Gomphonema Ehrenberg species from the Yuntai Mountains,Henan Province, China*
2021-06-15QiLIUNiniCUIJiaFENGJunpingFangruNANXudongLIUShulianXIEJohnPatrickKOCIOLEK
Qi LIU , Nini CUI, Jia FENG, Junping LÜ , Fangru NAN, Xudong LIU , Shulian XIE ,**,John Patrick KOCIOLEK
1 School of Life Science, Shanxi University, Taiyuan 030006, China
2 Museum of Natural History and Department of Ecology and Evolutionary Biology, University of Colorado, UCB 218, Boulder CO80309, USA
Abstract We report on eight species of the freshwater diatom genus Gomphonema from aquatic habitats from the Yuntai Mountains of Henan Province, China. This work represents the first detailed descriptions of the freshwater diatoms from this centrally-located province in China. Of the eight taxa considered, five are new to science and another is given a “cf.” designation. Four of the eight species are similar in morphological features to other, “classical” species, with external areolar occlusions forming C-shaped openings. Another species is astigmate, and has distinct siliceous papillae located marginally on the exterior of the valves;this morphology has not been recognized previously for the genus. Two of the species have internal areolar and stigma occlusions. The diversity of morphological types of species in Gomphonema may relate to the ecosystems, with more diverse ecosystems harboring greater diversity of these morphological groups of Gomphonema.
Keyword: Gomphonema; new species; Bacillariophyceae; morphology; diatoms
1 INTRODUCTION
Work on the taxonomy of freshwater gomphonemoid diatoms of Asia has a long history and has been enjoying a revival in the last decade or so. Descriptions of new taxa occurred during the middle and the end of the 19thcentury, with species described from the region by Ehrenberg (1854),Grunow (in van Heurck, 1880) and Brun (in Schmidt,1899). New species were described from Asia into the 20thcentury, including the work by Hustedt (1922,1927) Meister (1932), Amossé (1969), Gandhi (1958,1960), Foged (1971), Lange-Bertalot and Genkal(1999), and similar studies have continued into the 21stcentury by Kulikovskiy and colleagues(Kulivoskiy and Kociolek, 2014; Glushchenko et al.,2017) and Kociolek et al. (2015a, 2018).
In China, specifically, Skvortzow (1929, 1935,1937, 1938, 1946) documented many new gomphonemoid species from mostly the eastern part of the country. A review of gomphonemoid diatoms from China was presented by Shi (2004), who described 9 new taxa. Bao and Reimer (1992)described a new variety, and Fan et al. (2004)described two new taxa, all from northeastern China.Liu et al. (2013) described six newGomphonemaspecies from the same region within China. Li and her colleagues (Li et al., 2006, 2010; Gong and Li, 2012),as well as Jiang et al. (2018) documented new gomphonemoid diatoms from Yunnan Province, and You et al. (2015) documented new species from Xinjiang Uygur Autonomous Region, China.Kociolek et al. (2015b) described a new genusGomphosinicaof gomphonemoid diatom from China and an additional species was subsequently described from Yunnan Province (Cheng et al., 2018).

Fig.1 Sampling sites of Yuntai Mountains, Henan Province, China; Digital Elevation Model (DEM)
The purpose of the present report is to use light and scanning electron microscopy to report on eightGomphonemaspecies from the Yuntai Mountains of Henan Province, China, five of which are new to science.
2 MATERIAL AND METHOD
2.1 Collection
Diatom samples were collected during an expedition to the Yuntai Mountains, Xiuwu County,Henan Province, China. Five samples were noted to contain new or interesting species (Fig.1). These are described below:Samples:
201805021-1: Henan Province, Yuntai Mounweitains, Tanpu valley, river, periphyton, China(35°27′31′′N, 113°20′50″E), there were fourGomphonemaspecies identified in this sample,Gomphonemalanceolatoidessp. nov.,Gomphonemamontanavivasp. nov.,Gomphonemapseudintricatumsp. nov. andGomphonemavibrio.
201805036-1: Henan Province, Yuntai Mountains,Tanpu valley, wet wall, periphyton, China(35°30′10″N, 113°25′4″E), this sample contained twoGomphonemaspecies,GomphonemalateripunctatumandGomphonemayuntaiensissp. nov.
201805045-1: Henan Province, Yuntai Mountains,Tanpu valley, wet wall, periphyton, China(35°29′54′′N, 113°25′40″E), there is only oneGomphonemaspecies appear in this sample,Gomphonemapseudosphaerophorum.
201805046-1: Henan Province, Yuntai Mountains,Tanpu valley, wet wall, benthic, China (35°29′54′′N,113°25′40″E), a singleGomphonemaspecies,Gomphonemafortissimo.
201805050-1: Henan Province, Yuntai Mountains,Tanpu valley, wet wall, periphyton, China(35°29′34′′N, 113°26′9″E), there were twoGomphonemaspecies in this sample,Gomphonemalanceolatoidessp. nov.,Gomphonemafortissimo.Surface water temperature and pH were measured using a HANNA HI 98128 pH meter.
2.2 Sample processing
Samples were digested with hydrochloric acid to remove organic matter by cooked. After repeated dilutions of the treated material with distilled water,aliquots were dried on cover glasses and mounted in Naphrax (Brunel Microscopes Ltd., Chippenham,UK). All Light microscopy (LM) observations were performed using an Olympus BX-51 light microscope with a 100×oil immersion objective (numerical aperture 1.40). Specimen images were taken using an Olympus DP72 digital camera at Shanxi University,China.
For Scanning electron microscopy (SEM)observations, the cleaned specimens were air-dried onto glass cover slips, attached to copper stubs, and coated with approximately 15 nm of gold-palladium.Scanning electron microscope observations were made using a Hitachi SU8010 SEM at a working distance less than 6mm and accelerating voltage set at 2 kV at Shanghai Normal University, China.
Holotype slides and material are stored at the Q.LIU Diatom Collection in the Freshwater Algal Herbarium of Shanxi University, China. Isotype slides are deposited in the Kociolek Collection at the University of Colorado (COLO), USA.
