毛果杨WRKY基因家族鉴定与表达分析
2021-06-21孙晶琦许炜清孙晨许晓倩陈晶晶王晓立
孙晶琦 许炜清 孙晨 许晓倩 陈晶晶 王晓立

摘 要:WRKY转录因子是植物中最大的转录调控家族之一,是调控植物许多生物过程信号网络的组成部分,具有多种生物学功能。该研究根据已公布的拟南芥基因组中当前确定的WRKY基因家族成员,利用其序列进行结构域的分析,从而确定WRKY基因家族的基本特征,并以此为基础,从毛果杨序列库中筛选出WRKY基因家族的成员。再利用MEGA5软件、在线工具ExpasyProtparma、BioEdit等进行理化性质分析、motif结构对比、进化树构建、基因表达分析等。结果表明:毛果杨WRKY基因家族20个成员编码序列长为189~624bp,有多个染色体定位重复分布于第1、5、14号染色体上。WRKY蛋白均定位于细胞外,亲水性较差,具有较高的脂溶性和不稳定性。20个WRKY基因均在毛果杨中表达,但有明显的差异。
关键词:毛果杨;WRKY蛋白;基因家族;生物信息
中图分类号 Q943.2文献标识码 A文章编号 1007-7731(2021)09-0016-04
Abstract: WRKY transcription factors, one of the largest transcriptional regulatory families in plants, are part of the signal networks that regulate many biological processes in plants, and have a variety of biological functions. In this study, the WRKY gene family members were identified in Arabidopsis thaliana on the published Arabidopsis genome, and the basic characteristics of the WRKY gene family were determined by using their sequences for domain analysis. On this basis, the members of the WRKY gene family were screened out from the sequence library of Populustrichocarpa. MEGA5 software, online tools such as ExPASyProtparma and Bioedit were used to analyze physical and chemical properties, motif structure comparison, evolutionary tree construction and gene expression analysis. The results showed that the coding sequence length of 20 members of the WRKY gene family was 189~624bp, and multiple chromosomes were located and repeated on chromosomes 1, 5 and 14. All WRKY proteins were extracellular, with poor hydrophilicity and high lipid solubility and instability. All the 20 WRKY genes were expressed in Populustrichocarpa, but there were significant differences.
Key words: Populustrichocarpa; WRKY proteins; Gene family; Biological information
轉录调控是生物在生长发育及代谢过程中调控细胞或组织中基因表达的重要机制[1]。转录因子(Transcription factors,TFs)是一类锚定在目标基因启动子区域特异序列上的蛋白质,能对下游目标基因的转录进行促进或抑制作用,对生物体的正常生长发育和应对外界胁迫过程都具有重要的作用[1,2]。WRKY基因家族是近年来发现的一类植物特有的转录调节因子,它是一种植物特有的调控因子,已从多种植物中分离,如甜土豆[3]、野燕麦[4]、皱叶欧芹[5]、拟南芥[6]、烟草[7]等。根据目前所有的研究资料来看,WRKY蛋白在植物的各项生理活动过程中发挥着重要的调控作用,如胚胎形态的发生[8]、叶片的衰老等[9],同时在非生物胁迫包括寒冷、干旱、盐碱等[10]和生物胁迫包括细菌、真菌、病毒等[11]逆境响应及信号转导过程中也起着重要的调控作用。
