γ-氨基丁酸对热应激雏鸡胰腺组织结构、抗氧化能力、消化酶活性及细胞凋亡的影响
2021-06-05余哲琪田佳迎
温 静 余哲琪 田佳迎 陈 忠
(海南师范大学热带岛屿生态学教育部重点实验室,海口 571158)
近年来,由于畜牧业生产的集约化发展,热应激对畜禽造成的影响越来越严重[1]。热应激影响着家禽生产力的诸多方面,如导致肉禽生产性能下降[2]、繁殖能力降低[3],同时阻碍家禽的法氏囊等免疫器官的发育,降低抗氧化能力,提高细胞凋亡率[4];热应激还会引起雏鸡小肠黏膜的抗氧化功能明显损伤[5],并且显著降低消化酶的活性,从而导致肠道吸收功能和免疫屏障严重受损[6];同时,热应激也影响家禽肠道中蛋白质的利用率以及营养转运体的基因表达,甚至增加家禽的死亡率[7-8]。因此,高温环境导致的热应激问题使家禽业的发展面临着巨大的挑战[9]。胰腺是家禽重要的代谢器官和消化器官,在胰液的分泌、消化酶的合成以及营养物质的代谢等方面具有十分重要的作用。消化酶活性是反映机体消化生理机能的一项重要指标,消化酶活性的高低反映了动物机体对营养物质的消化能力,直接影响蛋白质、脂肪等营养物质的消化吸收,最终影响动物的生产性能[10-11]。
γ-氨基丁酸(GABA)是一种广泛存在于动物体内的抑制性神经递质,国家卫生部2009年批准其用于食品加工。现在GABA作为一种安全的食品添加剂,广泛应用于食品和畜牧业,能够有效减轻热应激对家畜生理功能的影响,被认为是一种安全的饲料添加剂[12-13]。本课题组前期研究已经证实,每天灌喂50 mg/kg GABA可以减缓热应激[(40.0±0.5) ℃]对雏鸡肠道造成的损伤,有效改善热应激雏鸡小肠黏膜的免疫功能[14];另外,将30 mg/kg GABA添加到仔猪饲粮中能够抑制肠上皮细胞凋亡,保持肠绒毛的完整性,降低仔猪的腹泻率[15]。有研究指出,在热应激[(30±2) ℃]蛋鹌鹑的饲粮中添加25 mg/kg GABA可提高蛋鹌鹑的生产性能以及抗氧化能力[16];而在饲粮中添加5 g/kg谷氨酰胺和100 mg/kg GABA可以提高肉鸡在高温(30~34 ℃)环境下的抗热应激能力以及血清胰高血糖素、谷氨酸、胰岛素含量和肌酸激酶、碱性磷酸酶活性等血清参数[17]。目前关于热应激对雏鸡消化系统影响的研究主要集中在肠道方面,而对胰腺的研究报道较少。因此,研究热应激对雏鸡胰腺组织结构、生理生化等方面的损伤以及缓解这些损伤的措施至关重要。因此,本研究拟探讨GABA对热应激雏鸡胰腺组织结构、抗氧化能力、消化酶活性及细胞凋亡情况的影响,明确GABA能否恢复热应激对胰腺组织结构、消化酶、抗氧化酶造成的损伤,为GABA在家禽抗热应激领域的进一步应用提供依据。
1 材料与方法
1.1 试验设计
