为(32.18±9.02)%、(15.52±6.24)%,可知大花黄牡丹具自交亲和性。网袋组2019年结实率为(30.21±5.79)%,略低于对照组的(33.75±9.10)%;2020年网袋组结实率(18.21±8.46)%,高于对照组的(14.33±6.26)%,但无显著性差异。2020年同株异花授粉的结实率为(16.95±5.93)%,异株异花授粉的结实率为(14.64±4.62)%,与对照组无显著性差异。
去花冠组的结实率为(18.46±8.69)%,略高于对照组。仅去雄组、去雄套网袋组和去雄蕊+去花冠组均未结实,去除雄蕊会严重影响传粉过程。去除雄蕊意味着花粉丧失,花朵既不能自花授粉,又失去传粉报酬,大大降低了其传粉成功的概率。花冠朝上组2019年与2020年的结实率分别为(33.75±8.66)%和(14.38±7.20)%,与同年对照组的结实率近乎一致。

表4 大花黄牡丹雌蕊柱头可授性日变化

表5 大花黄牡丹的花粉-胚珠比

表6 大花黄牡丹的杂交指数
表8 大花黄牡丹不同授粉处理的结果

表7 原生地与试验地自然授粉的结实率
2.5 访花昆虫
试验地内大花黄牡丹花期访花昆虫共16种(表9),其中膜翅目(Hymenoptera)3种,双翅目(Diptera)4种,半翅目(Hemiptera)4种,鞘翅目(Coleoptera)4种,蜘蛛目(Araneae)1种。主要访花昆虫有膜翅目的意大利蜜蜂(Apismelliferali-gustica)、芦蜂(Ceratinasp.)和隧蜂(Halictussp.)以及双翅目的灰带管蚜蝇(Eristaliscerealis)和黑带食蚜蝇(Episyphusbalteatus)。
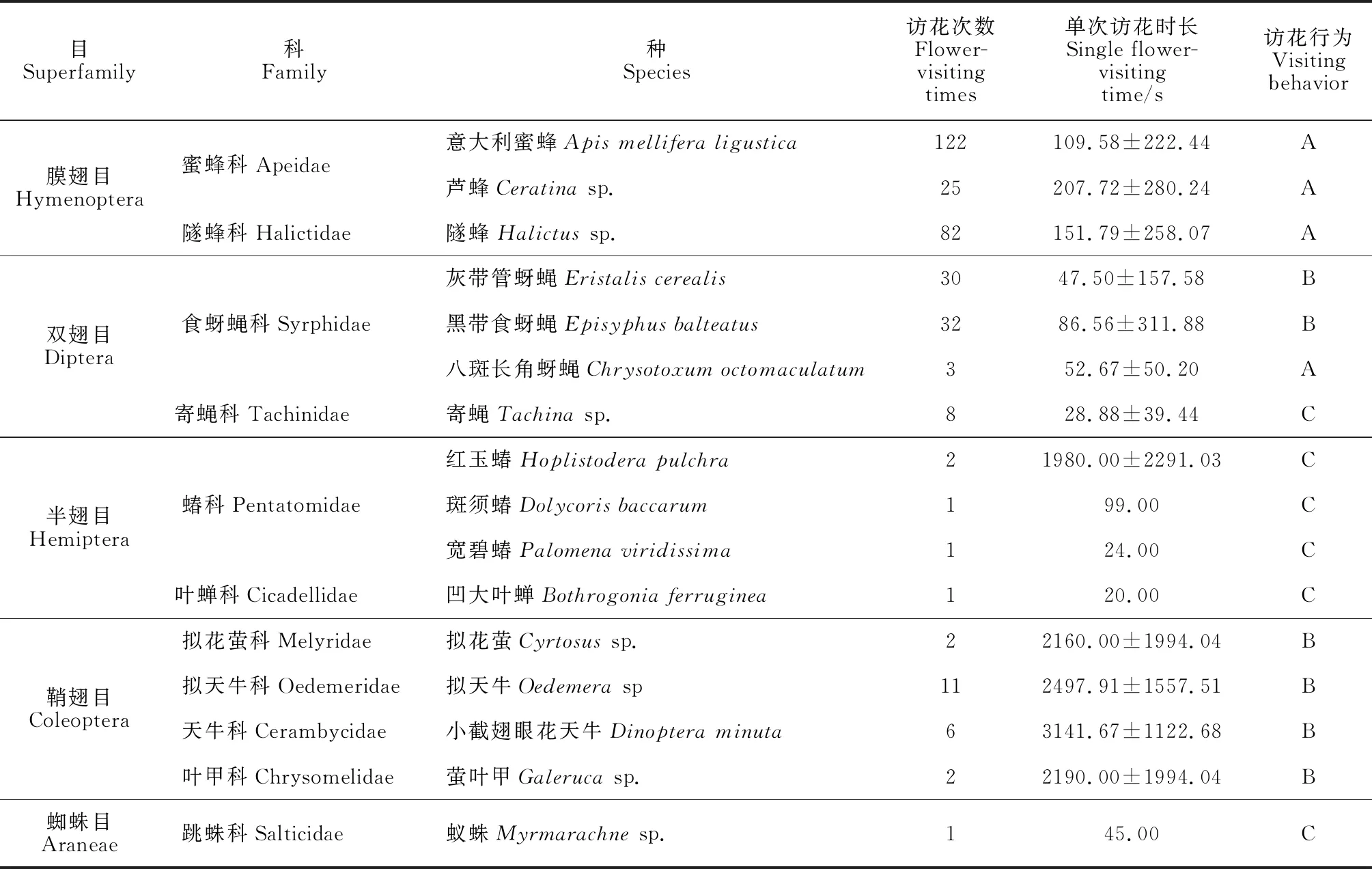
表9 大花黄牡丹访花昆虫种类及访花行为
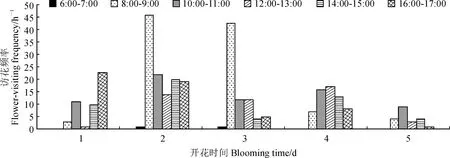
图6 大花黄牡丹不同时段访花昆虫的访花频率Fig.6 The visiting frequency of pollinators in different periods of P. ludlowii flowering
2.5.1 不同种类访花昆虫的访花行为、访花次数及时长膜翅目访花昆虫的主要访花行为是采集花粉,双翅目既吸食花蜜又采花粉。鞘翅目主要吸食花蜜,半翅目多在花上休憩,花蜜和花粉不是鞘翅目昆虫的访花目的,寄蝇(Tachinasp.)和蚁珠(Myrmarachnesp.)也只是在花上停留。膜翅目访花次数、访花时长在所有访花昆虫中居首位,其中意大利蜜蜂访花最勤,观察期内共访花122次,单次访花时长(109.58±222.44)s;其次是隧蜂,访花82次,平均每次(151.79±258.07)s;芦蜂访花22次,单次停留(207.72±280.24)s。