陈山红心杉1.5代种子园遗传多样性和子代父本分析
2021-06-04陈兴彬徐海宁肖复明孙世武娄永峰邹元熹徐小强
陈兴彬,徐海宁,肖复明*,孙世武,娄永峰,邹元熹,徐小强
(1.江西省林业科学院,江西省植物生物技术重点实验室,江西 南昌 330032; 2.吉安市青原区白云山林场,江西 吉安 343000;3. 吉安市青原区林业局,江西 吉安 343000)
种子园是林木提供良种的主要形式,其目标是稳定提供遗传改良的种子。种子园既要有较高的遗传增益,又要保持较宽的遗传基础,才能确保子代具有较强的适应性[1]。同时,对种子园交配参数进行深入研究,可以了解种子园的花粉污染率、随机交配状况和自交水平等[2],这对种子园的经营和管理有着重要意义。
近交和亲本贡献率是重要的交配参数。近交,特别是自交,常常导致种子园种子产量的下降和种子的生长不良[3]。自交和外界花粉污染能使种子园的遗传增益减少50%以上[4]。海岸松(Pinuspinaster)子代试验林实验表明,近交可使树高降低27%,胸径减少37%,亲本繁殖力降低89%[5]。同时,不均等的亲本贡献不仅降低子代的育种值,还可能降低子代的遗传多样性和适应性[6]。
杉木[Cunninghamialanceolata(Lamb.) Hook]是我国南方最重要的用材树种之一,具有重要的经济和生态价值。杉木种源间存在显著的遗传差异,陈山红心杉是江西特有杉木优良种源[7]。我国自20世纪60年代开始构建杉木1代无性系种子园以来,杉木种子园的营建、种子产量和质量提升等方面取得了显著进步,并在生产上取得了大量成果[8-9]。然而,杉木种子园交配参数还鲜见研究报道。笔者以陈山红心杉1.5代种子园为材料,比较种子园亲子代遗传多样性的差异,并对种子园子代进行父本分析,以期为红心杉种子园良种生产和经营管理提供理论指导。
1 材料与方法
1.1 试验材料
研究材料取自江西省青原区白云山林场陈山红心杉1.5代种子园和其子代测定林。由于杉木的种子成苗率不高,以测定林为材料进行子代父本分析研究,对种子园的经营和管理具有指导意义。20世纪90年代中期,科研人员在陈山林区的杉木人工林与天然林分中选出陈山红心杉优树53株,于1998年营建成陈山红心杉1代种子园,2000年种子园开始生产种子。2001年进行了单系采种,并在赣东北、赣中、赣南进行子代测定工作。根据子代测定结果,选出32个生长快、材性更加优良的无性系用于1.5代种子园的营建。