目前,对毛果杨(Populustrichocarpa)WRKY基因家族研究尚不深入。本研究通过各种途对选中的20个毛果杨WRKY基因进行理化性质分析、motif结构比对、进化树构建、基因表达分析等,为今后更深入的研究奠定基础。
1 数据来源与分析方法
1.1 数据来源 以毛果杨、拟南芥为实验材料。毛果杨(Populustrichocarpa)、拟南芥(Arabidopsis thaliana)WRKY家族成员的基因序列信息均在美国生物技术信息中心即NCBI进行的下载。
1.2 分析方法
1.2.1 毛果杨中WRKY基因的确认与分类 根据已公布的拟南芥基因组中当前确定的WRKY基因家族成员,利用其序列进行结构域的分析,从而确定WRKY基因家族的基本特征,以此为基础,从毛果杨序列库中筛选出WRKY基因家族的成员。
1.2.2 序列比对和进化树分析 利用MEGA5软件,对毛果杨的20个WRKY基因进行氨基酸序列比对,分析它们的同源性。毛果杨的20个WRKY基因和拟南芥(Arabidopsis thaliana)WRKY家族的5个成员进行系统进化树构建,将bootstrap值设置为1200次,根据不同的需求选择不同的进化树格式。
1.2.3 WRKY蛋白的理化性质分析 毛果杨WRKY蛋白相关基因的DNA长度、基因序列长度、CDS序列以及外显子数量均从Phytozome的毛果杨数据库中获得。毛果杨WRKY蛋白的氨基酸数量、分子量、脂溶性、亲水性和稳定性均通过在线工具ExpasyProtparma进行研究分析。对WRKY蛋白的亚细胞定位使用Plant-mPLoc进行在线预测。
1.2.4 基因组织表达分析 在Affy网站(http://www.affymetrix.com /estore/index.jsp)检索毛果杨WRKY基因家族成员的探针集,然后通过BAR数据库(http://bar.utoronto.ca /welcome.htm)分析并在线绘制热图,研究其在不同组织中的表达模式以及相对表达量。
2 结果与分析
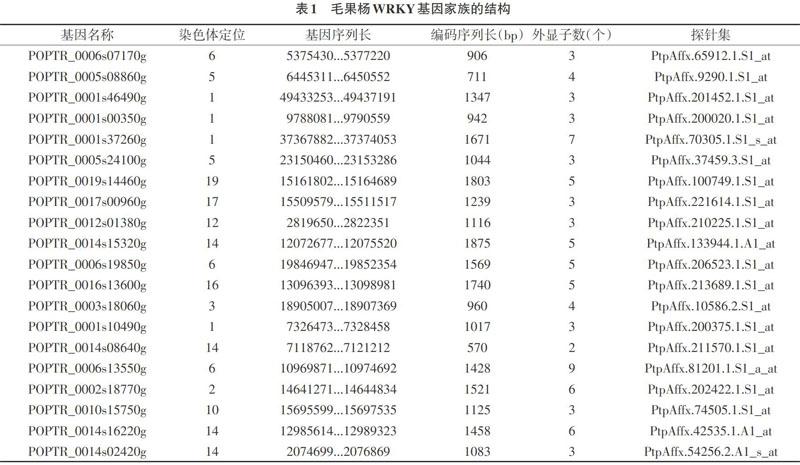
2.1 毛果杨WRKY基因家族的结构 通过分析,到最后选用20个毛果杨WRKY基因家族成员(见表1)。WRKY基因的序列长度差异较大。其中POPTR_0014s08640g只有570bp,而POPTR_0014s15320g有1875bp。除POPTR_0014s08640g和POPTR_0006s13550g基因外,外显均为3-7个。从上面的数据分析可以看出,不同的基因有着不同的结构形式,造成了它们可以用不同的方式发挥不同的作用。
2.2 毛果楊WRKY基因序列的同源性 利用MEGA5软件,对20个毛果杨WRKY蛋白家族的氨基酸序列进行在线分析(见图1)。标注标星号序列的同源性较高(515D-592H),表现出高的保守性。
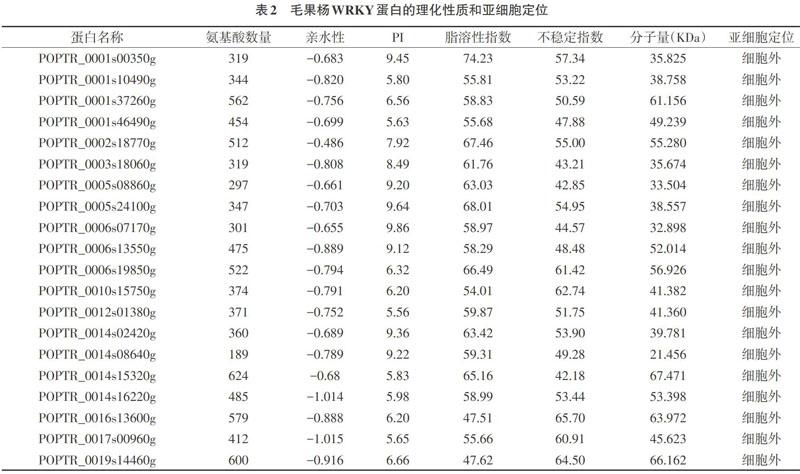
2.3 毛果杨WRKY基因的理化性质 对毛果杨WRKY蛋白的理化性质以及亚细胞定位进行了预测分析(见表2)。由表2可知,所有WRKY家族成员的氨基酸序列均较长,含189~624个氨基酸。WRKY蛋白的等电点(PI)为5.56~9.86,说明WRKY蛋白带正电荷。WRKY蛋白的亲水性为-0.1015~-0.486,表现为亲水性低。所有WRKY家族成员的稳定性指数均大于40,表现为不稳定。WRKY蛋白的脂溶指数(Aliphatic index)较高,为47.51~74.23。预测大部分WRKY蛋白定位于细胞外,这与其行使功能的位置保持一致。
2.4 进化树的构建及分析 对拟南芥5个基因和毛果杨20个基因构建了系统进化树。如图2所示,这部分基因组成的进化树可分为以下4类:A类支持率较高,由拟南芥4个基因和毛果杨17个基因组成;剩下3类支持率较低,即B类由毛果杨1个基因组成、C类由拟南芥1个基因组成、D类由毛果杨2个基因组成。由此可见,WRKY基因家族的亲缘关系远近表现出相当高的保守性,且在进化中扮演着相当重要的角色。A类中POPTR_0002s18770g和POPTR_0014s15320具有较高的重复性,且亲缘关系紧密地联系在一起。