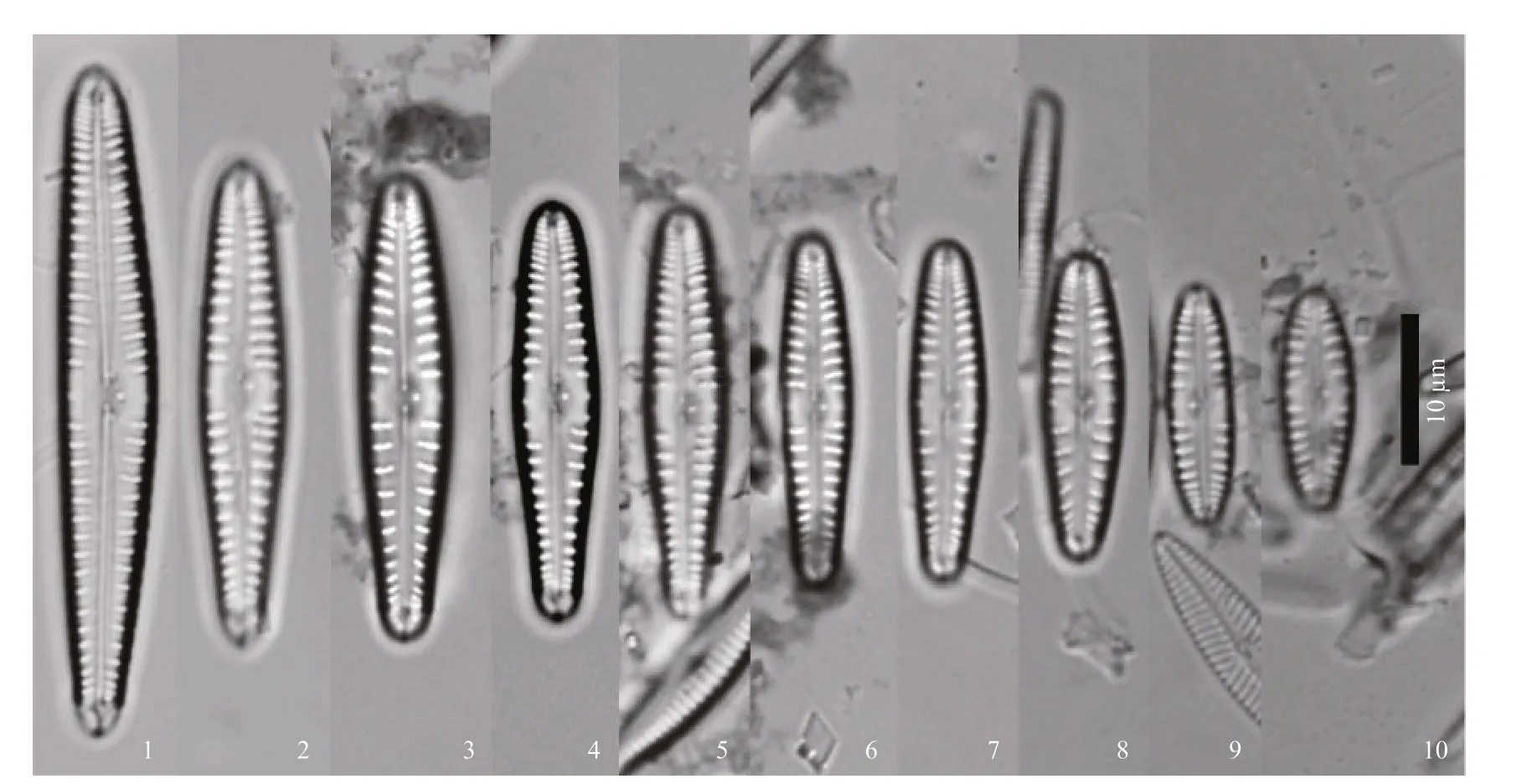
Figures 1–10: LM, valve views, showing size diminution series.
3 RESULT
Gomphonemacf.lateripunctatumReichardt &Lange-Bertalot 1991 Figures 1–18.
Description: valves liner-lanceolate-clavate, with rounded poles. Length 15–45 μm, breadth 4.5–6.0 μm. Axial area narrow, widening towards the center, expanded laterally to form a wide,asymmetrical, nearly rectangular central area. One short stria is positioned on one side of the central area,the side bearing the stigma. A single, round stigma is located in the central area, near the proximal raphe ends. Raphe straight or weakly undulate, lateral, with distinct, dilated external proximal raphe ends,recurved internal raphe ends. Striae indistinctly punctate, parallel to radiate most of the length of the valve, strongly radiate at the poles. Striae density (8–9)/10 μm in the center of the valve, condensed at both the headpole and footpole. Septa and pseudosepta are distinct. A bilobed apical pore field (APF) is evident at the footpole.
SEM observations: externally, the areolae are round to ellipsoidal in shape, and without occlusions(Figs.11–14). At the headpole, striae are more condensed than at the center of the valve (Figs.11,12), and the distal raphe end is deflected before extending onto the valve mantle (Fig.12). Striae extend from the valve face onto the mantle. In the center of the valve, the stigma opening is round and similar in size and shape to the areolae. Proximal raphe ends are dilated (Fig.13). At the footpole, the raphe bends between the two lobes of the APF. Porelli of the APF are of physically separated from the areolae, but resemble them in overall size and shape(Fig.14). The APF extends onto the valve face, but is mostly on the mantle.
Internally, there is a distinct central nodule and pseudosepta at the poles (Figs.15–18). The raphe appears slightly undulate, with proximal ends recurved on the central nodule. Thee raphe terminates distally as prominent helictoglossae (Figs.15–18).Areolae occur in troughs between thick interstriae,and are occluded with round hymenes (Figs.15–18).The stigma is also occluded internally.
Comments: Our specimens match up well with some interpretations of this taxon, such as those presented by Delgado et al. (2013) from Spain and Jüttner (2018) from Scotland. It is reported from alpine areas in Italy (Cantonati et al., 2012), Austria(Trobej et al., 2017) and in Nepal (Jüttner et al., 2018),though the latter population has a more linear outline than the specimens we encountered from the Yuntai Mountains.
Our designation of these specimens as “cf.”involves uncertainty around the original description of the taxon by Reichardt and Lange-Bertalot (1991).Specimens included in the original description appear to represent several taxa (compare the shapes and striae densities of their Figs.8 & 9, which appears to be a separate taxon from what might be considered the commonly reported morphology in their Figs.3 &12). The original description reported breadth up to 14 μm, and striae densities up to 14/10 μm, but subsequent reports of breadth are up to 6 μm (here) or lower, and striae densities of (8–12)/10 μm.
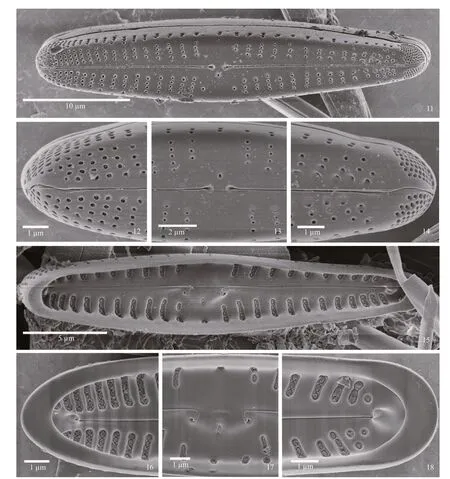
Figures 11–18: SEM; Figs.11–14: external views; Fig.11: entire valve, showing the straight raphe with dilated proximal ends and rounded areolae without occlusions; Fig.12: headpole, with round areolae, striae ending onto the mantle and deflected distal raphe end evident; Fig.13: center of the valve, showing expanded central area, dilated proximal raphe ends and small, round stigma opening; Fig.14: footpole has an apical pore field that is separated from the striae but the porelli resemble areolae in size and structure; Figs.15–18: internal views; Fig.15: whole valve with large central nodule, helictoglossae and pseudosepta evident; Fig.16: headpole, with prominent helictoglossa and pseudoseptum shown. Areolar occlusions are evident; Fig.17: central area, showing stigma opening and areolae with occlusions. Proximal raphe ends are recurved at each end of the central nodule; Fig.18: footpole, with helictoglossa and wide pseudoseptum present. Areolar occlusions are evident.