本试验中的1.5代种子园于2006年4月嫁接建成,面积11 hm2,共32个无性系,小区内无性系配置以随机与调整相结合定植,株行距为 4 m×4 m,同一无性系不同个体间距20 m以上。2017年秋,分单系采种,2018年春育苗,2019年春在白云山林场营造子代测定林。测定林采用随机区组设计,5株单行小区,10次重复,以普通杉木苗为对照。2020年7月,随机选择测定林的14个家系,采集前7个重复单株的针叶,每个家系采集子代数量30~35个不等,共计459个子代个体,同时采集种子园32个亲本的针叶,一并带回实验室放入-70°冰箱中备用。采用改良CTAB法提取基因组DNA。
1.2 PCR扩增及产物检测
利用12对SSR引物对亲本和子代的基因组DNA进行PCR扩增[10],所得扩增产物由北京睿博兴科生物技术有限公司进行毛细管电泳检测。PCR扩增反应程序为:95 ℃预变性5 min;95 ℃变性30 S,56 ℃退火30 S,72 ℃延伸45 S,共进行30个反应循环;72 ℃延伸7 min,4 ℃保存。
1.3 数据处理
利用Gene Marker 2.2.0分析毛细管电泳结果,得到基因型数据。利用Popgen 1.32计算亲本和子代群体的遗传多样性参数。参数包括等位基因数(number of allele,Na)、有效等位基因数(number of effective allele,Ne)、Shannon’s信息指数(Shannon’s information index,I)、观测杂合度(observed heterozygosity,Ho)、期望杂合度(expected heterozygosity,He)和固定指数(fixed index,F,F=1-Ho/He)。应用混合交配模型MLTR 3.4估计多位点异交率(multilocus outcrossing rate,tm)、单位点异交率(singlelocus outcrossing rate,ts)、双亲近交指数(biparental inbreeding coefficient,tm-ts,数值越大表示父母本个体间亲缘关系越近)、多位点父本相关(biparental inbreeding,Rp(m)),单位点父本相关(multilocus correlation of paternity,Rp(s))[11]。Rp是父本相关性的估计,表示家系中异交子代是全同胞的比例。根据Nep=1/Rp(m),计算有效花粉供体数[12]。用软件Cervus 3.0进行子代父本分析[13]。
