过表达酰基蛋白硫酯酶促进产油真菌卷枝毛霉脂肪酸合成
2021-06-18辛菲菲王瑞雪赵燕磊张怀渊
高 萌,辛菲菲,王瑞雪,赵燕磊,张怀渊,张 瑶
山东理工大学 农业工程与食品科学学院 (淄博 255000)
卷枝毛霉(Mucorcircinelloides),接合菌纲毛霉科毛霉属,通过孢子和接合子进行繁殖[1],因其具有高产油能力而被关注,特别是其油脂中含有的γ-亚麻酸(GLA)使其具有重要的经济价值[2],卷枝毛霉已经被作为研究丝状真菌脂质积累机制的模式生物。本实验室筛选到一株野生型高产油卷枝毛霉菌株WJ11,其油脂产量能够达到细胞干重的36%,卷枝毛霉基因组学、蛋白组学和酶学已经被深入研究以探索脂质积累机制[3]。
酯酰基硫酯酶(thioesterase,TE)起到终止脂肪酸链继续延长的作用,水解脂酰基-S-酰基载体蛋白(acyl-ACP)的硫酯键,释放游离脂肪酸(free fatty acid,FFA)[4]。各种各样的硫酯酶使多种底物中硫酯键的裂解而发挥各种重要的生物功能[5],因此产油微生物中的硫酯酶基因常被遗传改造以提高脂肪酸产量[6]。酰基蛋白硫酯酶(Acyl-protein thioesterase,APT)就是另一类硫酯酶,也称溶血磷脂酶和羧酸酯酶,在硫酯酶家族中被称作TE21硫酯酶,水解蛋白质上的半胱氨酸(Cys)残基和酰基链之间的硫酯键,它们存在着一个经典的α/β水解酶结构域[7]。人类APT1和APT2为最典型的TE21硫酯酶,与棕榈酰转移酶共同作用,调节棕榈酰化蛋白的细胞表面表达、生长相关蛋白和受钙离子激活的钾离子通道,通常定位在高尔基体[8]。
卷枝毛霉WJ11的基因组中有一个APT硫酯酶(evm.model.scaffold00021.24,TE21)基因,本实验旨在通过基因操作过表达卷枝毛霉WJ11的te21基因,重组菌种经发酵罐培养后测定脂质组成和总脂肪酸含量,来探究TE21在卷枝毛霉WJ11脂肪酸合成与代谢中的作用。
1 材料与方法
1.1 材料
1.1.1菌种
菌种为卷枝毛霉M65,是野生型卷枝毛霉WJ11的尿嘧啶缺陷型;构建表达载体所用大肠杆菌DH5α和质粒pMAT2075均保藏于本实验室[9]。
1.1.2试剂
Trizol试剂,购自生工生物工程(上海)有限公司;Plasmid Mini Kit I试剂盒,购自美国Omega 生物科技公司;qPCR SYBR Green Master Mix试剂盒,购自上海翊圣生物科技有限公司。
1.1.3培养基
大肠杆菌培养基采用LB培养基[3],抗生素为氨苄青霉素(Ampicillin,Amp);转化所用的培养基YPG、MMCS、YPGS等参照赵利娜等[3]方法配制;实验所用的种子培养基为K&R培养基,发酵培养基为改良的K&R培养基[10]。
1.2 方法
1.2.1pMAT2075-te21表达载体的构建
用Trizol提取卷枝毛霉WJ11的基因组总RNA,反转录得到cDNA,利用te21的引物(上游引物F1:ATGTCTCTTACTTCTGTTGTAG,下游引物R1:TTAGATGGAAGGGATTGTCT)进行PCR扩增,PCR产物凝胶电泳后胶回收,将基因片段和空质粒pMAT2075用XhoI和NheI酶切,用T4连接酶于16 ℃过夜连接后,热击转化到大肠杆菌DH5α感受态细胞,筛选获得大肠杆菌转化子,测序验证,得到pMAT2075-te21,于-80 ℃甘油管中保菌。
1.2.2卷枝毛霉WJ11的转化
用Plasmid Mini Kit I提取pMAT2075-te21质粒,加入SmaⅠ及AatⅡ酶切过夜,凝胶电泳验证后纯化得到目的基因的片段。转化所用的菌种为卷枝毛霉M65,方法参照赵利娜等[3]。
1.2.3卷枝毛霉转化子的筛选
过表达通过同源重组的方式导入卷枝毛霉基因组中,经过多代筛选直到在筛选培养基与全营养培养基上长出相同数量的单菌落,提取转化子的基因组DNA,PCR验证,引物为F2:GATAAGCATAAACCAGATCTGC R2:GTATCTGACATAGTCGAGCTTC。
1.2.4重组菌种发酵培养
摇瓶培养方法与条件参考赵利娜[3]等,在发酵的48 h和96 h取样。1.5 L发酵罐装液量为1 L,接种量为10%,转速为700 r/min,pH为6.0(2 mol/L NaOH调节),温度为28 ℃,通气量为2 L/(L·min),发酵时间为96 h,在发酵的12 h、24 h、36 h、48 h、72 h、96 h取样。取样量为20 mL,用于转录分析,生物量和油脂分析。滤液用于测定培养基中残余的葡萄糖和NH4+。葡萄糖浓度采用还原糖生物传感分析仪(SBA-40E,济南柏盛)测定,NH4+采用靛酚蓝比色法[11]。
1.2.5荧光定量PCR测定TE21的mRNA表达水平
荧光定量PCR的反转录方法同1.2.1,然后用qPCR SYBR Green Master Mix进行反转录,荧光PCR仪型号为LightCycler 96(Roche),PCR反应程序参照说明。mRNA的相对表达量使用2-ΔΔCt法分析。
1.2.6薄层层析色谱(TLC)测定脂质组成
脂质组成测定采用TLC方法参照鲍方宇[12]等。采用灰度扫描计算各组分比例。用刀片刮下显色后的条带,经10%盐酸甲醇甲酯化后用气相色谱(GC,Agilent 7890B)测定其脂肪酸组成,程序参考Khan等[10]。
1.2.7脂肪酸的提取及测定
油脂提取采用氯仿-甲醇方法,参考唐鑫[11]等的实验方法。脂肪酸组成的测定方法同1.2.6。
2 结果与讨论
2.1 过表达TE21卷枝毛霉菌株的构建
卷枝毛霉转化时以片段化的空载体作为阴性对照,得到对照菌株Mc-2075和阳性转化子Mc-TE21。提取转化子基因组为模板,对照组Mc-2075经PCR扩增后为约5 361 bp,而阳性转化子Mc-TE21为6 051 bp的条带,经电泳后可以看出Mc-2075的条带在6 000 bp以下,Mc-TE21在6 000 bp上方(图1),证实TE21基因已经整合在卷枝毛霉基因组上。
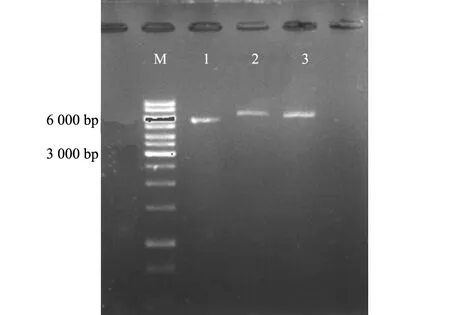
注:M(Marker)为核酸分子量标准;1为Mc-2075;2和3为Mc-TE21转化子。图1 转化子PCR凝胶电泳图
2.2 摇瓶中TE21过表达对卷枝毛霉脂肪酸含量的影响
挑选出3个卷枝毛霉转化子进行摇瓶发酵,发酵时长为96 h,3个转化子的生长状况与对照组Mc-2075并无差异(图2),这意味着过表达TE21基因对卷枝毛霉并未损伤,测定96 h时脂肪酸含量,结果显示3个转化子的总脂肪酸含量均高于对照组(图3),脂肪酸含量最高的是Mc-TE21-3,达到细胞干重的27.1%,选取Mc-TE21-3进行发酵罐培养以准确测定其对卷枝毛霉脂质积累的影响。此结果说明TE21同酯酰基硫酯酶一样也能促进对产油生物的脂质积累[13]。
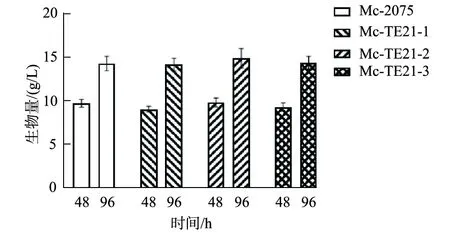
图2 转化子在摇瓶中发酵的生物量

