94份小麦育种亲本的功能基因分子检测
2021-06-09李玮宋国琦商航李玉莲张淑娟张荣志李吉虎高洁李根英
李玮,宋国琦,商航,李玉莲,张淑娟,张荣志,李吉虎,高洁,李根英
(1.山东省农业科学院作物研究所/农业部黄淮北部小麦生物学与遗传育种重点实验室/小麦玉米国家工程实验室,山东 济南 250100;2.新疆农业大学科学技术学院,新疆 乌鲁木齐 830052)
小麦是复杂的异源六倍体物种,其基因组约为17 Gb[1],是水稻的36倍多、玉米的7倍多。巨大的基因组给基因克隆和基因功能分析带来重重困难,是导致小麦分子育种滞后于水稻和玉米的原因之一。2012年Liu等[2]系统总结了小麦中已克隆的30多个功能基因或遗传位点及其相关的97个功能标记,主要涉及小麦加工品质、农艺性状和抗病性。功能标记是根据功能基因上的等位变异开发的分子标记,不同于连锁标记,不会因为染色体交换导致标记与表型不符,是用于分子标记辅助选择的理想标记。这97个功能标记主要为STS、SCAR和CAPS,需要通过琼脂糖或聚丙烯酰胺凝胶电泳进行检测,较为费时费力。2016年Rasheed等[3]报道了70个小麦KASP功能标记,相比需要通过凝胶电泳进行检测的分子标记,通过荧光检测的KASP标记检测速度提高了45倍。2017年国际小麦基因组测序联盟提前释放了中国春参考基因组序列,为小麦功能基因研究铺平了道路。2019年Khalid等[4]报道了小麦中已克隆功能基因或遗传位点增加到87个,对应的KASP标记数量也增加到124个。随着越来越多的KASP功能标记被开发,功能标记也逐渐被应用。2019年Zhao等[5]对来自全球的1 152份小麦材料进行了47个基因的KASP标记检测分析,粒重、抗旱、黄色素、穗发芽和株高相关的几个基因优异等位变异占比较高,而抗病相关的Lr68、Fhb1、Yr15和品质相关的Wx-B1优异等位变异占比较低,可作为未来改良的目标。2020年单子龙等[6]利用22个品质相关的KASP功能标记对153份河北省历年审定的小麦品种进行了检测,仅有5个基因的优异等位变异占比超过60%,7个基因的优异等位变异占比低于10%,表明通过辅助选择优异等位变异改良河北省小麦品质性状仍有力可为。2020年王志伟等利用抗病[7]、株高和粒重[8]相关的KASP标记对云南省42份小麦品种(系)进行了检测,绝大部分基因的优异等位变异占比较低,仍有遗传改良潜力。2021年Zhang等[9]对宁夏自治区种植的小麦地方品种、现代品种(系)和引进品种共207份进行了44个KASP标记检测,与粒重、硬度、黄色素、株高等相关的几个基因优异等位变异占比较高,其他大部分基因的优异等位变异占比有待提高。