灰带管蚜蝇和黑带食蚜蝇访花次数相近,分别有30次和32次,但黑带食蚜蝇单次访问时间近1.5 min,灰带管蚜蝇单次停留时间不足1 min。4种鞘翅目访花昆虫虽然访花次数不多,但滞留时间较长,单次访花时长均超过2 000 s。
2.5.2 不同时段访花昆虫的访花频率由图6可知,访花昆虫在开花后第2天的访花频率最高,每小时可访问20次;开花后第3天访问频率次之,每小时访问13次。一天中访花次数最多的时段是8:00-9:00,共计103次。开花第2天的8:00-9:00访花次数高达46,为各时段最高,其次是开花第3天的8:00-9:00,访花次数为43。10:00-11:00这一时段的访花频率也不低,每小时14次,合计访花70次。
3 讨 论
3.1 大花黄牡丹的繁殖保障
植物与其传粉者的相互作用被认为是被子植物花进化的重要推动力,相互作用表现在花部特征对访问者行为和花粉传递机制的影响,被访问者传送的花粉数量和质量又反作用于植物作为雌性或雄性亲本的生殖成功[16]。花自身的结构特点、展示、设计方式及引诱传粉者的能力是影响植物繁殖成功的重要因子[17]。植物个体开花物候(开花持续时间和开花强度格局)常在两个极端之间变化,即集中开花模式和持续开花模式,一般认为,集中开花模式有利于物种吸引更多昆虫,有助于传粉成功[18]。大花黄牡丹花大,花色明黄,花香清淡、集中开花可增强其对传粉者的吸引力。花粉和花蜜富含营养,可作为传粉者访花报酬补偿其能量消耗[19]。大花黄牡丹分泌花蜜,单花花粉量(3 461 333~10 237 333)大,花朵下垂可避免雨水冲刷造成的花粉损失,有效增加访花昆虫的访问频率。
暂时性花闭合是花对环境条件如雨水、露水、低温、风或有限的传粉者的响应,是为保护植物繁殖器官及提高繁殖成功而进化出的花部机制[20-21]。昼开夜合的开放方式可节约能量,延长花寿命[7],大花黄牡丹这一开放方式有效地保护了其花朵寿命。其展瓣期为(5.40±0.75)d,且期间花粉活力为(76.97±4.50)%~(93.86±3.10)%,柱头可授期长达12 d,延长了授粉时间。花粉活力和柱头可授性因植物而异,花粉与柱头同时处于高度活力状态,有利于植物顺利完成授粉、识别、受精过程[18]。大花黄牡丹大部分花药高于柱头,花粉活力最好与柱头可授性最强时期高度重合(开花后第3、4天),在此期间,以采集花粉为主要行为特征的3种膜翅目昆虫访花次数、访花时长在所有访花昆虫中居首位,花蜜也吸引了双翅目和鞘翅目访花昆虫,昆虫种类多,行为活跃,这些都增加了授粉成功的机率。大花黄牡丹的花部综合特征、性器官寿命和花开放式样、访花昆虫为其传粉过程提供了较充分的保障。