图3 转化子在摇瓶中发酵的脂肪酸含量
2.3 发酵罐中TE21过表达对卷枝毛霉脂质的影响
2.3.1TE21过表达对mRNA表达水平的影响
TE21基因是卷枝毛霉WJ11的内源基因,荧光定量PCR用于检测TE21过表达对其转录水平的影响。重组菌种Mc-TE21-3和对照菌种在经过发酵罐培养后,6 h、24 h、72 h、96 h取样测定TE21的mRNA的表达量。与Mc-2075相比,Mc-TE21-3中TE21 mRNA的表达水平显著提高,特别是在24 h和72 h时,TE21的mRNA表达水平分别提高26.7倍和24.3倍(图4),说明TE21基因经过遗传改造后其转录水平明显增强。
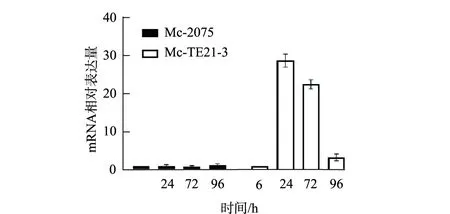
图4 对照组Mc-2075与Mc-TE21-3中TE21相对表达量
2.3.2发酵罐中生物量、残余葡萄糖和NH4+浓度的变化
转化子在发酵罐中培养时,葡萄糖和NH4+均被快速消耗,在12 h NH4+时被耗尽,此时卷枝毛霉开始将培养基中的葡萄糖转化为脂质储存于细胞内,细胞内脂肪酸含量逐渐增加。与Mc-2075相比,Mc-TE21-3的葡萄糖消耗速率稍微降低(图5a),而对NH4+的消耗速率几乎没有差别(图5b)。卷枝毛霉菌种生物量随着发酵时间的逐渐增加,而TE21过表达导致生物量稍有下降,在96 h时,Mc-2075的生物量为18.4 g/L,而Mc-TE21-3的生物量为15.8 g/L,生物量降低了14.13%。

