铁皮石斛WOX转录因子的鉴定和分析
2021-06-03曾丹琦张明泽何春梅王浩斌俞振明司灿赵聪慧李冬妹段俊
曾丹琦, 张明泽, 何春梅, 王浩斌, 俞振明, 司灿, 赵聪慧,李冬妹, 段俊*
铁皮石斛WOX转录因子的鉴定和分析
曾丹琦1,2, 张明泽1,2, 何春梅1, 王浩斌1,2, 俞振明1, 司灿1,2, 赵聪慧1,2,李冬妹3, 段俊1*
(1. 中国科学院华南植物园,广东省应用植物学重点实验室,中国科学院华南农业植物分子分析与遗传改良重点实验室,广州 510650;2. 中国科学院大学,北京 100049;3. 顺德职业技术学院,广东 佛山 528300)
为了解铁皮石斛() WOX转录因子的功能,采用全基因组分析技术对铁皮石斛WOX家族成员进行鉴定,并进行生物信息学和表达模式分析。结果表明,铁皮石斛基因组中有9个WOX转录因子(DoWOX 1DoWOX 9),大部分的含有2~3个外显子,启动子含有与激素诱导、逆境胁迫和生长发育有关的顺式作用元件。qPCR分析表明,、、、和在类原球茎中的表达量最高,在小花蕾中表达最高, 随着花的发育,表达呈现下降趋势。此外,、和在合蕊柱上的表达量最高,而在唇瓣的表达量最高。因此,可能参与调控铁皮石斛的生长发育,且可能在维持类原球茎状态和花的发育中起重要作用。
铁皮石斛;WOX转录因子;生物信息学;基因表达;qPCR
植物展示出非凡的再生能力,保证发育的可塑性。植物干细胞是未分化的细胞,位于分生组织, 未分化的状态为稳定的遗传性、较高的线粒体活性以及很强的分化和自我更新能力等奠定了基础[1-2],进而对环境变化信号快速响应和对植物发育进程参与调控[3]。植物细胞全能性在生物技术上具有广泛的应用市场。目前对于植物的干细胞研究主要在拟南芥()等的茎尖干细胞[4-5]、水稻()等的根尖干细胞[6]上,植物干细胞发生机理在种苗生产、生物技术等领域扮演举足轻重的角色。
WOX家族(WUSCHEL-related homeobox)是真核细胞homeobox (HB)超家族中的一员[7-8]。WOX转录因子具有“螺旋-环-螺旋-转角-螺旋”的典型结构[8-9],在植物发育过程中扮演重要的调控角色[10-11],包括在植物干细胞维持、胚胎形成及茎干、根和花的发育等。