2.5 毛果杨WRKY基因的表达模式 为了进一步地深入剖析WRKY基因家族在毛果杨植物体中转录水平的高低,在其根、幼叶、成叶、雌蕊、雄蕊、木质部6个不同组织以及光生苗中进行了分析并绘制成图(图3)。从图3可以看出,20个WRKY基因在成叶中的表达量最低,在其他5个组织及光生苗中均有较高的表达水平。其中,POPTR_0001s00350g在营养器官中的表达量均呈现较低水平。几乎所有的家族成员都存在着差异表达,而且大多数基因在根部和雌、雄蕊处有较高的表达量。值得关注的是,POPTR_0005s08860g、POPTR_0001s37260g在木质部中的表达较高,而在其他组织中表达量低,这意味POPTR_0005s08860g、POPTR_0001s37260g可能以某种方式参与毛果杨木质部形成的分子调控过程中。而POPTR_0012s01380g主要在雌、雄蕊中表达,其可能参与雌、雄蕊的发育过程。
3 讨论与结论
3.1 讨论 基因是控制生物性状的基本遗传单位[12]。基因是个体生命活动以及发挥功能作用的基础,在它里面储存着大量的关于个体生长发育的一系列过程的信息数据。WRKY基因存在于毛果杨染色体的不同位点上,具有不同的基因形式。WRKY转录因子具有非常显著的结构特点,其蛋白结构基本含有1~2个WRKY结构域,为DNA结合域(DBD),约由60个高度保守的氨基酸残基组成,包括位于N端的七肽和位于C端的锌指结构。位于N末端的七肽WRKYGQK绝对保守,是核心序列,位于C端的序列由C2H2(C-X4-5-C-X22-23-H-X-H)或C2HC(C-X7-C-X23-H-X-C)型锌指结构组成[13-15]。WRKY蛋白普遍存在于高等植物中,在拟南芥的74个成员中,绝大多数的生物学功能还未弄清楚[16]。WRKY转录因子可通过WRKY结构域与靶基因启动子区的顺式作用元件TTGAC(C/T)核苷酸序列(W-box)特异性结合,以此激活或抑制转录,进而调控下游基因的表达。有报道拟南芥WRKY4结构域由1个四链β片层组成,在β片层的C端由保守的半胱氨酸/组氨酸(Cys/His)残基形成1个锌结合袋,WRKYGQK残基对应于序列的N端链,在序列的中间被Gly残基扭折,使得涉及Trp(色氨酸)残基的广泛疏水作用,进而促使β链的结构具有稳定性。β链的WRKYGQK基序可以接触1个大约6bp区域,这在很大程度上与W-box(TTGACY)的长度是一致的。表明WRKYGQK的基序可以与靶基因启动子区的W-box结构特异性结合。除WRKY结构域之外,WRKY转录因子还包含其他结构域,包括TIR-NBS-LRR(Toll/interleukin-1receptor-nucleotidebindingsite-leucine-richrepeat)、激酶结构域、脯氨酸富集区、谷氨酰胺富集区、丝氨酸-苏氨酸富集区、亮氨酸拉链、核定位结构域等。拟南芥AtWRKY7同时含有1个WRKY结构域和1个钙调蛋白CaM结合结构域。WRKY转录因子多样的结构域表明,拥有特殊结构的WRKY转录因子可以在基因表达调控中发挥重要的特殊功能作用。
3.2 结论 本研究获得了WRKY基因家族的结构特征,20个基因家族成员主要分布在12条染色体上。毛果杨WRKY基因家族20个成员编码序列长为189~624bp,有多个染色体定位重复分布于第1、5、14号染色体上。WRKY蛋白均定位于细胞外,亲水性较差,具有较高的脂溶性和不稳定性。20个WRKY基因均在毛果杨中表达,但有明显差异。从基因家族成员结构来看,毛果杨20个WRKY基因中大多数含有3个及以上的内含子,说明不同的基因有着不同的结构形式,从而它们可以用不同的方式发挥不同的作用。从毛果杨WRKY基因家族的热图来看:毛果杨WRKY基因在成叶中的转录水平最低,而在根、幼叶、雌蕊、雄蕊、木质部其他5个组织及光生苗中均具有较高的转录水平。从理化性质分析结果来看:毛果杨WRKY基因多数定位在细胞外,其不稳定指数较高,脂溶性指数较低,亲水性较低。
本研究利用多种计算机软件工具,对毛果杨WRKY基因家族进行了生物信息学的分析,对其家族成员进行了鉴定,为今后研究该类基因的功能提供了信息依据。另外,本研究对毛果杨WRKY基因家族的研究还只停留在对其基因序列等的简要分析,而要想真正了解它的相关作用,还需要在未来对各个领域进行更深入的研究。
参考文献
[1]Zhang YJ,WangLJ.The WRKY transcription factor superfamily:its origin in eukaryotes and expansion in plants[J]. BMC Evol Biol.,2005,5(1):1.
[2]Martinez E. Multi-protein complexes in eukaryotic genetranscription[J]. PlantMolBiol,2002,50(6):925-947.
[3]lshiguro S,Nakamura K. Characterization of a cDNA encoding a novel DNA-binding protein,SPF1,that recognizes SP8 sequences in the 5′ upstream regions of genes coding for sporamin and beta-amylase from sweet potato[J]. Mol Gen Genet.,1994,244:563-571.