图5 Mc-2075和Mc-TE21-3在1.5 L发酵罐中发酵时指标的变化
2.3.3过表达TE21对卷枝毛霉脂质的影响
12 h时氮源耗尽后细胞内的脂质开始快速积累[3],12~48 h之间细胞生长和脂肪酸合成的快速时期(图5c),48 h后总脂肪酸积累速度逐渐放缓并趋于稳定,与对照菌种相比,过表达TE21使得脂肪酸含量显著提高,在96 h时Mc-2075的脂肪酸含量为(25.9±1.4)%,Mc-TE21-3脂肪酸总量达到(30.0±1.4)%,整体高于Mc-2075(图5d)。TE21过表达对卷枝毛霉脂肪酸组成并没有影响(表1)。
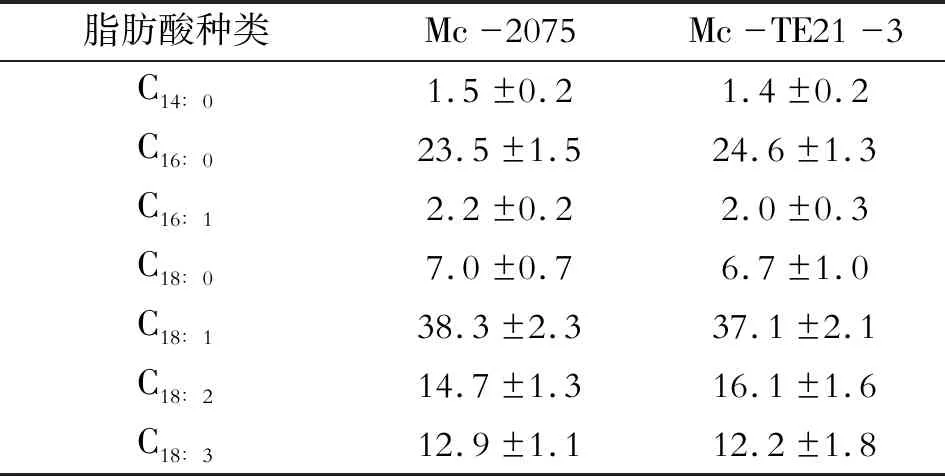
表1 发酵罐中卷枝毛霉重组菌种的脂肪酸组成 %
TLC中各种物质在薄层硅胶板上移动的距离随在展开剂中的迁移率(Rf)的不同而变化,甘油三酯(TAG)、FFA、1,3-甘油二酯(1,3-DAG)、1,2-甘油二酯(1,2-DAG)及甘油单酯(MAG)的极性依次增大,而离开原点的距离依次减少[13](图6)。在TLC硅胶板显色处理后发现,卷枝毛霉的脂质产物中TAG所占总脂质含量的比例最多,在Mc-2075和Mc-TE21-3中的比例分别为67.8%和62.2%。而Mc-TE21-3的FFA、1,3-DAG和1,2-DAG均有不同程度的提高,尤其是FFA,达到脂质组成的18.1%,与Mc-2075中FFA相比,提高了29.1%(图7)。

图6 Mc-2075与Mc-TE-3的脂质在薄层层析硅胶板显色

图7 Mc-2075与Mc-TE-3的脂质组成比例
在卷枝毛霉脂质组分的脂肪酸组成中,Mc-TE21-3与Mc-2075的TAG和MAG的脂肪酸组成没有明显差异,而过表达TE21对FFA、1,3-DAG和1,2-DAG的脂肪酸组成有较大的影响。在FFA脂肪酸组成中,过表达TE21显著提高了C16∶1的比例,而降低了C18∶1和C18∶2的比例,说明TE21对C16∶1有一定的偏好性,而不亲和C18∶1和C18∶2(表2)。对1,3-DAG和1,2-DAG脂肪酸组成的影响与FFA的结果类似。

表2 Mc-2075和Mc-TE21-3脂质中各组分脂肪酸组成 %
3 结论
APT属于硫酯酶家族中的硫酯酶TE21,其生理功能是水解脂酰基-S-酰基载体蛋白的硫酯键,释放FFA,起到催化脂肪酸链的终止作用。本研究通过过表达卷枝毛霉的TE21,使实验菌株Mc-TE21-3的脂肪酸含量达到细胞干重的30.0%,相比于Mc-2075的脂肪酸含量提高了15.9%,TE21过表达还导致了卷枝毛霉脂质的组成,显著提高了FFA的比例,且对FFA和DAG的脂肪酸组成有较大的影响。本研究证实TE21在卷枝毛霉中的生理功能,显著提高了脂肪酸含量,丰富产油微生物脂肪酸合成路径中关键酶的研究。