将240只1日龄健康雄性文昌鸡雏鸡随机分为对照组(CK组)、热应激组(HS组)、对照+GABA组(CK+组),热应激+GABA组(HS+G组),每组60只(每组6个重复,每个重复10只),称重和编号,使雏鸡组间体重、采食量无显著差异(P>0.05)。所有雏鸡均自由采食同一饲粮(市售肉小鸡配合饲料,营养水平见表1)和饮用蒸馏水。各组雏鸡均在常规条件下饲养6周,每天记录直肠温度、采食和饮水情况。从2周龄开始,CK组与HS组每日每只灌喂0.2 mL的生理盐水,CK+G组和HS+G组每日每只灌喂0.2 mL的GABA水溶液(GABA灌喂量为50 mg/kg BW)。每天灌喂2 h后于12:00—14:00对HS组和HS+G组文昌鸡雏鸡进行热应激,气候箱设定温度在(40.0±0.5)℃、相对湿度为(70±5)%,同时将CK组和CK+G组的文昌鸡雏鸡放于温度(32.46±0.16) ℃、相对湿度(70±5)%的气候箱中,热应激结束后,再将全部雏鸡放回鸡笼常规饲养。

表1 饲粮营养水平(风干基础)
1.2 样品采集和处理
于2~6周龄末,每组分别取6只文昌鸡雏鸡,剖取胰腺,去除脂肪称其鲜重。一部分胰腺组织保存于-80 ℃,以供进一步分析;另一部分胰腺组织用Bouin固定液固定,用于制作组织切片。
1.3 胰腺组织结构观察
将胰腺组织切片常规苏木精-伊红(HE)染色,随机选取3张,用Olympus-BX50F显微镜观察胰腺组织结构,再以YD400C数字摄像系统进行拍摄,每张切片拍摄5个不同视野。
1.4 胰腺组织中抗氧化指标测定
用低温磷酸盐缓冲液(PBS)清洗胰腺组织以除去血液,并用滤纸吸干。以质量体积比9∶1的比例添加pH 7.4的PBS,在4 ℃下用研磨仪研磨3 min,直到细胞完全破碎,液体颜色均匀。随后,将胰腺组织匀浆在4 ℃下3 000 r/min离心10 min,收集上清液并在-20 ℃下储存,用以测定谷胱甘肽过氧化物酶(GSH-Px)、超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量,操作严格按照试剂盒说明书进行。
1.5 胰腺组织中GSH-Px、SOD mRNA相对表达量的测定
1.5.1 胰腺组织总RNA的提取及浓度检测
用电动研磨器对胰腺组织进行研磨匀浆,按照总RNA提取试剂盒(货号:dp431,天根生化科技有限公司)提取胰腺组织总RNA,通过超微量紫外分光光度计测量胰腺组织总RNA的浓度和纯度。根据OD260 nm/OD280 nm来判断RNA纯度,二者比值在1.8~2.0为宜。
1.5.2 cDNA的合成