Gomphonemayuntaiensissp. nov.
Figures 19–36. Figure 21 is of the holotype.
Description: Valves elliptical-lanceolate-clavate,with rounded apices. Length 25–70 μm, breadth 6–12 μm. Axial area narrow, widening to form an irregular to rectangular, laterally-expanded central area with one-two shortened, more coarsely-spaced striae on either side of the central area. Stigma is wanting. Striae slightly arched, lateral. External proximal raphe ends slightly dilated, external distal raphe ends deflected onto the mantle. Internal distal raphe ends evident, recurved before the external ends.Striae punctate, radiate, (9–12)/10 μm near the center of the valve, (10–14)/10 μm at the apices. A distinct,large bilobed apical pore field is present at the footpole. Distinct septa and pseudosepta are present.

Figures 19–28: LM, valve views, showing size diminution series. Figure 21 is of the holotype.
SEM observations: Externally, the valve face appears to have siliceous papillae on both sides (see arrow) of the axial area which are especially evident near the margins (Figs.29–32). At the headpole, the separation between the areolae towards the margin versus closer to the axial area is evident. The distal raphe end curves sharply onto the mantle and towards one side. Striae appear disorganized on the mantle.Thickened projections are present near the margins(Fig.30). At the center of the valve the proximal raphe ends are dilated and deflected slightly in the same direction. Areolae are ellipsoidal and occur in depressions. Between areolae near the margins,relatively thick siliceous projections are evident.Areolae of the striae are more separate from another near the margins than towards the central area(Fig.31). At the footpole striae have areolae interrupted, and on one side the raphe opening the slit-like areolae appear to occur in a common groove or trough. The raphe bisects the apical pore field,deflecting before reaching the APF. Porelli comprising the pore field are rounded, being morphologically distinct from the areolae. Thick marginal siliceous projections are evident at the margins (Fig.32).
Internally, the valve has large pseudosepta and helictoglossae at the poles, and an elongated central nodule (Fig.33). At the headpole, small, teeth-like projections are seen to form the serrated edges of the alveoli. The large helictoglossa is off set from the raphe slit. The pseudoseptum is evident (Fig.36). At the central area, the elongated, raised central nodule bears the recurved proximal raphe ends. Alveoli of the striae have serrated edges. Individual openings appear round, with a thin siliceous covering (Fig.35).The footpole bears a pseudoseptum much smaller than at the headpole. The helictoglossa is off set from the raphe slit. Porelli of the pore fields are occluded(Fig.34).
Holotype: 201805036-1, holotype illustrated in Fig.21. Isotype JPK Collection, COLO, slide number to be indicated upon acceptance of manuscript.
Type locality: Henan Province, Yuntai Mountains,Tanpu valley, wet wall, periphyton, China(35°28′28′′N, 113°22′39″E).
Etymology: Named for the mountains in which it occurs.
GomphonemapseudosphaerophorumH. Kobayasi in Ueyama & Kobayasi 1988
Figures 37–50.
Description: valves elliptical-lanceolate clavate,with headpole broadly capitate and rounded. Length 32–45 μm, breadth 8–9 μm. Axial area narrow at the poles, narrowly lanceolate towards the center, then laterally expanded to form a narrowly-rectangular central area. There is an isolated stria on either side of the central area, one terminating near the center with a round stigma, on the other side striae not reaching the center. Raphe undulate, lateral, with external proximal ends dilated slightly. Striae indistinctly punctate, radiate through the valve becoming parallel towards the headpole. Striae number (9–12)/10 μm. A distinct, bilobed apical pore field is present at the footpole. Septa and pseudosepta are present at the poles.
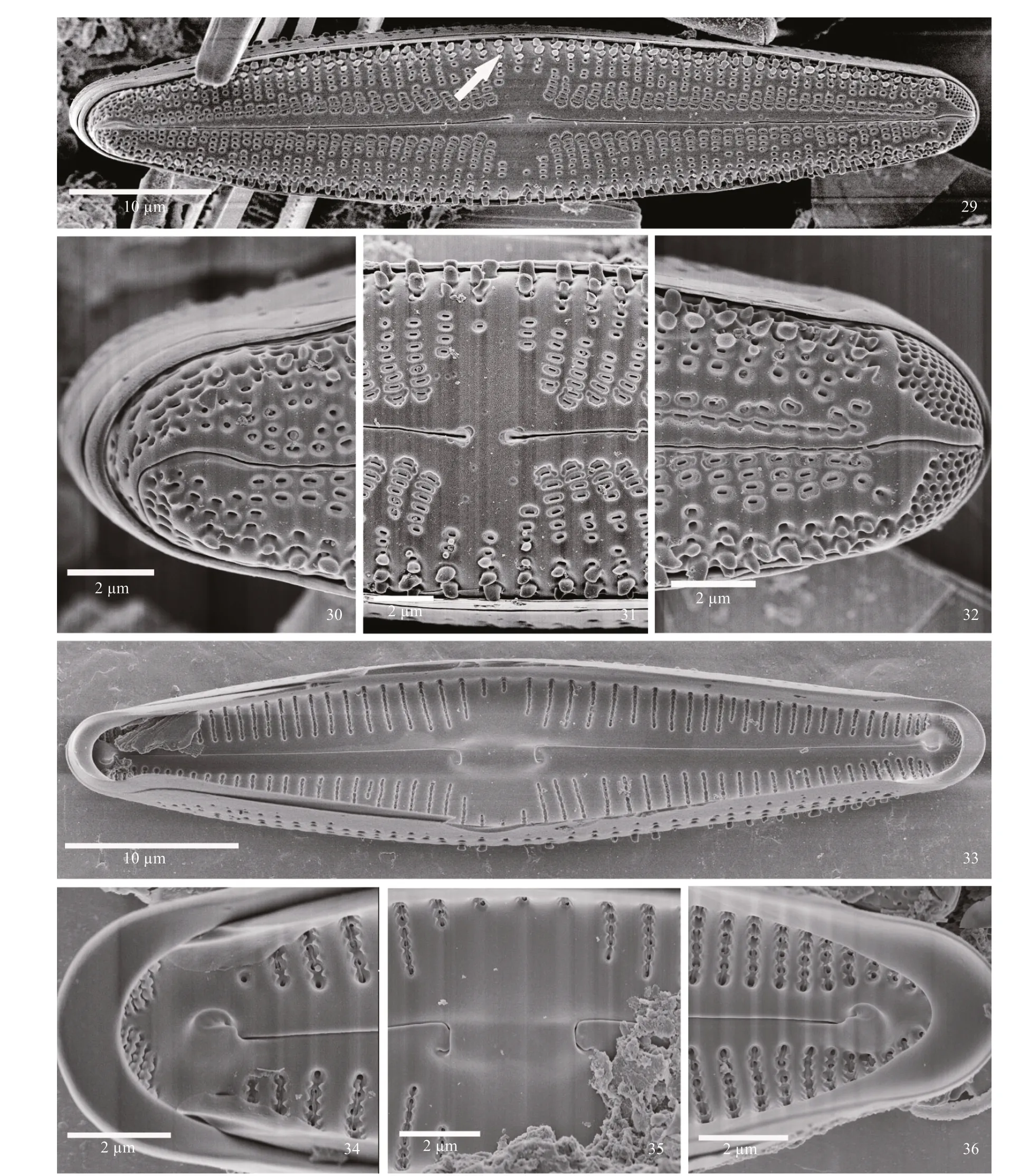
Figures 29–36: SEM; Figs.29–32: external views; Fig.29: whole valve, showing outline, raphe with proximal end curved the same direction, APF and evident papillae around the edge of the valve; Fig.30: headpole, with hooked distal raphe end; Fig.31: central area with raphe ends dilated slightly and areolae with thickened rims. Marginal papillae are shown; Fig.32: footpole, with bent distal raphe end, areolae close to the raphe slit and APF porelli that are physically separate and morphologically distinct from areolae; Figs.33–36: internal views; Fig.33: entire valve view, with prominent central nodule, and large helictoglossae and pseudosepta at the poles; Fig.34: footpole, showing striae in troughs with small, peg-like protuberances. The helictoglossa is robust, and the pseudoseptum is prominent. Porelli are occluded; Fig.35: central nodule with proximal raphe ends hooked and placed at the base of the central nodule;Fig.36: headpole, with striae, helictoglossa and septum evident.