有研究表明,在根和茎分生组织中干细胞生态位的发育上具有重要作用[11-12];植物的胚胎发育是个复杂的过程,不仅受到运输调控, 而且受到局部生长素反应的影响[13],可以决定植物胚胎发育的细胞命运[12];在根发育中,在不定根基源细胞中表达[12,14]以激活LBD16,进而保证根的正常发育;在形成层干细胞中,和其余WUS成员在干细胞维持中起重要作用[15-16]; WOX转录因子在花的结构和发育上具有重要的调控作用[17],被认为是花干细胞维持必需的基因[17-18],在矮牵牛()和烟草()中是保证花器官结构正常的必需基因[19-20]。WUS家族蛋白成员可以分为3个进化支: 分别为WUS、Ancient和Intermediate[8],但对其功能还不清楚。
铁皮石斛()为兰科(Orchi- daceae)石斛属草本植物,有较高的药用价值, 具有抗肿瘤、增加免疫力和益胃生津等功效[21-22]。由于对过度采挖导致野生资源大量减少,因此从分子水平研究铁皮石斛的生长发育对保护其资源具有重大意义。目前关于WOX转录因子的功能研究主要在模式植物拟南芥和水稻上,对中药材如铁皮石斛[23-24]的相关研究鲜见报道。此外,兰科植物的花在结构上高度特化,如具有形态特异的萼片、唇瓣和合蕊柱[25]。本研究根据已报道的铁皮石斛基因组数据[26],对其WOX家族进行鉴定和生物信息学分析,并分析基因在不同生长发育阶段和花的不同组织中的表达模式,为探讨WOX转录因子在调控铁皮石斛生长发育中的作用提供理论依据。
1 材料和方法
1.1 材料
铁皮石斛()种植于中国科学院华南植物园苗圃。取约1 cm长的花蕾,分别收集萼片、花瓣、唇瓣和合蕊柱,液氮速冻15 min,保存于-80℃冰箱备用。
铁皮石斛组培苗在1/2MS+0.1%活性炭+2%蔗糖+0.6%琼脂培养基(pH 5.4)上培养,培养条件为(26±1)℃、32.57mol/(m2·s)、12 h光照/12 h黑暗, 相对湿度约为60%。采集的类原球茎、丛生芽(无根)和小苗(约3 cm高)经液氮速冻后保存于-80℃冰箱备用。
1.2 WOX转录因子家族成员的鉴定和序列分析
从NCBI (https://ftp.ncbi.nlm.nih.gov)基因组数据库下载铁皮石斛基因组文件,利用Pfam、Swiss_Prot、eggNOG_class、NR注释结果检索WOX转录因子。然后对检索到的WOX蛋白序列与拟南芥WOX蛋白序列进行比对,并构建进化树,删除同源性低且遗传距离较远,剩余基因即为铁皮石斛。
1.3 多重序列比对和进化树分析
采用DNAMAN8软件对铁皮石斛WOX家族成员的氨基酸序列进行比对。采用Clastal X version 2.0软件对铁皮石斛的WOX蛋白进行比对,输出Fasta格式文件,导入MEGA,利用邻接法(Neighbor- Joining)构建系统发育树(NJ树),进行1 000次重复bootstrap统计学检测。