采用Fast Quantc DNA第1链合成试剂盒(货号:kr116,天根生化科技有限公司),于冰上进行cDNA的反转录。先配制gDNA去除反应体系:取5×gDNA Buffer 2 μL、总RNA 1 μg,用RNase-Free ddH2O补足至10 μL,彻底混匀,短暂离心后置于42 ℃孵育3 min,然后置于冰上;再配制反转录反应体系:10×Fast RT Buffer 2 μL、RT Enzyme Mix 1 μL、FQ-RT Primer Mix 2 μL,用RNase-Free ddH2O补足至10 μL,然后与第1步的gDNA去除反应体系(10 μL)混匀,42 ℃孵育15 min后,再于95 ℃孵育3 min,得到的cDNA于-20 ℃低温保存用于后续试验。
1.5.3 引物设计与实时荧光定量PCR条件
PCR引物运用Primer软件设计,由生工生物工程(上海)股份有限公司合成,引物序列及参数见表1。将GSH-Px、SOD引物浓度稀释至10 μmol/L。以内参基因β-肌动蛋白(β-actin)为内标,对GSH-Px、SODmRNA进行相对定量分析。
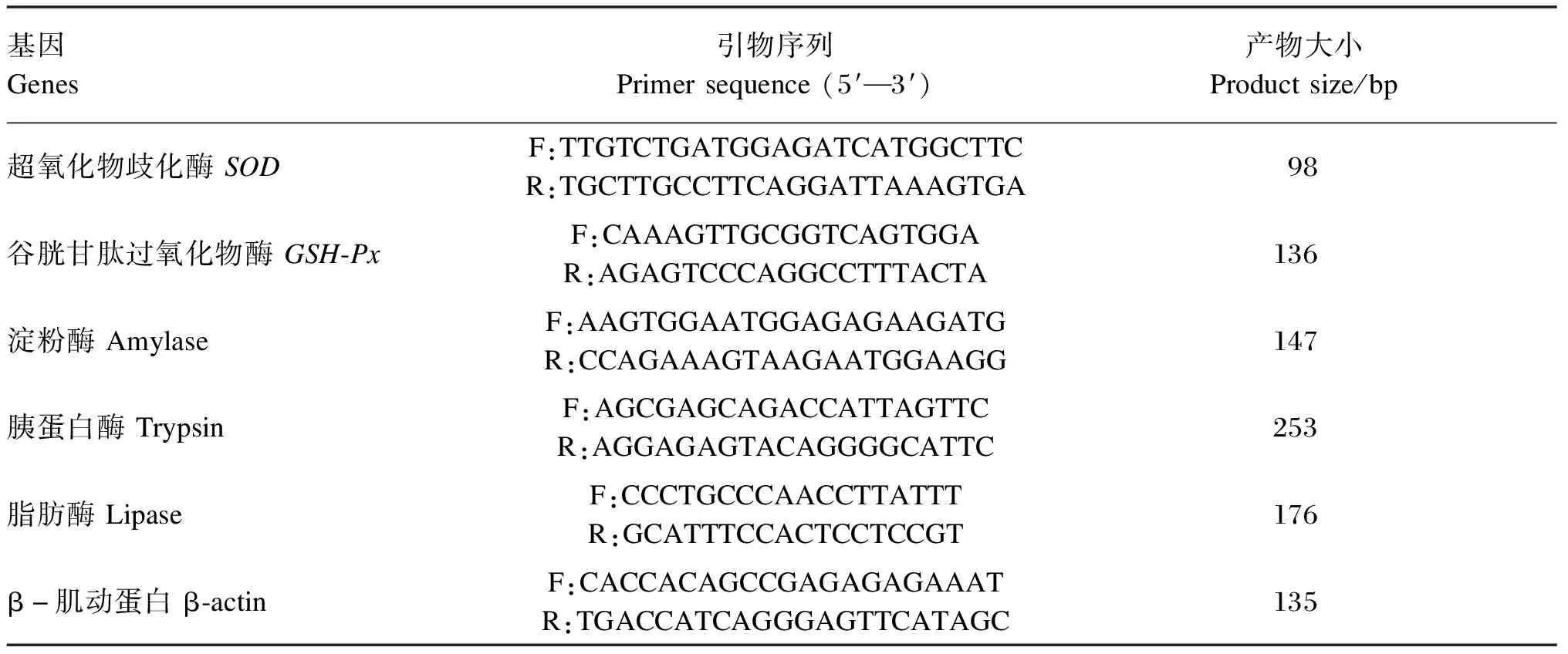
表2 引物信息
1.6 胰腺组织中消化酶活性的测定
胰腺组织上清液的提取方法同1.4。采用南京建成生物工程研究所生产的淀粉酶试剂盒(货号:C016-2-1)、脂肪酶试剂盒(货号:A054-1-1)和胰蛋白酶试剂盒(货号:A080-2-1)测定相应的酶活性,并严格按照说明书进行操作。
1.7 胰腺组织中淀粉酶、脂肪酶和胰蛋白酶mRNA相对表达量的测定
胰腺组织总RNA的提取、浓度检测及cDNA的合成同1.5。PCR引物运用Primer软件设计,引物序列及参数见表1。将淀粉酶、脂肪酶、胰蛋白酶引物浓度分别稀释至10 μmol/L,以内参基因β-actin为内标,对淀粉酶、脂肪酶、胰蛋白酶mRNA进行相对定量分析。
1.8 胰腺细胞凋亡及分布密度
采用荧光法TUNEL细胞凋亡检测试剂盒(货号:G002-2-2,南京建成生物工程研究所)对常规石蜡切片进行处理,每个样本选取3张切片,每张切片取5个视野,经YD400C数字摄像系统对组织切片进行拍照处理,运用Image-Plus Pro软件测量凋亡细胞数量及视野面积,通过(凋亡细胞数量/视野面积)得出凋亡细胞分布密度。
1.9 数据处理与统计分析
试验数据用平均值±标准误(mean±SE)表示,应用SPSS 21.0软件对数据进行单因素方差分析,并采用Duncan氏法进行多重比较,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果与分析
2.1 胰腺组织结构观察
观察雏鸡胰腺组织结构(图1)发现,与CK组比较,HS组雏鸡胰腺组织腺泡破坏,纹理紊乱,有的细胞核固缩,溶解消失,呈现一定的空泡化;而CK+G组雏鸡胰腺组织腺泡结构完整,轮廓清晰,无明显异常;HS+G组雏鸡胰腺组织腺泡结构完整,轮廓较清晰,部分纹理紊乱,胰腺组织结构较正常。

A:CK组;B:CK+G组;C:HS组;D:HS+G组。比例尺为50 μm。5W:5周龄。箭头所指为发生病理变化。