Figures 37–45: LM, valve views, showing size diminution series.

Figures 46–50: SEM; Figs.46–49: external views; Fig.46: entire valve, with undulate raphe, C-shaped areolae and small round stigma positioned near a central stria in the central area; Fig.47: headpole, with narrowly-lunate areolar openings and raphe deflected onto mantle; Fig.48: central area showing dilated proximal raphe ends radiate striae and round stigma opening; Fig.49: footpole, with strongly radiate striae and APF porelli physically separate and morphologically distinct from areolae; Fig.50: internal view. Entire valve with large central nodule and pseudosepta evident.

Figures 51–62: LM, valve views, showing size diminution series. Figure 52 is of the holotype.
Externally, the undulate raphe is evident (Figs.46–49). At the apices the raphe extends onto the mantle at the headpole (Fig.47) and bisecting the APF at the footpole (Fig.49). The APF is composed of round porelli, which are physically separate and morphologically distinct from the areolae (Fig.49). At the central area of the valve, the proximal raphe ends are dilated. There is a round stigma opening on one side of the central area, quite distinct in size and shape from the narrow, lunate areolae (Fig.48). The uniseriate striae are composed oflineate areolae near the margins, and C-shaped or lunate areolae near the axial area (Figs.46–49).
Internally, there is a small, barely-distinguished central nodule (Fig.50). Distally, the raphe terminates as helictoglossae. On the central nodule the stigma opening is slit-like. Striae bear thickenings that separate the areolae and connect between the robust interstriae. At the headpole and footpole pseudosepta are distinct.Comments: This species is known throughout Asia(Ueyama and Kobayasi, 1988; Wang et al., 2000), and has more recently been documented from Hawaii(Miscoe et al., 2016).
Gomphonemalanceolatoidessp. nov.
Figures 51–70. Figure 52 is of the holotype.
Description: valves linear-lanceolate to lanceolateclavate, with rounded apices. Length 29.5–60.0 μm,breadth 6.5–8.5 μm. Axial area narrow, straight,expanded slightly towards both margins to form an indistinct central area. An isolated stria on one side of the central area is elongated and ends at a distinct,round stigma. The striae on the other side not reaching the center. Raphe is weakly lateral and undulate.External proximal ends indistinct. Striae indistinctly punctate, radiate near the center, become parallel towards the headpole and footpole but radiate again at the footpole. Striae number (9–11)/10 μm. Apical pore field small, bilobed. Septa and pseudosepta present at the poles.
SEM observations: Externally, the valve face is dominated by the areolae with C-shaped occlusions which, in some specimens, appear as though they have additional silica on them (Figs.63–66). The raphe is undulate, with proximal raphe ends dilated slightly, and distal raphe ends deflected onto the valve margin (Figs.63–66). The stigma opening is round,positioned just off a central stria (Fig.65). At the footpole, the raphe bisects the APF, which is almost entirely situated on the mantle. The APF is composed of round porelli, physically diff erentiated and separated from the areolae.
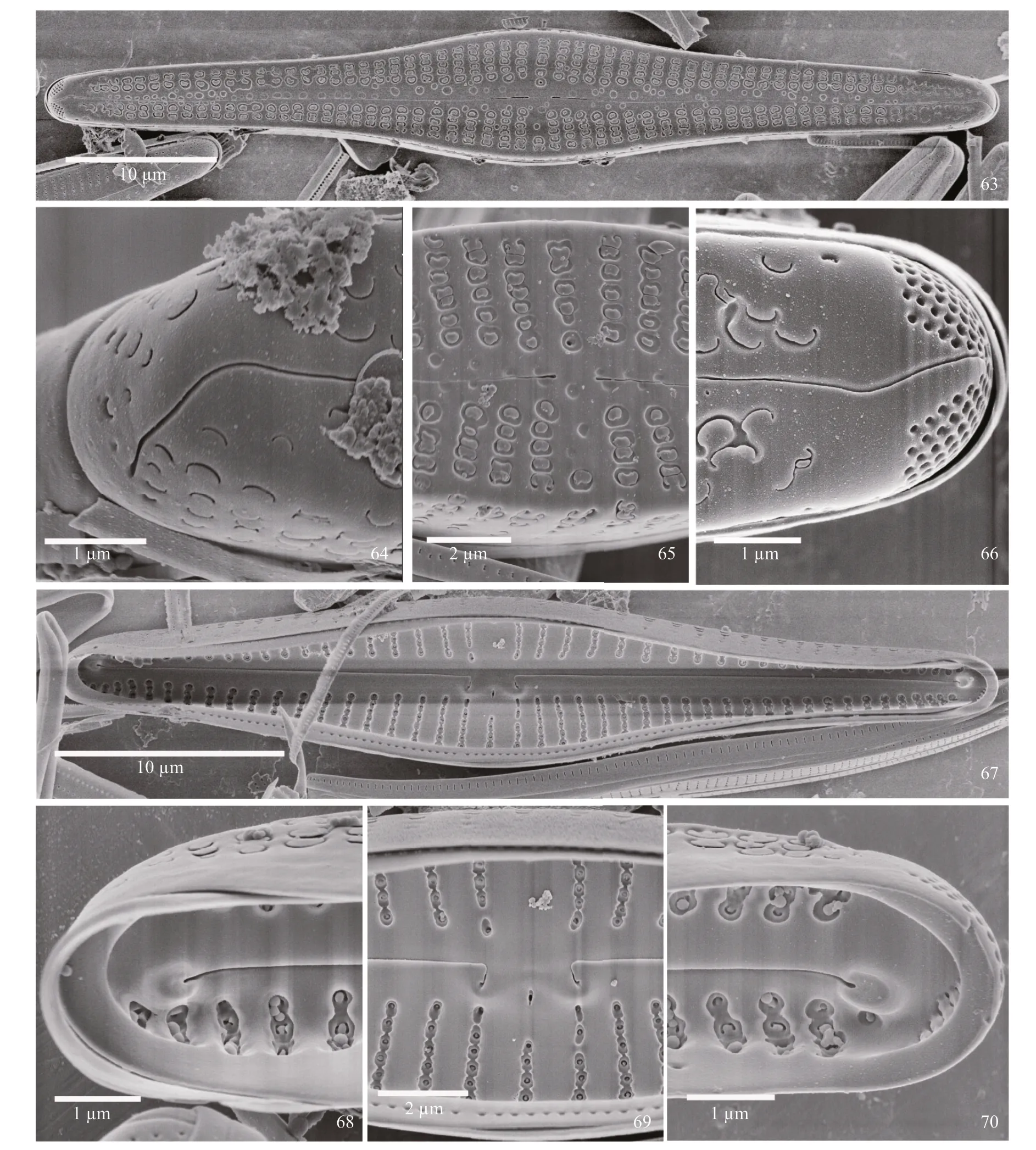
Figures 63–70: SEM; Figs.63–66: internal views; Fig.63: entire valve, showing outline and C-shaped areolae with externally-raised flaps; Fig.64: headpole,showing distal raphe end curved onto the valve mantle; Fig.65: central area with striae composed of areolae with occlusions. Areolae and small, round stigma opening appear in depressions. The proximal raphe ends are dilated slightly; Fig.66: footpole, with raphe curved on the valve face and continuing onto the mantle, bisecting the APF. APF more or less positioned on the mantle only. Porelli are distinct, physically removed and morphologically diff erentiated, from the areolae; Figs.67–70: internal views; Fig.67: whole valve with distinct, bilobed central nodule evident. Striae occur in troughs, and the stigma is positioned on one side of the base of the central nodule. Pseudosepta are helictoglossae are evident at the poles; Fig.68: headpole. Helictoglossa is off set from main axis of the raphe. Areolae appear to have internal occlusions; Fig.69: central area, with central nodule bearing recurved proximal raphe ends and stigma opening that is round-ellipsoidal in shape. Troughs with small pegs protruding into them, sometimes connecting from the sides. Areolae have occlusions; Fig.70:footpole, with pseudoseptum and helictoglossa that is off set from the main raphe axis are prominent. APF porelli appear occluded as do areolae.