现代育种技术已经从传统经验育种向精准育种转变,通过提高与育种目标相关功能基因优异等位变异的占比可以促进育种基础的提高,功能基因的分子标记辅助选择对小麦分子育种意义重大。山东省是我国小麦生产大省,为了促进小麦分子育种技术在山东的发展和应用,了解山东省小麦优异等位变异占比情况,本研究从课题组保存的小麦育种亲本出发,选择部分分型效果较好的KASP功能标记进行检测,旨在明确功能基因优异等位变异在育种亲本材料中的占比,促进山东省小麦育种单位利用小麦分子标记辅助选择进行遗传改良。
1 材料与方法
1.1 试验材料
本研究选用的小麦材料(表1)是本课题组收集保存用于育种的小麦品种或高代品系,共94份。2019年11月播种于山东省农业科学院作物研究所济南试验基地。播种机点播,行距33 cm,行长2 m,株距5 cm,按行长划排,排间留1 m的田间走道。
1.2 DNA提取
于小麦抽穗期取2 cm左右旗叶,液氮冷冻,用Geno/Grinder 2010组织研磨机(SPEX,美国)进行研磨,800 r/min研磨20 s。用快捷型植物基因组DNA提取系统[天根生化科技(北京)有限公司]提取DNA,去离子水溶解,调整浓度至100 ng/μL。
1.3 KASP标记检测
KASP反应体系和反应程序均参考Rasheed等[3]的,所用42个KASP标记引物序列来自Rasheed[3]、Khalid[4]和高洁[10]等,引物由青岛擎科生物技术有限公司合成。KASP Master Mix购自LGC公司(北京)。使用PHERAstar SNP分型检测仪(LGC,英国)检测KASP反应最终荧光颜色,检测结果导入Kluster Caller软件进行分型。基因名称和相应基因标记名称见表2。
表1 参试小麦品种(系)
表2 基因名称和相应基因标记名称
2 结果与分析
2.1 KASP标记检测效果
本研究共检测了42个KASP标记,基本上所有标记都能较好地分型(图1)。除TaGS2B1_1936检测缺失数据为15个外,其余标记的缺失数据均较少(图2),平均每个标记的缺失数据个数为2.4。基因的优异等位变异占比从0至100%,优异等位变异占比低于10%的有7个基因,高于80%的有11个。
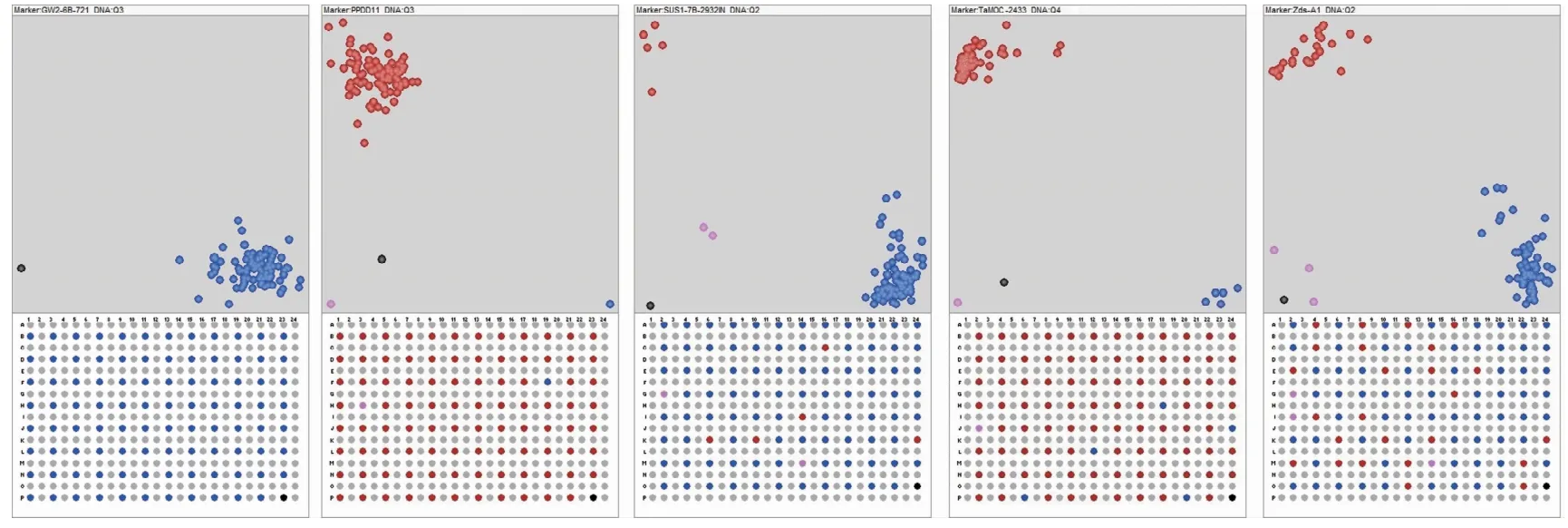
图1 部分KASP标记检测结果
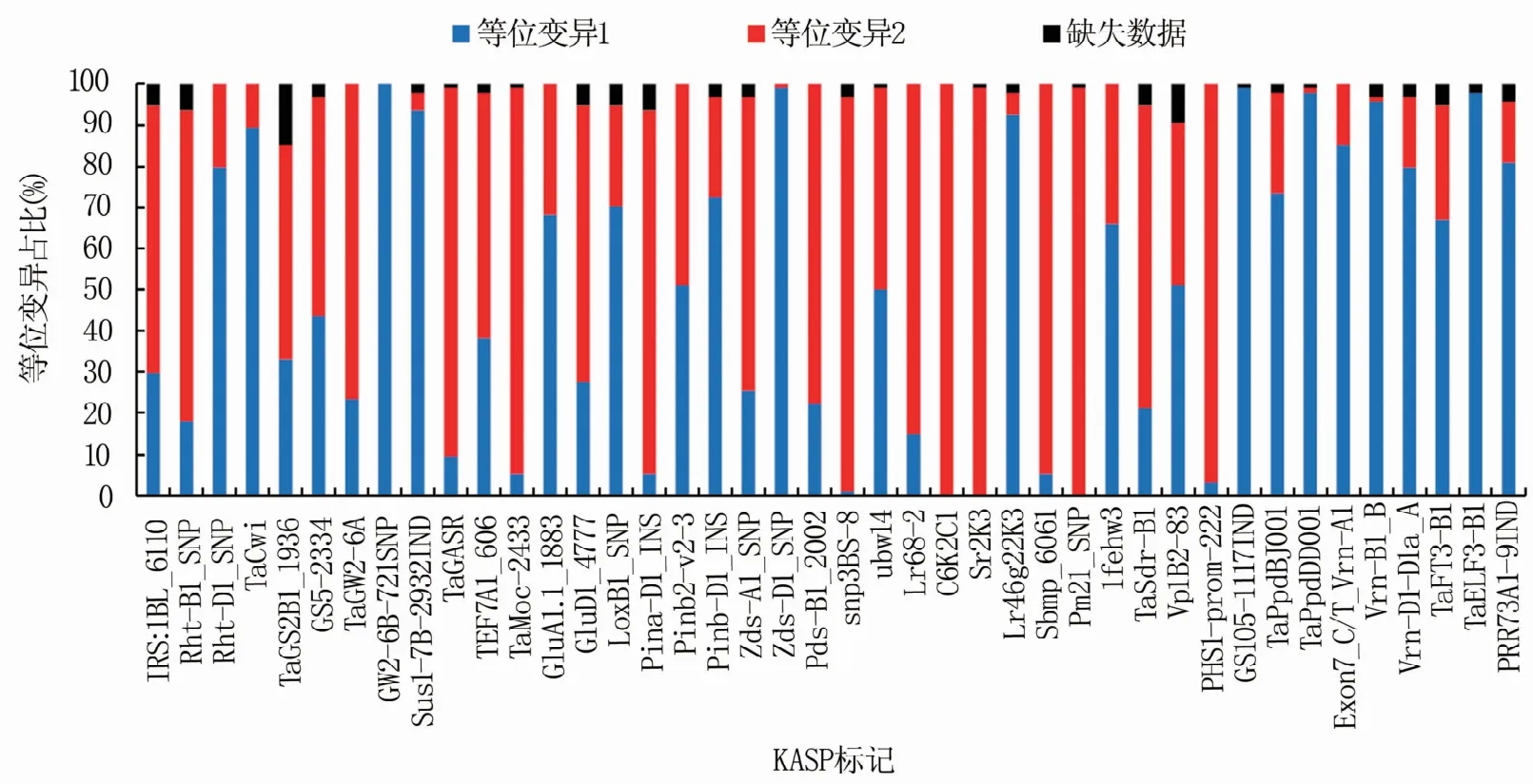
图2 94份小麦材料的等位变异占比
2.2 1B/1R易位检测结果