2.2 GABA对热应激雏鸡胰腺组织抗氧化能力的影响
如表3所示,在2~6周龄时,4组雏鸡胰腺组织SOD活性均随着周龄的增长呈现先上升后下降的趋势,HS组雏鸡的SOD活性一直为4组中最低水平。在5、6周龄时,CK+G组和HS+G组的SOD活性高于CK组和HS组(P>0.05),而在2、4周龄时,CK组的SOD活性显著高于HS组(P<0.05)。在2~6周龄时,HS组雏鸡胰腺组织GSH-Px活性随着周龄的增长一直为下降趋势,CK组和CK+G组则总体呈现上升趋势,在6周龄时达到峰值且CK组GSH-Px活性显著高于HS组(P<0.05)。HS+G组雏鸡胰腺组织GSH-Px活性随着周龄的增长呈现先下降后上升的趋势,而且在5周龄时显著高于HS组(P<0.05)。通过对4组雏鸡胰腺组织MDA含量进行比较发现,在6周龄时CK+G组和HS+G组MDA含量显著低于HS组(P<0.05)。组别对雏鸡胰腺组织SOD和GSH-Px活性有显著影响(P<0.05),且周龄与组别间的交互作用对雏鸡胰腺组织MDA含量有显著影响(P<0.05)。
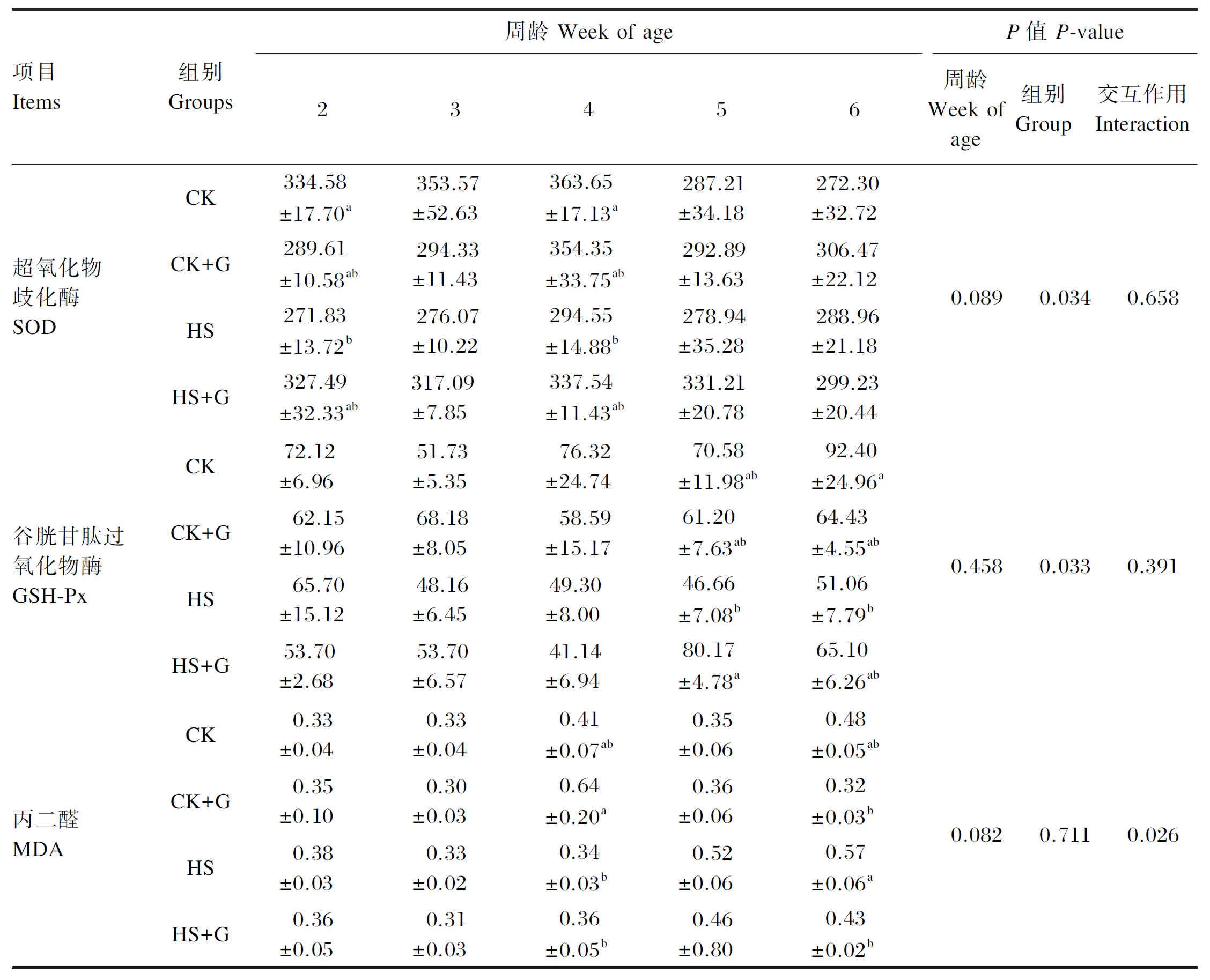
表3 GABA对热应激雏鸡胰腺组织抗氧化能力的影响
2.3 GABA对热应激雏鸡胰腺组织中SOD、GSH-Px mRNA相对表达量的影响
如表4所示,在2~4周龄时,4组雏鸡胰腺组织中SOD mRNA相对表达量无显著差异(P>0.05),且均随着周龄的增长总体呈现先上升后下降的趋势。HS组胰腺组织中SODmRNA相对表达量在5周龄时出现峰值,显著高于CK组和HS+G组(P<0.05)。CK+G组胰腺组织中SODmRNA相对表达量在6周龄时达到峰值,显著高于HS组(P<0.05)。在2~6周龄时,随着周龄的增长,CK组和HS+G组胰腺组织中GSH-PxmRNA相对表达量呈现上升趋势,而CK+G组则呈现先上升后下降的趋势,且在5周龄时达到峰值,显著高于HS组(P<0.05)。在2周龄时,HS组胰腺组织中GSH-PxmRNA相对表达量显著高于CK组(P<0.05),但是在6周龄时,CK组胰腺组织中GSH-PxmRNA相对表达量显著高于HS组(P<0.05)。组别仅对雏鸡胰腺组织中SODmRNA相对表达量有显著影响(P<0.05),而周龄对雏鸡胰腺组织中SODmRNA相对表达量有极显著影响(P<0.01),对GSH-PxmRNA相对表达量有显著影响(P<0.05)。
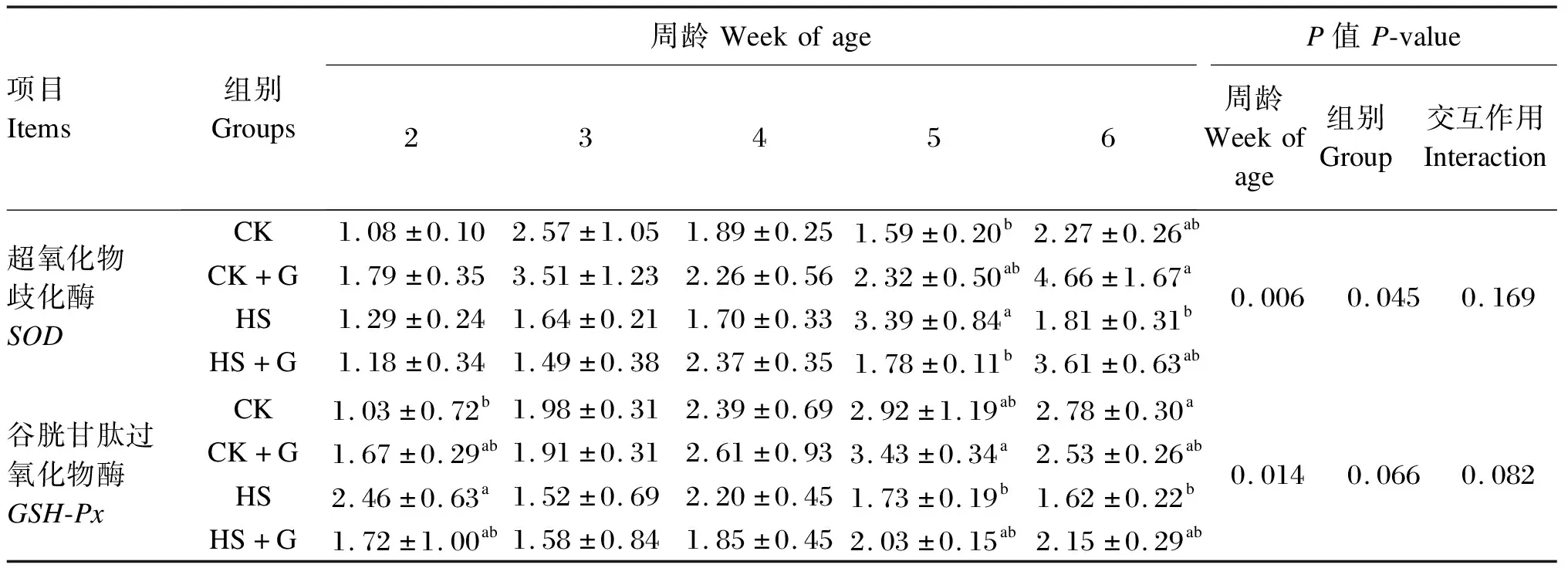
表4 GABA对热应激雏鸡胰腺组织中SOD、GSH-Px mRNA相对表达量的影响
2.4 GABA对热应激雏鸡胰腺组织中消化酶活性的影响