2 结果与分析
2.1 种子园亲本与子代遗传多样性分析
12对SSR引物在32个亲本中检测到的等位基因数(Na)范围为3~7,平均为4.41个,平均有效等位基因数(Ne)为2.330,其中LX-18位点的Na和Ne最大,分别为7和3.457(表1)。亲本群体的平均Shannon’s信息指数为0.992,实际观察杂合度(Ho)和期望杂合度(He)的均值分别为0.571和0.557,观测杂合度高于期望值杂合度。在包含459个体的子代群体中,12对引物分别检测到4~11个等位基因,平均为6.50个,较亲本群体高出2.09。子代群体的Ne为1.680~3.916,平均为2.306,稍低于亲本群体。子代群体的Shannon’s信息指数为1.004,说明子代群体中遗传多样性略高于亲本群体。子代群体平均Ho和He分别为0.525和0.538,相对于亲本群体略有减少,说明子代群体的杂合个体的比率和基因分布的均匀程度有所降低。子代群体的固定指数(F)为0.024,说明存在纯合子过量现象。在分析的12个位点中,有11个位点检测到1~5个子代特有而亲本没有的等位基因。

表1 亲子群体的遗传多样性
2.2 种子园亲本交配参数分析
多位点交配系统分析表明,陈山红心杉种子园总的多位点异交率(tm)是1.012±0.075,单位点异交率(ts)为0.991±0.042,说明种子园异交率较高。双亲近交指数(tm-ts)为0.021±0.087,表明种子园无性系间近交水平比较低。多位点父本相关性[Rp(m)]和单位点父本相关性[Rp(s)]分别为0.128±0.019和-0.016±0.079,单位点和多位点父本相关性的差值Rp(s)-Rp(m)=-0.144<0,表明亲本间没有明显的近亲关系。种子园的有效花粉供体数目(Nep)为7.81。
对14个家系分别进行交配系统分析(表2),家系间的多位点异交率(tm)变化幅度为0.938~1.200,有10个家系的多位点异交率为1.200,其余4个家系的多位点异交率为0.938~1.114。近交指数(tm-ts)变化幅度为-0.127~0.150,其中5、8、27、29、51号家系近交指数等于或小于零,其余家系的近交系数都大于零,说明部分家系存在近交现象。