选择1RS∶1BL_6110检测1B/1R易位,94份参试小麦材料中28份含有1B/1R易位,占比30%。1RS染色体同时含有抗病基因和不利品质的黑麦碱基因,在本研究中将含有1B/1R易位列入优异等位变异。含有优异等位变异的材料有鲁原502、周麦22、周8425B、北京841、濮麦053、内乡188、HM-1、济宁12、西农511、周麦36、周麦38、JM268、德05-10、华麦1号、宁麦11、淮麦20、宁 洪17237、17J218、17J236、JN331、科 农3106、菏麦0662、菏麦0746-2、漯抗2号、K35、山农0713-2、山农多粒1号、山融3号。
2.3 株高相关标记检测结果
对Rht-B1和Rht-D1两个株高基因进行了KASP标记检测,分别有17份和75份材料含有矮秆等位变异,占比为18%和80%。同时含有两个矮秆基因的有3份材料,分别是豫麦47、周8425B和金麦1号。不含上述两个矮秆基因的材料是京411和北京841。
2.4 产量相关标记检测结果
检测了9个产量相关的KASP标记。千粒重相关的TaCwi、GW2-6B-721SNP和Sus1-7B-2932IND优异等位变异占比较高,分别是89%、100%和94%,而TaGS2B1_1936、GS5-2334、TaGW2-6A优异等位变异占比较低,分别是33%、44%和23%;TaGASR是粒长的KASP标记,优异等位变异占比为10%;TEF7A1_606是小穗粒数的KASP标记,优异等位变异占比38%;TaMoc-2433是穗数的KASP标记,优异等位变异占比仅为5%。参试小麦材料中,山融3号含有除TaMoc-2433外的8个优异等位变异,是含有产量相关优异等位变异最多的材料;其次是山农9540-55,含有除TaMoc-2433和TEF7A1_606的7个优异等位变异。同时含有TaMoc-2433和TEF7A1_606优异等位基因的材料有西农511和科农3106。
2.5 品质相关标记检测结果
检测了9个与品质相关的KASP标记。GluA1.1_1883和GluD1_4777是高分子量麦谷蛋白亚基2*/1和(5+10)标记,分别占68%和28%,参试小麦材料中济麦20、郑麦366、冀师02-1、洲元9369、藁优5766、烟农1212、烟农999、HM-1、硬B2 