[4]Rushton PJ,MacDonald H,Huttly AK,et al. Members of a new family of DNA-binding proteins bind to a conserved cis-element in the promoters of alpha-Amy2 genes[J].Plant MolBiol.1995,29:691-702.
[5]ushton PJ,Torres JT,Parniske M,et al. Interaction of elicitorinduced DNA-binding proteins with elicitor response elements in the promoters of parsley PR1 genes[J]. EMBO J,1996,15:5690-5700.
[6]de Pater S,Greco V,Pham K,et al. Characterization of a zincdependent transcriptional activator from Arabidopsis[J]. Nucleic Acids Res.,1996,24:4624-4631.
[7]Chen CH,ChenZX.Isolation and characterization of two pathogen-and salicylic acid-induced genes encoding WRKT DNA-binding proteins from tobacco[J].Plant Mol Biol.,2000,42(2):387-396.
[8]Lagace,M,Matton,et al. Characterization of a WRKY transcription factor expressed in late torpedo-stage embryos of Solanumchacoense[J]. PLANTA,2004,219(1):185-189.
[9]RobatzekS,Somssich IE. Targets of ArWRKY6 regulation during plant senescence and pathogen defense[J]. Genes Dev.,2002,16(9):1139-1149.
[10]Fu QT,Yu DQ. Expression profiles of AtWRKY25.AtW(_RKY26and AtWRKY33under abioticstresses[J].Hereditas(Beijing),2010,32(8):848-856.
[11]EulgemT,RushtonPJ,RobatzekS,et al The WRKY superfamily of plant transcription factors[J]. Trends Plant Sci.,2000,5(5):199-206.
[12]龐文玉,王安,杨宝谊,等.大白菜ENT基因家族的鉴定与生物信息学分析[J].江苏农业科学,2019,47(12):52-57.
[13]EulgemT,RushtonPJ,RobatzekS,et al.The WRKY superfamily of plant transcription factors[J]. Trends in Plant Science,2000,5(5):199-206.
[14]Rushton PJ,SomssichIE,RinglerP,et al.WRKY transcription factor[J]. Trends in Plant Science,2010,15(5):247-258.
[15]MaeoK,HayashiS,KojimaSH,et al.Role of conserved residues of the WRKY domain in the DNA-binding of tobacco WRKY family proteins[J]. Bioscience Biotechnology,2001,65(11):2428-2436.
[16]Dong J,Chen C,Chen Z. Expression profiles of the Arabidopsis wrky gene superfamily during plant defense response[J].Plant MolBiol.,2003,51:21-37.
(责编:张宏民)