1.4 WOX的生物信息学分析
从铁皮石斛全基因组测序GFF文件中获得的GFF注释,利用GSDS在线软件(http://gsds. cbi.pku.edu.cn/)绘制铁皮石斛WOX家族成员的基因结构。利用在线工具MEME (http://alternate.meme- suite.org/tools/meme)搜索铁皮石斛WOX基因家族成员的motif,用PlantCare在线软件(http://bioinfor-matics.psb.ugent.be/webtools/plantcare/html/)预测启动子中的顺式作用元件,并用TBtools制图。
1.5 RNA的提取和反转录
参照柱式植物RNAOut 2.0试剂盒(北京天恩泽基因科技有限公司)说明书,分别提取铁皮石斛不同发育时期(类原球茎、丛生芽和小苗)和花的不同组织(萼片、花瓣、唇瓣和合蕊柱)的总RNA。提取的RNA经DNA酶消化后,取2L进行电泳检测,再采用Promega公司的GoScript™ Reverse Transcription System试剂盒,取4g已纯化的总RNA进行反转录,反应体系为20L。
1.6 表达分析
采用美国BIO-RAD公司iTaq Universl SYBR Green supermix反应试剂对9个进行实时荧光定量PCR (qPCR)分析。使用Primer Premier 5.0软件在基因特异性区域设计引物(表1),以铁皮石斛为内参基因(NCBI accession No.: JX294908), 于ABI 7500 Real-time PCR仪上进行qPCR,并使用其系统软件对数据进行处理, 采用2−ΔΔCT方法[27]计算相对表达量。铁皮石斛花的发育分为3个时期:小花蕾期(约0.5 cm)、中花蕾期(约1 cm)和全开期。采用qPCR对在花不同发育阶段和不同组织的表达模式进行分析。用百迈客云平台(http://www. biocloud.net/)在线软件绘制热图(heatmap)。
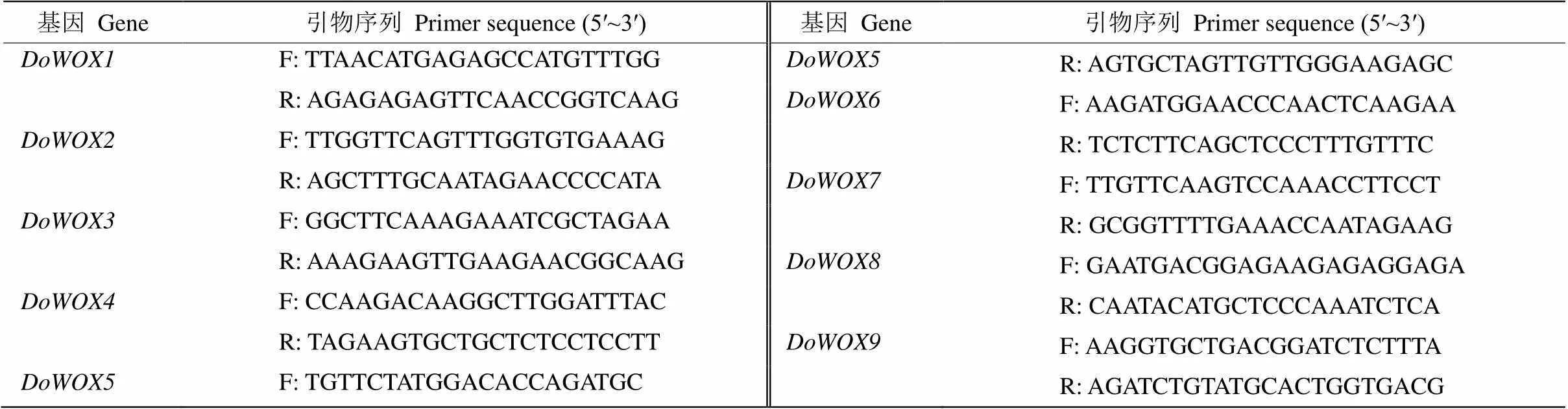
表1 实时荧光定量PCR所用引物
2 结果和分析
2.1 WOX转录因子的鉴定和进化树构建
在NCBI基因组数据库下载铁皮石斛基因组文件,并利用Pfam、Swiss_Prot、eggNOG_class、NR注释结果检索WOX转录因子,共鉴定出9个铁皮石斛WOX家族基因成员,分别命名为、、、、、、、和。