Internally, the apices bear septa and pseudosepta(Figs.67, 68, 70). The position of the helictoglossae is off set from the raphe branch. The central nodule is undulate and bears the recurved proximal raphe ends.The stigma opening is small and rounded (Fig.69).Siliceous peg-like growths can be seen in the striae,which occur in deep troughs. The external occlusions of the areolae can be seen from the valve interior(Figs.68–70).
Holotype: 201805021-1, holotype illustrated in Fig.52. Isotype JPK Collection, COLO, slide number to be indicated upon acceptance of manuscript.
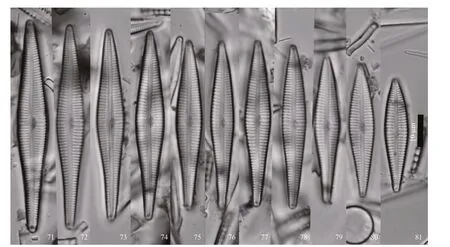
Figures 71–81: LM, valve views, showing size diminution series. Figure 77 is of the holotype.
Type locality: Henan Province, Yuntai Mountains,Tanpu valley, river, periphyton, China (35°27′31′′N,113°20′50″E).Etymology: named for the similarity in valve shape toG.lanceolatumC. A. Agardh.
Gomphonemamontanavivasp. nov.
Figures 71–89. Figure 77 is of the holotype.
Description: valves lanceolate-clavate, with apices rounded. Length 42–70 μm, breadth 8–9 μm. Axial area linear-lanceolate, with central area barely distinguished. Striae in the center of the valve slightly coarser than the rest of the valve, with one side of the central area with a short stria, the other side with a more elongated stria. A single round stigma opening is present at the end of one of the elongated central striae. Raphe lateral, arched. External proximal raphe ends dilated slightly. Striae punctate, radiate. Striae number (8–10)/10 μm at the center of the valve, (11–15)/10 μm at the spices. Bilobed apical pore field present at the footpole. Small septa and pseudosepta present at the poles.
SEM observations: externally, the elongated valves have an undulate raphe and distinct, round stigma(Fig.82). At the headpole, striae are seen to extending around the pole onto the mantle. The distal raphe end is deflected onto the mantle. C-shaped areolae are prominent in the striae (Fig.83). In the center of the valve, striae are composed on C- or E-shaped areolae,formed by external occlusions. Proximal raphe ends are dilated slightly, and the stigma opening is round(Fig.84). At the footpole, the distal raphe end is strongly deflected onto the mantle, bisecting the apical pore field. The bilobed pore field is comprised on round porelli, organized into discreet rows. Porelli are morphologically distinct from the C-shaped areolae and physically separated from them (Fig.85).
Internally, the frustules can be seen to have septa(Fig.87) and pseudosepta (Figs.86 & 89). At the headpole the helictoglossa is off set from the raphe branch. The small tooth-like projections can be seen at the headpole (Fig.87). At the center of the valve the small central nodule bears recurved proximal raphe ends and the slit-like opening of the stigma. Alveoli of the striae have thin, peg-like projections, and many fused to form distinct squarish compartments (Fig.88).At the footpole, the helictoglossa is aligned with the raphe branch, and porelli of the pore field are occluded(Fig.89).
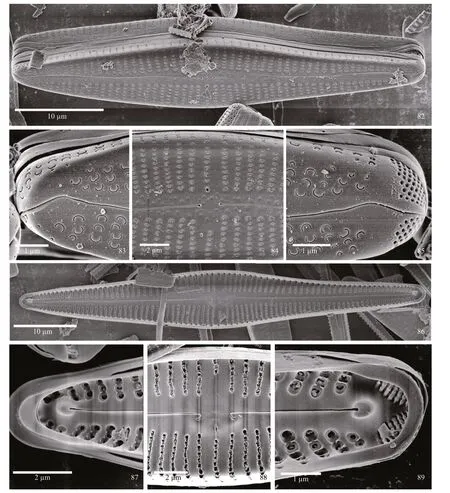
Figures 82–89: SEM; Figs.82–85: external views; Fig.82: whole frustule, with the shape of the valve and undulate nature of the raphe evident; Fig.83:headpole. Areolae are C-shaped with occlusions and the distal raphe end bends onto the valve mantle; Fig.84: central portion of the valve has parallel to slightly radiate striae, a distinct, round stigma opening and proximal raphe ends that are dilated slightly; Fig.85: footpole, with striae strongly radiate bearing C-shaped areolae. The APF is positioned on the mantle and has round porelli; Figs.86–89: internal views; Fig.86: entire valve view. The central nodule is small and rounded and helictoglossae and pseudosepta are present at the poles; Fig.87: headpole, with both septum and pseudoseptum evident. The helictoglossa is prominent, and off set from the main axis of the raphe. Small, peg-like projections are evident in the etroughs bearing the striae; Fig.88: central portion of the valve, with small round central nodule with recurved proximal raphe ends and slit-like stigma present; Fig.89: footpole, with pseudoseptum and helictoglossa evident. APF porelli appear occluded. Troughs of the striae have small, peg-like projections.