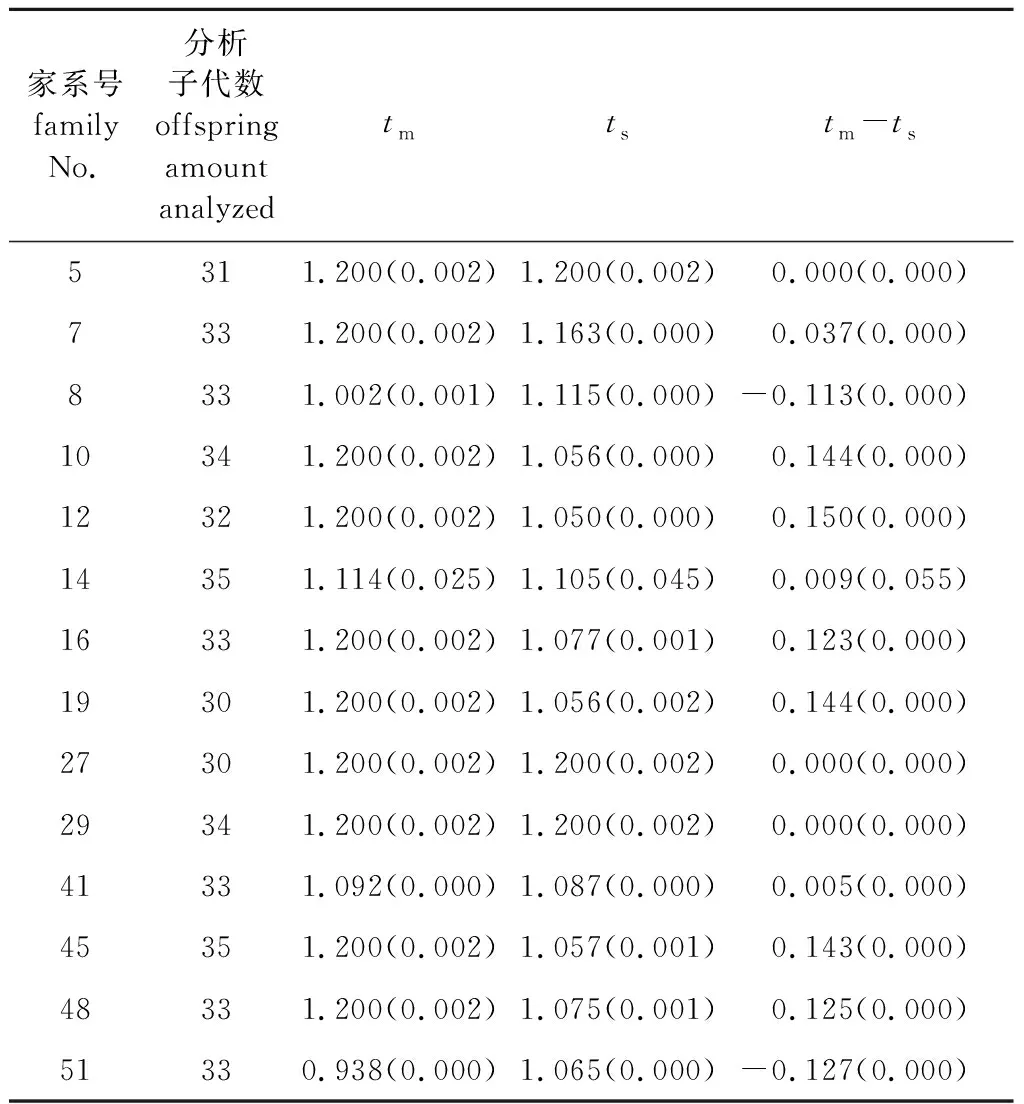
表2 14个家系的异交率
2.3 子代父本分析
对14个家系的459个子代进行父本分析,在80%可信度下推断出325个子代的父本,占子代总数的70.8%(表3)。子代的亲本均不是同一无性系,说明种子园无自交现象。各家系子代确定父本的比率不一致,41号家系子代确定父本的比率最高,为93.9%,其他家系子代确定父本比率为54.3%~90.9%。
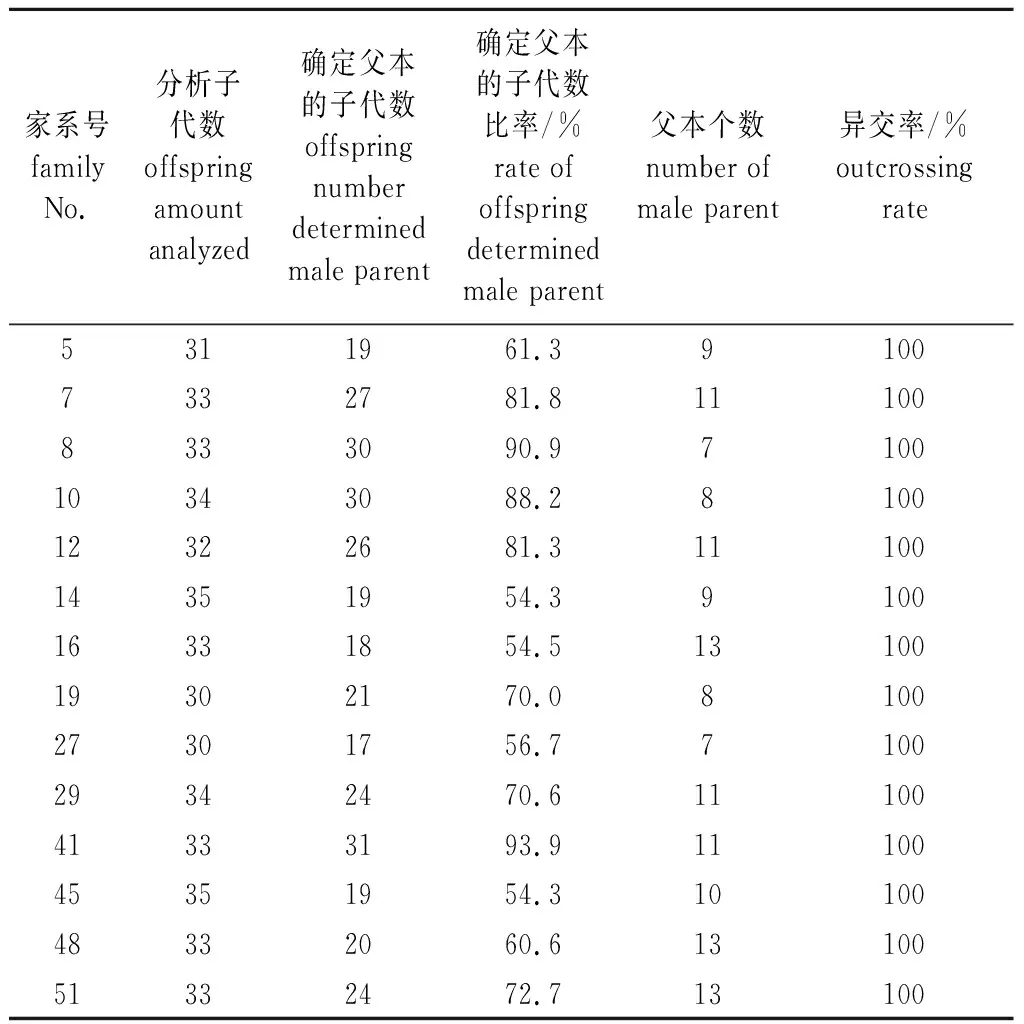
表3 14个家系的子代父本来源分析
对325个子代的父本来源进行分析,可以明确每个家系子代的父本组成(图1)。其中,8号家系确定父本的30个子代中,16个子代的父本为20号无性系,占家系子代总数的53.3%;12号家系确定父本的26个子代中,12个子代的父本为22号无性系,占家系子代总数的46.2%,说明无性系间授粉亲和性不同。在32个潜在无性系中,有26个无性系提供了有效花粉,其中父本贡献率最高的是22号无性系和29号无性系,各为33个子代提供了花粉,其贡献率均为10.2%,其他无性系的父本贡献率为0.3%~8.9%。父本贡献率最高的前11个无性系共计产生了70.2%的子代。
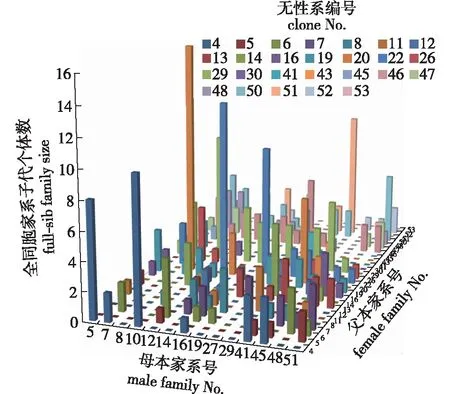
图1 子代父本来源分析Fig.1 Paternal origin of offspring
3 讨 论
遗传多样性包括了一个物种所有个体间遗传变异的总和,是群体生存和发展的前提[14]。对遗传资源进行遗传多样性分析,有利于育种策略的制定和资源的科学管理。种子园所产种子保持一定的遗传多样性,有利于所造林分适应栽培环境。本研究中陈山红心杉1.5代种子园亲本和子代观测杂合度(Ho)分别为0.525和 0.571,说明子代群体中实际杂合单株比率较亲本有所下降,但差异不大。由于亲本间的近交,使得子代杂合性低于亲本。亲本群体和子代群体的Shannon’s信息指数(I)分别为0.992和1.004,说明子代群体的遗传多样性略高于亲本群体。在马尾松(Pinusmassoniana)种子园[15]、华北落叶松(Larixgmeliniivar.principis-rupprechtii)2代种子园[16]均发现种子园子代群体的遗传多样与亲本群体差异不显著。本研究的结果与上述研究结果相一致,即经过1代的自由授粉,种子园子代群体的遗传多样性并没有降低。