如表5所示,在2~6周龄时,各组雏鸡胰腺组织中淀粉酶活性随着周龄的增长都呈现先上升后下降的趋势,CK组、CK+G组和HS+G组均在5周龄时出现峰值,而HS组在3周龄时出现峰值,且在5周龄时CK组和CK+G组与HS组具有显著差异(P<0.05)。在6周龄时,CK+G组胰腺组织中淀粉酶活性显著高于HS组(P<0.05)。在2~6周龄时,随着周龄的增长,CK组和HS组胰腺组织中脂肪酶活性呈现先上升后下降的趋势,而CK+G组和HS+G组则呈现上升趋势。在3周龄时,HS+G组胰腺组织中脂肪酶活性显著高于CK组(P<0.05)。在5周龄时,CK+G组胰腺组织中脂肪酶活性显著高于HS组和HS+G组(P<0.05),在6周龄时,胰腺组织中脂肪酶活性在CK组、HS组和CK+G组间均具有显著差异(P<0.05),CK+G组显著高于CK组和HS组(P<0.05)。在2、3周龄时,4组雏鸡胰腺组织中胰蛋白酶活性均较低,但随着周龄的增长呈现上升趋势。在4周龄时,CK+G组和HS+G组胰腺组织中胰蛋白酶活性急剧增加,且CK+G组显著高于HS组(P<0.05)。周龄、组别对雏鸡胰腺组织中淀粉酶、脂肪酶和胰蛋白酶活性均有极显著影响(P<0.01),而两者间的交互作用仅对淀粉酶活性有显著影响(P<0.05)。

表5 GABA对热应激雏鸡胰腺组织消化酶活性的影响
2.5 GABA对热应激雏鸡胰腺组织中淀粉酶、脂肪酶和胰蛋白酶mRNA相对表达量的影响
如表6所示,在2~6周龄时,CK+G组胰腺组织中淀粉酶mRNA相对表达量均高于其他3组,但是在2、3周龄时各组之间没有显著差异(P>0.05),在4周龄时CK+G组显著高于其他3组(P<0.05),在5、6周龄时CK+G组有下降趋势,但仍显著高于其他3组(P<0.05)。在2~6周龄时,CK组和HS+G组胰腺组织中脂肪酶mRNA相对表达量随着周龄的增长均呈现上升趋势,且HS组雏鸡胰腺组织中的脂肪酶mRNA相对表达量普遍低于其他3组,仅在3周龄时接近CK组。在2周龄时,CK+G组胰腺组织中脂肪酶mRNA相对表达量显著高于CK组和HS组(P<0.05),在5、6周龄时,相对于CK组,CK+G组胰腺组织中脂肪酶mRNA相对表达量达到较高水平,且均显著高于HS组(P<0.05),但在3周龄时,HS+G组胰腺组织中脂肪酶mRNA相对表达量显著高于CK组(P<0.05)。在2~6周龄时,CK组、CK+G组和HS+G组雏鸡胰腺组织中胰蛋白酶mRNA相对表达量随着周龄的增长均呈现先上升后下降的趋势,而HS组则呈现上升趋势。在2、4、5周龄时,CK+G组胰腺组织中胰蛋白酶mRNA相对表达量具有较高水平,均显著高于HS组(P<0.05),而且在4周龄时,CK+G组胰腺组织中胰蛋白酶mRNA相对表达量显著高于HS组和HS+G组(P<0.05)。在3周龄时,HS+G组胰腺组织中胰蛋白酶mRNA相对表达量显著高于HS组(P<0.05)。组别对雏鸡胰腺组织中淀粉酶、胰蛋白酶和胰蛋白酶mRNA相对表达量均有极显著影响(P<0.01),周龄对淀粉酶和胰蛋白酶mRNA相对表达量有极显著影响(P<0.01),对脂肪酶mRNA相对表达量有显著影响(P<0.05),而两者间的交互作用仅对淀粉酶mRNA相对表达量有显著影响(P<0.05)。