Comments: this taxon appears similar to some treatments ofG.gracileC. G. Ehrenberg.Gomphonemagracilehas been treated in mostly a broad sense (see Germain, 1981; Krammer and Lange-Bertalot, 1986, 1991), with populations having relatively wide and relatively narrow proportions given the same name. Reichardt (2015) designated a Lectotype from Ehrenberg’s collection, and specimens from that gathering have headpoles that are more rounded and possibly more similar toG.dichotomumKützing. Reichardt suggested that diatoms more similar to the commonly-applied concept ofG.gracilecan be assigned to the nameG.naviculoidesW.Smith, a form that is nearly symmetrical to the transapical axis (as its name implies), which diff ers fromG.montanaviva. Reichardt (2015) illustrated Smith’s type, and suggested this species can be found in the tropics and North America, but that specimens conforming to the type are rare in Europe. Reichardt(2015) also described some new species that might have previously be confused with G. gracile sensu lato, includingG.graciledictumReichardt, a diatom with an overall shape similar toG.montanaviva, but where the vegetative valves are distinctly smaller(20–53 μm long, 6.0–7.5 μm wide inG.graciledictumas compared to 42–70 μm long, 8–9 μm wide inG.montanaviva). The longest valves illustrated by Reichardt for his taxon look quite unlike the smallest valves ofG.montanaviva, even though they are the nearly the same size.

Figures 90–101: LM, valve views, showing size diminution series. Figure 96 is of the holotype.
Holotype: 201805021-1, holotype illustrated in Fig.77. Isotype JPK Collection, COLO, slide number to be indicated upon acceptance of manuscript.
Type locality: Henan Province, Yuntai Mountains,Tanpu valley, river, periphyton, China (35°27′31′′N,113°20′50″E).Etymology: named for living in mountainous regions.
Gomphonemapseudintricatumsp. nov.
Figures 90–109. Figure 95 is of the holotype.
Description: valves linear-clavate, margins nearly straight, with apices rounded. Length 28.0–50.5 μm,breadth 5–8 μm. Axial area narrow at the poles,lanceolate in shape until becoming the rectangular central area with a single shortened stria on either side. Raphe straight, lateral, with dilated external proximal raphe ends and distinctly recurved internal proximal raphe ends. A single round stigma opening is positioned near the external proximal raphe ends.Striae indistinctly punctate, radiate, (9–11)/10 μm,striae condensed around the headpole. A distinct,bilobed apical pore field is located at the footpole.Septa and pseudosepta are present at the poles.
SEM observations: externally, each branch of the raphe is seen to be arched, with the proximal ends dilated and rounded and the distal ends weakly deflected and extending onto the valve mantle(Figs.102–105). A round stigma of approximately the same size as the areolae, is positioned near the proximal raphe ends in the central area of the valve(Figs.102 & 104). Striae are composed of rounded areolae that are without occlusions (see arrow)(Figs.103–105). Striae extend from the valve face to the mantle. At the headpole, striae are more condensed,and the areolae appear more lineate in shape (Fig.103).At the footpole, the raphe bisects the APF, which extends from the valve face onto the mantle (Fig.105).Porelli of the APF are physically separated and morphologically distinct from the areolae (Fig.105).
Internally, the central nodule is evident and the helictoglossa at each apex is quite large (Figs.106,107, S109). The central nodule bears the recurved proximal raphe ends. The stigma is covered by an occlusion, and the areolar openings are also occluded internally (Fig.108). A prominent septum and pseudoseptum is present at each apex (Figs.107 &109).
Holotype: 201805021-1, holotype illustrated in Fig.95. Isotype JPK Collection, COLO, slide number to be indicated upon acceptance of manuscript.

Figures 102–109: SEM; Figs.102–105: external views; Fig.102: entire valve view, showing linear-clavate outline, raphe branches that are slightly arched and have dilated proximal ends. APF is evident at the footpole; Fig.103: headpole, with lineate-like striae. Distal raphe end extends onto the valve mantle;Fig.104. Central part of the valve with slit-like or ellipsoidal areolae present. Proximal raphe ends are dilated and are positioned close to one another. A small, round stigma opening is evident; Fig.105: footpole. Raphe bisects the APF. Porelli are physically separated from the areolae, and morphologically distinct from them; Figs.106–109: internal views; Fig.106: entire valve view with distinct pseudosepta and helictoglossae at the poles; Fig.107: septum and pseudoseptum are evident, nearly covering the helictoglossae; Fig.108: central nodule, with recurved proximal raphe ends. The small round stigma opening is occluded. Areolae are also occluded internally; Fig.109: footpole. Septum and pseudoseptum are present. The helictoglossa is off set from the axis of the raphe.
Type locality: Henan Province, Yuntai Mountains,Tanpu valley, river, periphyton, China (35°27′31′′N,113°20′50″E).
Etymology: the name refers to the close resemblance of this toGomphonemaintricatum.
Comments: this species is diff erentiated fromG.vibrioandG.intricatoidesYou & Kociolek (in You et al., 2015) by the narrower, linear outline of the valve.Gomphonemavibrio(see below) is also missing the internal areolar and stigma occlusions found inG.pseudintricatum.
GomphonemavibrioEhrenberg 1843
Figures 110–126.
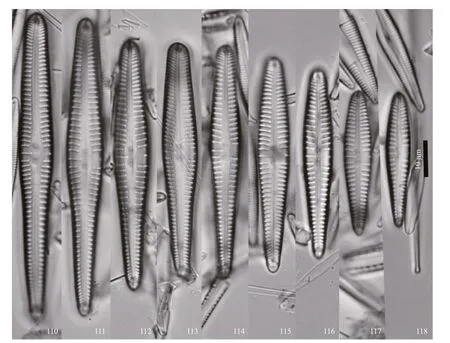
Figures 110–118: LM, valve views. Size diminution series.
Description: valves linear-lanceolate-clavate, with broadly rounded headpole and rounded footpole.Length 53–122 μm, breadth 8–13 μm. Axial area narrow at the poles, widening slightly and straight the rest of the length of the valve. Raphe straight, slightly lateral. External proximal raphe ends straight, slightly dilated; distal raphe ends bent in the same direction.Internal proximal raphe ends distinct, recurved.Central area comprised of striae more coarsely separate than the rest, a single stria on either side of the central area. On one side the stria is elongated and terminates at a small, round stigma; on the other side the stria is shortened. Stria are punctate, radiate,strongly so are the poles. Striae number (7–9)/10 μm most of the length of the valve, (11–13)/10 μm at the poles. A distinct bilobed apical pore field is present at the footpole. Septa and pseudosepta are present,distinct.