异交植物中,近交和自交会使有害隐性基因纯合,从而使自交子代易出现生长不良、抗性差和易致死等现象[17]。风媒传粉的针叶树种子园一般具有较高的异交率,如,马尾松的异交率为109.8%[18],海岸松的异交率为90.1%[3],日本柳杉 (Cryptomeriajaponica) 的异交率为94.4%[19]。本研究表明,陈山红心杉种子园子代测定林没有自交子代,多位点异交率(tm)为1.012,说明种子园具有较高的异交水平。异交率高可能与所用的实验材料有关,本研究材料为子代测定林,杉木自交导致种子败育,成苗率极低[20],自交子代生长较弱,可能在种子形成期和幼苗期已经被淘汰[21],因此没有发现自交子代。陈山红心杉种子园具有高的异交率,但仍存在少量的近交现象。双亲近交指数(tm-ts)为0.021,表明无性系间亲缘关系较低,只有低水平的近交发生,与得出的较高的有效花粉供体数目(Nep=7.81)相一致。无性系间不同的花期、花量以及选择受精均可导致种子园非随机交配,造成近交的发生[22]。
无性系提供均等的生殖贡献有助于提高种子园种子的遗传品质,因此,大量树种开展了父本贡献率研究[23]。云杉(Piceaasperata)种子园多年研究表明,平均10.4%无性系生产了种子园50%的子代[24]。日本落叶松(Larixkaempferi)种子园研究表明,无性系的父本贡献率为0~12.4%,50%的无性系提供了91.7%子代的花粉[25]。本研究在80%可信度下推断出325个子代的父本,在32个潜在父本无性系中,有26个无性系提供了有效花粉,无性系的父本贡献率为0~10.2%,父本贡献率最高的前11个无性系共产生70.2%的子代。因此,验证了前人研究得出的大部分无性系种子园普遍存在非随机交配现象[26]。该种子园花期物候观测发现,无性系间花期、花量均存在较大差异,无性系间生殖能力的差异可能是父本贡献率差异大的原因之一。陈山红心杉不同无性系间还存在授粉偏好性,如,8号家系确定父本的30个子代中,有16个子代的父本为20号无性系;12号家系确定父本的26个子代中,有12个子代的父本为22号无性系。下一步需研究亲和力高的无性系育种值状况,新建种子园时,通过增加亲和力和育种值均高无性系的数量,适当减少亲和力高,但育种值低的无性系数量,以建设高生产力红心杉无性系种子园。