表6 GABA对热应激雏鸡胰腺组织中淀粉酶、脂肪酶和胰蛋白酶mRNA相对表达量的影响
2.6 GABA对热应激雏鸡胰腺组织细胞凋亡的影响
凋亡细胞在荧光显微镜下呈现绿色荧光,图2显示,5周龄CK组雏鸡胰腺组织中凋亡细胞数量明显少于HS组和HS+G组,与HS组比较,HS+G组雏鸡胰腺组织凋亡细胞数量低于HS组,而CK+G组雏鸡胰腺组织凋亡细胞数量低于CK组。
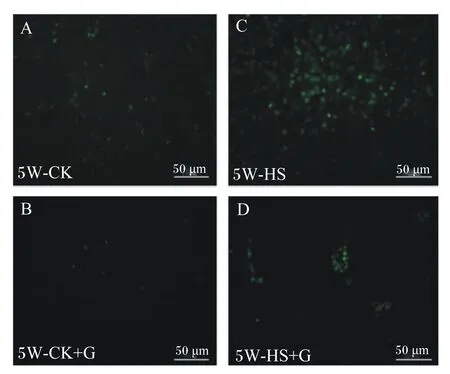
A:CK组;B:CK+G组;C:HS组;D:HS+G组。比例尺为50 μm。5W:5周龄。
如表7可知,在2~6周龄时,CK+G组和HS+G组的凋亡细胞分布密度随着周龄的增长均呈现出先上升后下降的趋势,在3周龄时达到峰值,而HS组则呈现上升趋势。在5、6周龄时,HS组凋亡细胞分布密度显著高于其他3组(P<0.05);在4、6周龄时,HS+G组凋亡细胞分布密度下降,且与CK组差异不显著(P>0.05)。组别对雏鸡胰腺组织中凋亡细胞分布密度有极显著影响(P<0.01),且组别与周龄间的交互作用对其有显著影响(P<0.05)。
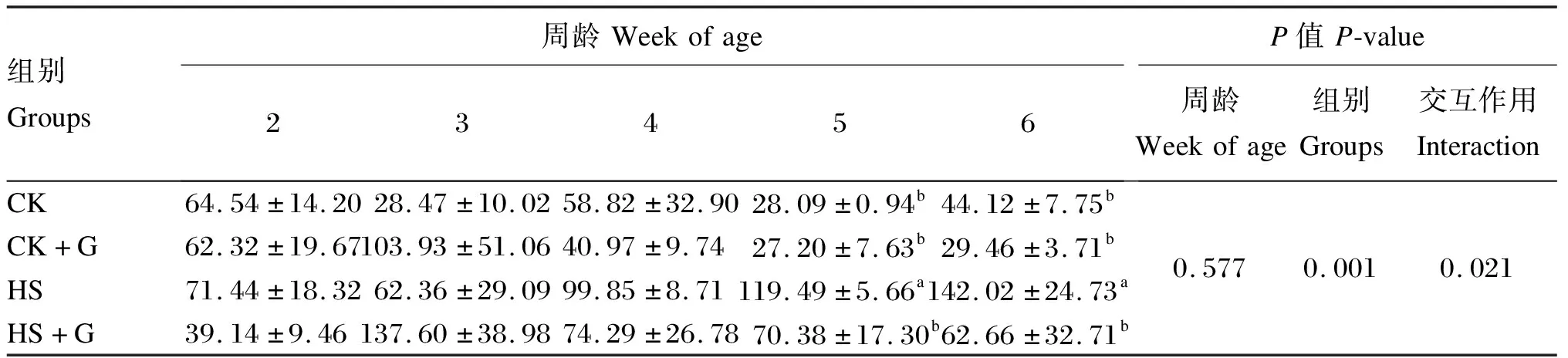
表7 GABA对热应激雏鸡胰腺组织凋亡细胞分布密度的影响
3 讨 论
有研究表明,与正常对照组小鼠相比,热应激小鼠的胰腺重量和系数略有降低,表明热应激可能损伤了小鼠的胰腺组织结构与功能[18]。在生长初期,雏鸡需要较高的温度来维持其生长发育[19],因此,早期热应激对雏鸡胰腺组织的影响并不大。然而在热应激后期,雏鸡胰腺组织出现结构紊乱,组织损伤严重。GABA处理则使热应激雏鸡胰腺组织结构的完整性接近CK组,表明GABA能够减轻热应激对雏鸡胰腺组织结构的损伤,维持胰腺组织结构发育接近正常水平。
本试验还发现,4组雏鸡胰腺组织中SOD活性仅在2、6周龄时具有显著差异,在6周龄时HS组、HS+G组和CK+G组均高于CK组,这可能是由于HS组和HS+G组的雏鸡胰腺受到长期热攻击,使得胰腺组织细胞内活性氧(ROS)过量产生,而为了维持胰腺组织内的氧化-抗氧化平衡,机体通过代偿机制上调SOD活性维持氧化-抗氧化平衡,而CK+G组可能是由于GABA的长期添加,GABA促使SOD基因表达上调,从而增加雏鸡胰腺组织的抗氧化能力。雏鸡胰腺组织中SODmRNA相对表达量在5、6周龄时具有显著差异,在5周龄时,HS组胰腺组织中SODmRNA相对表达量显著高于CK组,HS+G组胰腺组织中SODmRNA相对表达量高于CK组,也是机体通过代偿机制增强胰腺组织的抗氧化能力。