尽管如此,对比原产地和本试验2年的数据,大花黄牡丹自然授粉结实率只在(17.52±8.77)%~(32.50±8.29)%之间。改变传粉条件如去除花冠、适度阻断空气流动(直接套硫酸纸袋)、阻断传粉者访花(套网袋)以及改变花朵朝向,并未显著降低结实率,人工保障授粉(同株异花和异株异花授粉)也没有显著提高结实率。2年的试验结果也显示其可自交结实,侧花比顶花结实更稳定,本研究试验中还观察到大花黄牡丹无融合生殖现象。这些表明,大花黄牡丹的交配机制可能比我们认为的更灵活,既可异交授粉结实又可自交结实,开花特性又有多重保障机制。故本研究认为,大花黄牡丹结实率的主要影响因子可能不在于传粉过程和交配机制,可能与传粉后受精、果实或种子发育过程等因素有关,还需今后扩大实验数量并结合更多方法深入研究。
另外,根据李嘉珏等的报道,甘肃榆中和平牡丹园引种栽培4年的大花黄牡丹植株,每年人工浇水、喷施磷酸二氢钾3次,其套袋观察结果显示自花授粉结实率为66.7%[22],本试验自花结实率远低于这一数据的原因也可能与试验地点疏于管理有关。
3.2 大花黄牡丹的繁育系统类型及意义
本研究获得的大花黄牡丹的20组P/O数据,仅2组数据P/O值在Cruden的标准内[15],其他各组数据远超出评判标准P/O值。骆劲涛的研究发现,四川牡丹的P/O(120 000~270 000),也仅部分数据在Cruden标准范围内[9],鉴于此,结合OCI值和人工控制授粉实验结果,综合判断,本研究对大花黄牡丹繁育系统类型的判断与四川牡丹一致[9]:即异交、部分自交亲和、需要传粉者的混合交配系统。罗长维等、李奎等发现牡丹组‘凤丹’P/O(58 940)和滇牡丹P/O(6 124~9 713)虽都在专性异交的标准范围,但‘凤丹’OCI和授粉实验结果显示其繁育系统为以异交为主,自交部分亲和,传粉过程需要传粉者[10-11],虽然‘凤丹’P/O值与大花黄牡丹和四川牡丹差异较大,但三者的繁育系统类型一致。滇牡丹的繁育系统则为专性异交型,且和矮牡丹一样被认为是异花授粉植物,即使它们存在微弱的自交性[10, 12]。有意义的是,大花黄牡丹、四川牡丹和‘凤丹’都是专性有性繁殖的种类,自然界仅依赖种子繁殖,混合交配系统更有利多产种子;而滇牡丹和矮牡丹为兼性营养繁殖的种类,以多种形式的营养繁殖为主,种子繁殖为辅。即使专性异交,其繁殖特性也可保证后代数量。芍药属繁育系统研究范围可进一步扩大到其他种类,以全面分析该属繁育系统类型多样性的原因及与自然繁殖特性的内在联系。