SEM observations: externally, valves linearlanceolate clavate, with a relatively straight raphe, the areolae are rounded without occlusions, and apical pore fields are prominent (Fig.119). At the headpole the raphe is weakly deflected and extends onto the mantle. Striae are denser than at the center of the valve, and the areolae are oval to ellipsoidal (Fig.120).At the center of the valve the proximal raphe ends are distinctly dilated. The stigma opening is round. Silica bordering the raphe opening may have wave-like edges (Fig.121). At the footpole, areolae may be in small, semi-circular depressions near the raphe slit or flat on the face of the valve. The distal raphe end is weakly deflected and bisects the apical pore field and extends onto the mantle. Porelli of the apical pore field are round, being physically separated from the areolae and slightly morphological diff erentiated from them (Fig.122).
Internally, the swollen central nodule, and pseudosepta at the poles are distinct (Fig.123). At the headpole (Fig.124), the pseudoseptum is large; the helictoglossa is distinct and more or less aligned with the raphe. At the valve center, the elongated central nodule bears the recurved proximal raphe ends. The stigma opening is occluded. Alveoli have smaller compartments or depressions formed by small projections between the areolae within each striae. A few examples of areolar occlusions are evident(Fig.125). At the footpole (Fig.126) the helictoglossa is off set from the raphe slit and the pseudoseptum is large. Porelli are occluded internally.
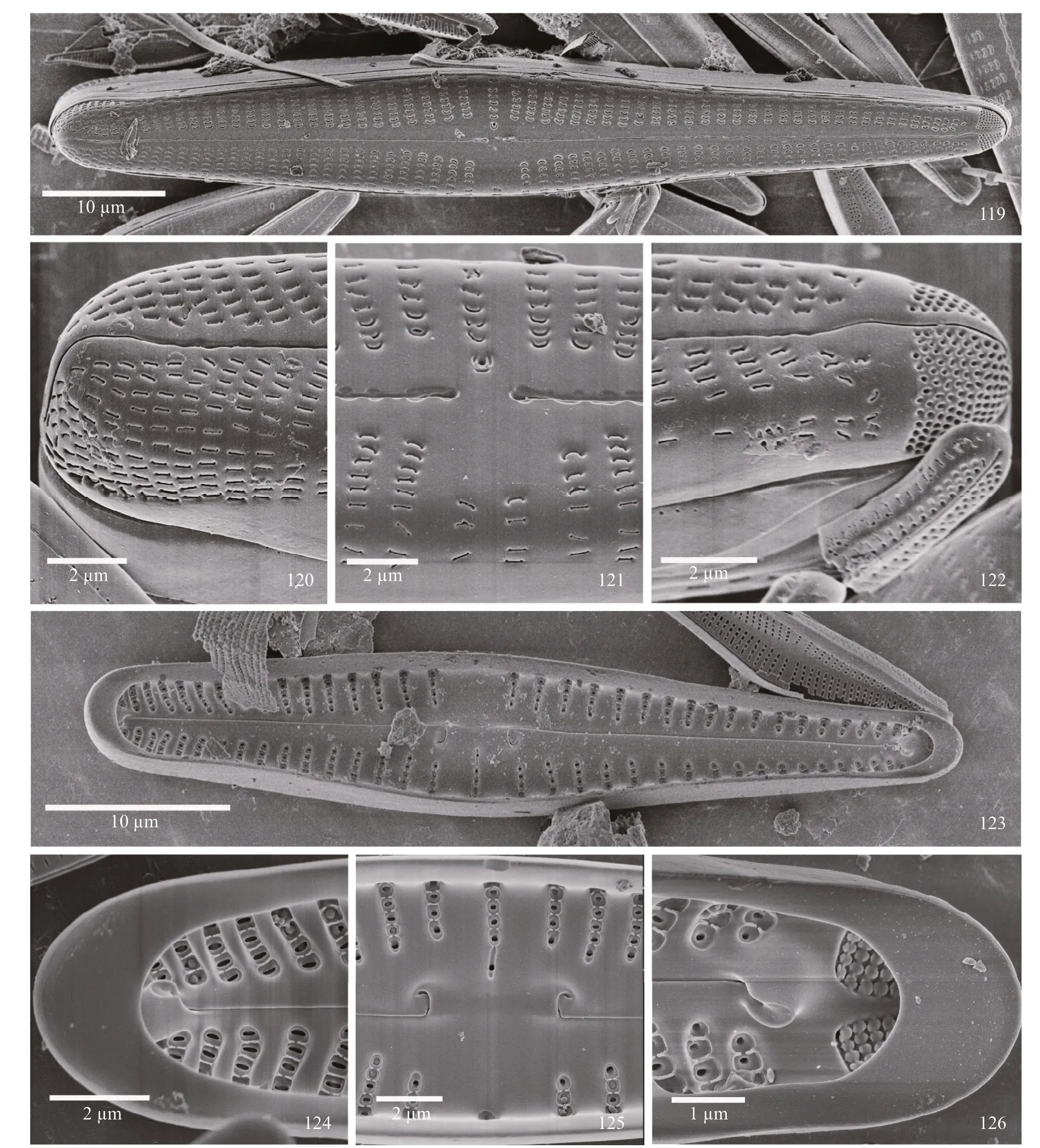
Plate 14 Gomphonema vibrio Ehrenberg
Comments: the internal occlusions of the areolae easily assign this taxon to the “Intricatum” group ofGomphonema(Kociolek and Stoermer, 1993; Jüttner et al., 2018).
Gomphonemafortissimosp. nov.
Figures 127–143. Figure 128 is of the holotype.
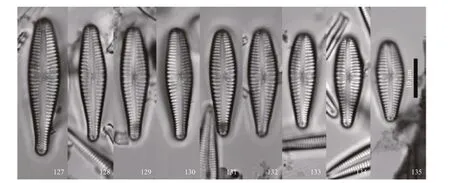
Plate 15 Gomphonema fortissimo sp. nov.
Description: valves lanceolate-clavate, with headpole indistinctly protracted and broadly rounded;footpole rounded. Length 21–47 μm, breadth 5–8 μm.Axial area narrow, widening slightly towards the center to form an irregular central area. A single stria,somewhat shorter than the others, is on either side on the central area. A single round stigma opening is located to one side of the valve center, near the proximal raphe ends. Raphe undulate, lateral for most ofits length. Striae radiate near the valve center,parallel towards and at both apices. Striae number(9–12)/10 μm. A small apical pore field is present at the footpole. Septa and pseudosepta are present at the poles.
SEM observations: valves distinctly clavate, with wide headpole and rounded footpole. Externally, the valve has a distinctly undulate raphe, with proximal ends dilated and distal ends strongly curved onto the mantle (Fig.136). At the headpole, the areolae are slitlike to C-shaped, and the distal raphe end can be seen curving at an approximately 45-degree angle onto the mantle. Striae are continuous about the headpole(Fig.137). In the center of the valve the dilated proximal raphe ends and rounded stigma are evident.Areolae comprising the striae are C-, E- or S-shaped(Fig.138). At the footpole, the striae are separated near the margin, with slit-like striae near the margin and C-shaped striae closer to the axial area. The raphe is curved, and strongly deflected on the mantle bisecting the apical pore fields. Porelli of the apical pore fields are positioned more on the mantle than the valve face. Porelli are round, morphologically diff erentiated from the areolae and physically separated from them (Fig.139).