16-6、济宁12、济麦229、济麦44、中麦578、新麦9、华麦1号、宁麦11、漯抗2号、山农0538、山农24、山农25同时含有优异亚基2*/1和(5+10);脂肪氧化酶标记LoxB1_SNP的低酶活优异等位基因占比70%,含有脂肪氧化酶优异等位变异的材料较多;与硬度相关的Pina-D1_INS、Pinb2-v2-3和Pinb-D1_INS的优异等位变异占比分别为5%、51%和72%,一般小麦中硬度优异等位变异仅含Pina-D1和Pinb-D1其中一个,本研究中含有Pina-D1优异等位变异的材料较少,冀师02-1、Manital和藁优5766同时含有Pina-D1和Pinb2-V优异等位变异;与黄色素相关的Zds-A1_SNP、ZDS-D1_SNP和Pds-B1_2002的优异等位变异占比分别为26%、99%和22%,同时含有三个黄色素优异等位变异的材料有新麦26、内乡188、济宁12和山农29。
2.6 抗性相关标记检测结果
检测了12个与抗性相关的KASP标记。抗赤霉病的Fhb1的snp3BS-8检测阳性材料仅有山农抗赤1号,占比1%;抗小麦黄花叶病的Sbmp_6061检测阳性占比5%,有烟农19、硬B2 16-6、济糯麦1号、漯抗1号和山农25;未检测到Pm21阳性材料;抗叶锈的ubw14和Lr68-2检测阳性占比分别是50%和15%,兼抗条锈、叶锈和白粉的多效抗病基因标记C6K2C1、Sr2K3和Lr46g22K3检测阳性占比分别是0、0和93%,ubw14、Lr68-2和Lr46g22K3均为阳性的材料有京411、北京841、陕7859、烟农1212、烟农999、济宁12、济麦60、西农511、德05-10、华麦1号、淮麦20、17J218、山融3号;抗旱的1fehw3检测阳性材料占比66%,含有该基因的材料较多;抗穗发芽的TaSdr-B1和PHS1-prom-222优异等位变异占比分别为21%和51%,Vp1B1有a、b、c三个等位变异,等位变异a不抗穗发芽,等位变异b和c抗穗发芽,Vp1B2-83不能区分a和b,故本研究中将c作为优异等位变异,占比51%,同时含有Vp1B2-83和PHS1-prom-222优异等位变异的材料有洲元9369和华麦1号,同时含有TaSdr-B1和Vp1B2-83优异等位变异的材料有陕7859、藁优5766、内乡188、中麦578、宁麦11、科农3106、漯抗1号、漯抗2号、K35、山农0538、山农抗赤1号、山农特大粒1号、山融3号。
2.7 春化、光周期相关标记检测结果
检测了6个与春化、光周期相关的KASP标记。光周期标记GS105-1117IND、TaPpdBJ001和TaPpdDD001优异等位变异占比分别为99%、73%和98%;春化基因标记Exon7_C/T_Vrn-A1、Vrn-B1_B、Vrn-D1-D1a_A优异等位变异占比分别为85%、96%和80%。光周期和春化基因决定了小麦的生态适应性,因此相同生态区的小麦主效基因的等位变异基本上是一致的且占比很高。