访花昆虫中双翅目和鞘翅目都吸食花蜜,双翅目同时也采集花粉,花蜜是否影响其P/O值尚待深入探讨。除大花黄牡丹外,滇牡丹[10]花朵、芍药(Paeonialactiflora)品种[23]的花蕾也有分泌花蜜的报道。芍药属植物是否具有典型的花蜜腺及其类型、具体位置和泌蜜规律有待进一步研究。
3.3 依据P/O判断繁育系统类型的适用性
目前判断植物繁育系统类型,都会将P/O值作为依据之一,但P/O值在繁殖生物学、授粉生态学、系统分类学等方面的应用未必完全适用于所有分类群[24]。Cruden采用的大多是亲缘关系较远的种类归纳P/O的范围[15, 25],涉及物种范围有限。且其归纳的标准除闭花受精外,每两个相邻类型的P/O范围都有重叠,对于P/O值介于重叠部分的物种此标准不具精确性。花和花序的构造特性、授粉有效性以及生境等会影响P/O值的大小,而亲缘关系近的种间不同的演化史,致使其P/O值差异非常大的情况下,也可能具有相似的繁育系统[26]。如天目地黄(Rehmanniachingii)和湖北地黄(R.henryi)的P/O值显示其繁育系统类型为兼性自交,裂叶地黄(R.piasezkii)和地黄(R.glutinosa)为兼性异交,事实上这4种地黄均属于异交,部分自交亲和,需要传粉者的类型[27]。有些植物,如榕属植物因花粉粒细小,曾喜育采用花药-胚珠比(A/O)代替P/O研究其繁育系统[28],作为区分繁育系统差异的参考。本研究试图计算大花黄牡丹的A/O值,但未得到可供分析比较的结果。
大花黄牡丹P/O超出标准范围并非个例。东北地区野豌豆属均为自花授粉植物,其P/O 超出 Cruden 标准中自花授粉的阀值,多数种类P/O值低于1 000,少数种类高于1 000[29]。凤仙花科(Balsaminaceae)中各个繁育系统类型都存在极高的P/O值,仅部分符合分类标准[30],华北驼绒藜(Ceratoidesarborescens)P/O(1 351 189 ± 391 779)远超Cruden的标准[31]。另外,尽管有些物种的P/O值没有超出标准范围,根据P/O判断的繁育系统类型却与实际情况不符,如百子莲(Agapanthuspraecoxsubsp.orientalis‘Big Blue’)[32]、喜树(Camptothecaacuminata)[33]、华北蓝盆花(Scabiosatschiliensis)[34]和吉首蒲儿根(Sinoseneciojishouensis)[35]等。Chouteau在研究天南星科(Araceae)Philodendron属23个种和Anthurium属20个种的P/O后甚至指出,P/O可能与其研究对象的繁育系统没有直接联系[36]。因此,植物繁育系统的判断,花粉/胚珠(P/O)可视为参考指标,而不是唯一的判断指标。