Internally, the hemispherical central nodule extends to one side of the raphe, bearing recurved proximal raphe ends and a slit-like stigma opening. Striae are contained in alveolar grooves or troughs, without any occlusions evident. Areolar openings are round(Figs.140 & 142). At the headpole, the helictoglossa is aligned, not off set, from the raphe slit. A narrow pseudoseptum is present (Fig.141). At the footpole,the apical pore field is seen to be asymmetrical with one group of pores larger than the other. The porelli are occluded. The helictoglossae is slightly off set from the raphe slit, and the pseudoseptum is small(Fig.143).
Holotype: 201805046-1, holotype illustrated in Fig.128. Isotype JPK Collection, COLO, slide number to be indicated upon acceptance of manuscript.
Type locality: Henan Province, Yuntai Mountains,Tanpu valley, river, periphyton, China (35°28′52′′N,113°39′21″E).Etymology: this species is named for its small but stout size.
4 DISCUSSION
This report represents the first detailed investigation of the freshwater diatom genusGomphonemaof Henan Province. While others have considered new taxa of the genus from a variety of places across the country, including the east (Zhang et al., 2016, 2018a,b), northeast (Skvortzow, 1929, 1935, 1946; Bao and Reimer, 1992; Fan et al., 2004; Liu et al., 2013),northwest (You et al., 2015), and southwest (Shi,2004; Li et al., 2006, 2010; Jiang et al., 2018), as well as Tibet (Mereschkowsky, 1906), few studies have focused on the central portion of China.
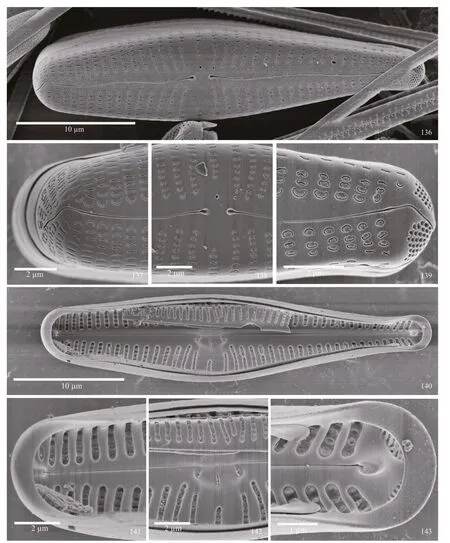
Plate 16 Gomphonema fortissimo sp. nov.
The Yuntai Mountains have a wide variety ofGomphonemamorphologies included in the region.Several morphological groups have been recognized for the genus (Kociolek and Stoermer, 1993; Reichardt and Lange-Bertalot, 1991; Reichardt, 1999; Kociolek and Kingston, 1999; Reichardt, 2015; Kociolek et al.,2016; Levkov et al., 2016; Jüttner et al., 2018) and,has been reported previously froom Hawaii (Kociolek et al., 2016) and Nepal (Jüttner et al., 2018). The Yuntai Mountains have a set of species that come from several of these groups. For example, in addition to the “classical” species of the genus, which have external flaps that give the external areolar openings a characteristic C- or S- shape (such asG.lanceolatoides,G.montanavivaandG.fortissimadescribed herein),as well as G.pseudosphaerophorumandG.vibrio,the latter taxon appears to lack areolar occlusions altogether (Jüttner et al., 2018). Also present are species that have internal areolae occlusions, such as
G. cf.lateripunctatumandG.pseudintricatumsp.nov. Cox (2004) termed these internal occlusions“tectula” and suggested they were supported by peglike projections. Further analysis is necessary to whether tectula and the peg-like struts that support them are homologous across the taxa that possess them.
TheGomphonemaflora of the Yuntai Mountains also includes an astigmate speciesG.yuntaiensisthat has unoccluded areolae, but also siliceous papillae positioned near the margin of the valve; this suite of morphological features has not been reported for the genus previously. Absent from this flora are species with striae composed of doubly rows of areolae (e.g.Hustedt, 1945; Reichardt, 2005; Kociolek et al.,2016), large, robust species (as seen in Lake Baikal but also SE Asia, Skvortzow and Meyer, 1928,Skvortzow, 1937; Skuja, 1937; Amossé, 1969;Kulivoskiy and Kociolek, 2014), and species with a wide axial area, often found in South America (Fricke,1902; see also Metzeltin and Lange-Bertalot, 1998)but also known from North America and Europe(Reichardt, 2007; Stancheva et al., 2016), Africa(Cholnoky, 1953) and Asia (Hustedt, 1937).
This morphological diversity ofGomphonemataxa represented in the Yuntai Mountains may be ofinterest as a measure of water quality status. The Yuntai Mountains occur in a Geological Park that has national and international significance. While the park is open to the public, there is an entrance fee and activities are highly regulated. There has been no monitoring of water quality of the GeoPark’s aquatic ecosystems, but a wide range of morphological diversity ofGomphonemaspecies has been reported from other pristine environments, including the Hawaiian Islands (Kociolek et al., 2016) and a highelevation lake in the Himalayas of Nepal (Jüttner et al., 2018). Future research may support the contention that high morphological diversity among forms currently assigned to the genusGomphonemais suggestive ofincreased water quality.
The diversity of forms assigned to the genusGomphonemamight suggest that additional subdivision of the genus is warranted. Such teasing apart of taxa formerly in the genusNaviculaBory andCymbellaC. A. Agardh, has led to new insights into the phylogeny of the former (Theriot et al., 2015;Kulikovskiy et al., 2019), and biogeographic trends in the latter (e.g. Kociolek, 2018). A finer-grained taxonomy of freshwater diatoms will be necessary across groups such as the genusGomphonema(with over 2 000 described taxa; Kociolek et al., 2020), as well asNitzschiaHassall,PinnulariaEhrenberg, andEunotiaEhrenberg, to name a few, to better understand the systematic relationships, evolution and biogeography of these successful lineages.
5 DATA AVAILABILITY STATEMENT
All data included in this study are available upon request by contact with the corresponding author.
6 ACKNOWLEDGMENT
We are grateful to Pan YU for his help in acquiring SEM images.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Steady increase in water clarity in Jiaozhou Bay in the Yellow Sea from 2000 to 2018: Observations from MODIS*
- Phylogenetic diversity and bioactivity of culturable deepsea-derived fungi from Okinawa Trough*
- Allelopathic eff ects of mixotrophic dinoflagellate Akashiwo sanguinea on co-occurring phytoplankton: the significance of nutritional ecology*
- Investigation of the decline of Ulva prolifera in the Subei Shoal and Qingdao based on physiological changes*
- Effi ciency of phosphorus accumulation by plankton,periphyton developed on submerged artificial substrata and metaphyton: in-situ observation in two shallow ponds*
- Petroleum exploitation enriches the sulfonamide resistance gene sul2 in off shore sediments