2.8 开花期相关标记的检测结果
检测了3个与开花期相关的KASP标记。开花期的优异等位变异与生态区有关,本研究中将检测数量较多的等位变异作为优异等位变异。TaFT3-B1、TaELF3-B1和PRR73A1-9IND优异等位变异的占比分别为67%、98%和81%。
3 讨论与结论
1B/1R易位系小麦品种具有适应性好、耐后期高温、高产稳产等优点[11],这主要得益于1RS染色体上携带有Lr26、Sr31、Yr9和Pm8等抗病基因和其他高产稳产基因或基因组合[12]。1B/1R易位系被作为优异亲本广泛利用,是外源基因用于小麦品种改良最成功的范例[13],选择是否携带1RS染色体最简便有效的方法就是分子标记辅助选择。近期,Qi等[14]提出了作物育种“布达拉”模型,首先将品种的优良性状和优异基因定向聚集,这就需要通过功能标记辅助选择来实现。小麦中已经克隆的功能基因相对较少,如何利用功能标记辅助选择将这些已克隆的功能基因用于育种是当务之急。虽然国内有关亲本基因型检测的报道不少,但利用功能标记辅助选择成功选育小麦品种还鲜有报道。加强对现有功能标记的利用,特别是育种亲本中占比较低的优异等位变异的利用对小麦改良有重要意义。本研究中与产量相关 的KASP标 记TaGASR、TaMoc-2433、TaGS2B1_1936、GS5-2334、TaGW2-6A,与品质相关的KASP标记GluD1_4777、Zds-A1_SNP、Pds-B1_2002,与抗性相关的KASP标记snp3BS-8、Sbmp_6061、Lr68-2、C6K2C1、Sr2K3、TaSdr-B1在检测的94份材料中优异等位变异占比均较低,可作为未来小麦分子育种应用的重要方向。
SNP是基因组中最丰富的变异类型,是未来分子标记利用的方向,随着越来越多功能基因被克隆,KASP功能标记数量也会逐步增加。KASP是检测SNP最简便灵活的技术,已被广泛应用。除LGC公司专用的检测仪器外,荧光定量PCR仪、荧光酶标仪等均可用于检测KASP标记。2019年KASP功能标记数量已达124个[4],Zhao[5]、Zhang[9]等 分 别 选 择 了47个 和44个KASP标记,本研究选用了42个标记,这是因为小麦基因组的复杂性导致将SNP转化成KASP标记相对比较困难[15]。已开发的KASP标记可能还存在假阳性等问题[3],部分KASP标记的检测结果不尽如人意,随着KASP标记开发和应用的成熟,这些问题都有望得到解决[10,15]。