在6周龄时,由于GABA的长期添加,CK+G组胰腺组织中SODmRNA相对表达量较HS组显著上调,增强了胰腺组织的抗氧化能力。Zhang等[20]的试验结果显示,饲粮中添加50 mg/kg GABA能提高罗马母鸡血清胰岛素水平,免疫球蛋白G(IgG)、免疫球蛋白A(IgA)和补体3(C3)含量,使抗氧化能力和免疫功能得到增强,从而提高母鸡的生产性能。在2~6周龄期间,仅在5周龄时HS+G组雏鸡胰腺组织中GSH-Px活性高于其他3组。胰腺组织中的抗氧化系统处于动态平衡状态,抗氧化系统中SOD和GSH-Px的活性不能同时升高,一种机制的激活和另一种机制的相应抑制可能就是体内的自我保护机制。然而,这种可能的机制仍需进一步研究和证实。而在5周龄时CK+G组胰腺组织中GSH-PxmRNA相对表达量显著高于HS组,可能是由于GABA代谢产生了大量谷氨酸[21]。谷氨酸是合成GSH-Px的关键元素,能够维持体内GSH-Px活性的保持在一定水平。另外,在5周龄时,HS组和HS+G组胰腺组织中MDA含量高于CK组,这表明由于胰腺组织受到累积热应激的攻击,使得胰腺细胞产生过多的MDA,而此时GABA发挥清除胰腺组织中MDA的能力不足。在6周龄时,CK+G组和HS+G组胰腺组织中MDA含量低于HS组和CK组,证明GABA在6周龄起到了清除胰腺组织中MDA的作用,这可能是由于投喂的GABA是间接作用于胰腺组织,使其SODmRNA相对表达量在6周龄处于较高水平,因此,GABA能够显著提高机体的抗氧化能力。
Wang等[22]研究发现,在饲粮中添加50 mg/kg GABA可以明显提高奶牛的采食量,并可提高奶牛的泌乳量,且具有剂量效应。有研究报道,35~36 ℃高温条件下在饲粮中添加50~100 mg/kg GABA有助于鸡对高温环境的适应,提高产蛋性能[23-24]。蒋磊等[25]的研究表明,32 ℃高温处理显著降低了肉鸡十二指肠中胰蛋白酶、脂肪酶及淀粉酶活性。阮晖等[26]报道,在34.7 ℃热应激条件下,肉鸡小肠内总蛋白水解酶、脂肪酶和淀粉酶活性均显著下降,且肉鸡日增重与这几种酶的活性呈极显著正相关,并通过配对试验证明上述酶活性的降低并非由采食量下降所致,而是高温环境直接作用的结果。在目前的研究中,36 ℃高温处理后肉鸡空肠脂肪酶和胰蛋白酶活性显著降低[27]。本研究则发现,在3~6周龄,HS组雏鸡胰腺组织中3种消化酶活性都低于其他3组,这一结果可能是因为热应激导致交感神经兴奋,引起血管收缩,减少胰腺的血流量,使胰液的分泌减少,从而各个消化酶的分泌量减少;而在6周龄时,CK+G组和HS+G组雏鸡胰腺组织中淀粉酶和胰蛋白酶活性高于CK组,说明GABA的添加可以增强胰腺组织中消化酶的活性。另外,在3周龄时,HS组雏鸡胰腺组织中淀粉酶和蛋白酶mRNA相对表达量高于CK组,这是由于雏鸡处于快速生长期,组织通过代偿功能来缓解热应激造成的损伤,从而刺激胰腺组织中淀粉酶和蛋白酶mRNA表达量增加,而且在长时间的热应激条件下雏鸡会通过增强新陈代谢来调节体温[28]。在2、4、5、6周龄时,CK+G组雏鸡胰腺组织中淀粉酶和脂肪酶活性都高于其他3组,HS+G组雏鸡胰腺组织中淀粉酶和脂肪酶活性都高于HS组和CK组,表明热应激会降低雏鸡胰腺组织中主要消化酶基因的表达,与消化酶活性测定结果基本一致。而在2~5周龄时,HS组雏鸡腺胰组织中蛋白酶mRNA相对表达量显著低于HS+G组,说明GABA的添加促进了热应激雏鸡胰腺组织中胰蛋白酶基因的表达。由于消化酶合成与分泌的机理目前还没有十分清楚,关于热应激对动物胰腺组织中主要消化酶基因表达的影响又少见报道,所以热应激使消化酶基因的表达量下降的机理还有待进一步的研究。
细胞凋亡作为生物体内的一个重要生命现象,对机体的正常发育具有重要作用,一旦细胞凋亡发生异常,机体发生疾病的概率就会增加[29]。在2~6周龄时,CK组雏鸡胰腺组织中细胞凋亡情况变化不大,随周龄的增长呈现出先上升后下降的过程,在4周龄时凋亡细胞分布密度达到最大,而在5周龄时细胞凋亡数量又下降到最低水平。这可能是因为4周龄为雏鸡发育最旺盛的时期,自然凋亡细胞数量增多所致,而在5周龄时凋亡细胞数量下降则可能与该时期雏鸡胰腺发育缓慢相关。在2周龄时,HS组雏鸡胰腺组织中细胞凋亡处于较低水平,而在3~6周龄期间,胰腺组织中细胞凋亡水平急剧上升,这可能是由于热应激对雏鸡胰腺组织造成的损伤还不足以使得胰腺组织中细胞的正常凋亡过于紊乱,而从3周龄开始,由于雏鸡快速生长,以及胰腺组织所受的热应激的损伤累积,使得雏鸡胰腺组织中细胞凋亡水平急剧上升。有研究发现慢性热应激导致肉鸡小肠中消化酶活性失调,细胞增殖被抑制,细胞凋亡增加[28]。HS+G组雏鸡胰腺组织中细胞凋亡水平在3周龄时较高,但在3周龄后HS+G组雏鸡胰腺组织中细胞凋亡水平逐渐下降,并于6周龄时接近CK组水平,这可能是由于GABA缓解了雏鸡的热应激,从而使得HS+G组胰腺组织中细胞凋亡水平在3周龄后逐渐下降。CK+G组雏鸡胰腺组织中凋亡细胞数量一直处于较低水平,仅在3周龄时高于CK组,可能是由于雏鸡的快速生长使得胰腺组织中的自然凋亡细胞的数量增加,而4~6周龄时CK+G组雏鸡胰腺组织中细胞凋亡水平处于下调状态,且低于其他3组,可能是由于GABA的长期添加对雏鸡胰腺的发育起到促进作用,缓解了雏鸡胰腺组织中细胞的凋亡。
4 结 论
综上所述,热应激能够改变雏鸡胰腺组织的结构,降低抗氧化酶和消化酶活性,增加细胞凋亡,严重阻碍了雏鸡胰腺的发育进程;GABA的添加可以使热应激雏鸡胰腺组织损伤程度减轻,使雏鸡胰腺组织结构保持完整性,还能提高雏鸡胰腺组织中抗氧化酶和消化酶的活性及其mRNA相对表达量,降低MDA含量。所以,GABA可以作为添加剂来减轻热应激对雏鸡胰腺组织结构、抗氧化能力和消化功能发育的负面影响。