以铁皮石斛WOX家族蛋白和拟南芥WOX家族蛋白构建进化树(图1),结果表明, 铁皮石斛的WOX家族可分成3个进化支,分别为WUS、Ancient和Intermediate,其中WUS的成员数量最多,分别是DoWOX3、DoWOX4、DoWOX5、DoWOX6和DoWOX9;其次是Intermediate,分别是DoWOX1、DoWOX2和DoWOX7;Ancient的数量最少,仅有DoWOX8。此外,WUS支的DoWOX4和DoWOX5的分支长度差比其他DoWOXs间的分支长度差短、置信度最高,说明DoWOX4和DoWOX5亲缘关系最近、同源性最高,暗示他们具有相似的功能。
2.2 DoWOXs的生物信息学分析
使用DNAMAN的默认参数对9个DoWOXs进行多重序列比对,结果表明,铁皮石斛WOX家族成员均含有保守的同源异型结构域(homeobox domain, HD),具有“螺旋-环-螺旋-转角-螺旋”的结构特征(图2)。此外,在螺旋I和螺旋III中存在许多高度保守的位点,暗示这些氨基酸残基可能在基因功能上发挥重要的作用。
利用MEME工具对铁皮石斛WOX家族成员的蛋白结构进行分析(图3), 结果表明,所有DoWOXs都含有串联的Motif1和Motif2,说明其在WOX家族成员中较为保守。在Intermediate成员中存在Motif3的保守基序;WUS家族成员中除DoWOX9外,其余成员在C端都有保守的Motif4;Ancient成员DoWOX8含有的保守基序最少,说明WOX可能具有复杂的进化过程。此外,同一亚家族中的DoWOX4和DoWOX5的保守基序都是5个,且位置相近,表现了很强的一致性,说明DoWOX4和DoWOX5具有相似的蛋白质功能和结构,同时从侧面反映了系统进化树分析的可靠性。
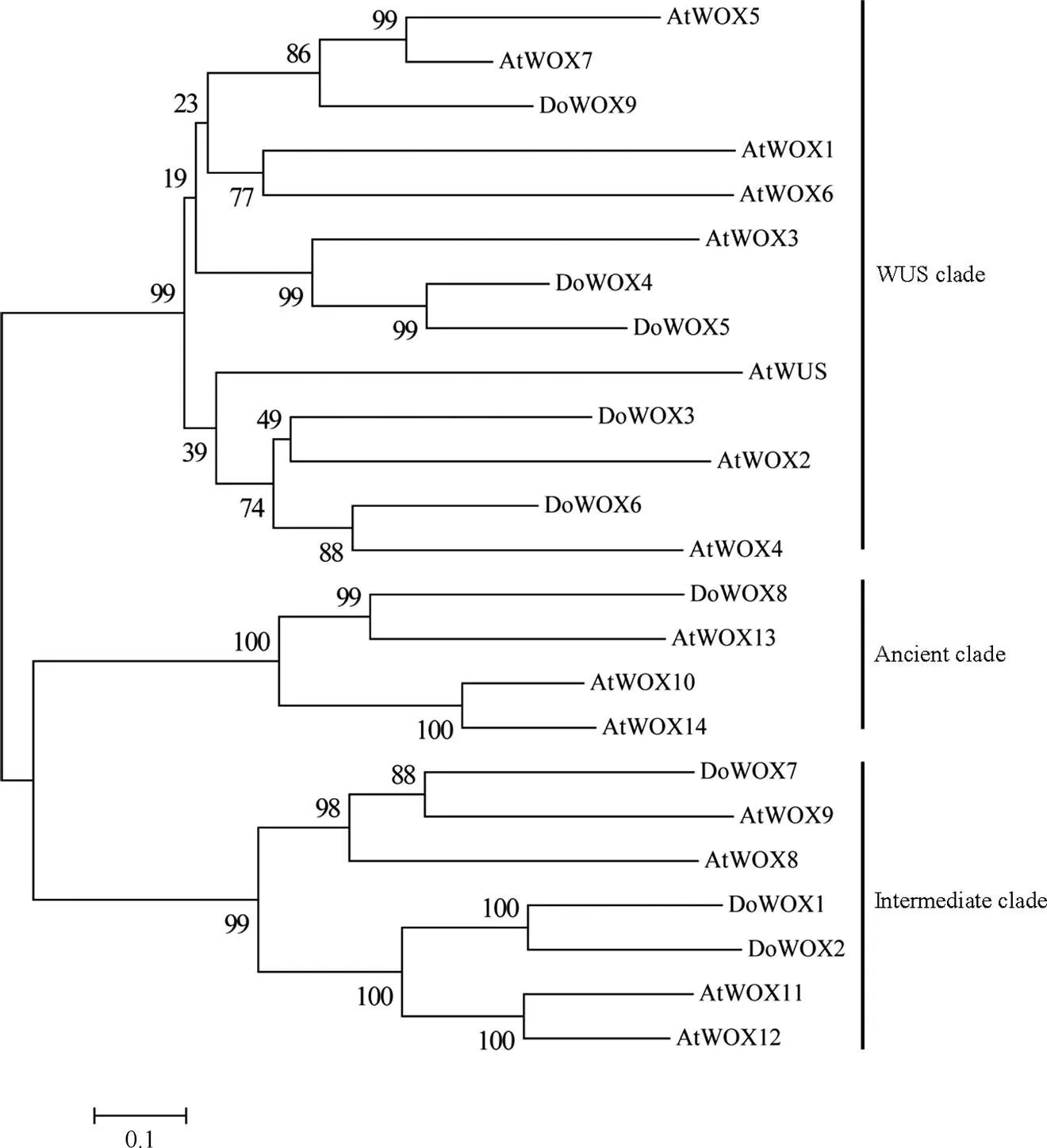
图1 铁皮石斛(Do)和拟南芥(At)的WOX家族成员构建的系统进化树

图2 铁皮石斛WOX家族成员氨基酸序列的多重比对
2.3 DoWOXs基因结构
利用在线软件GSDS构建铁皮石斛基因的内含子-外显子结构图(图4),可见,大部分有内含子1~3个,外显子1~3个。结合系统进化树,不同进化分支的基因结构差异明显, 即使是相同分支的基因间也存在一定差异,但同源性最高的和的基因结构极其相似。此外,同为WUS支的,其基因结构与和相似,皆为2个外显子,1个内含子,说明WUS支的基因结构较为保守。Ancient支中的有3个外显子和2个内含子。
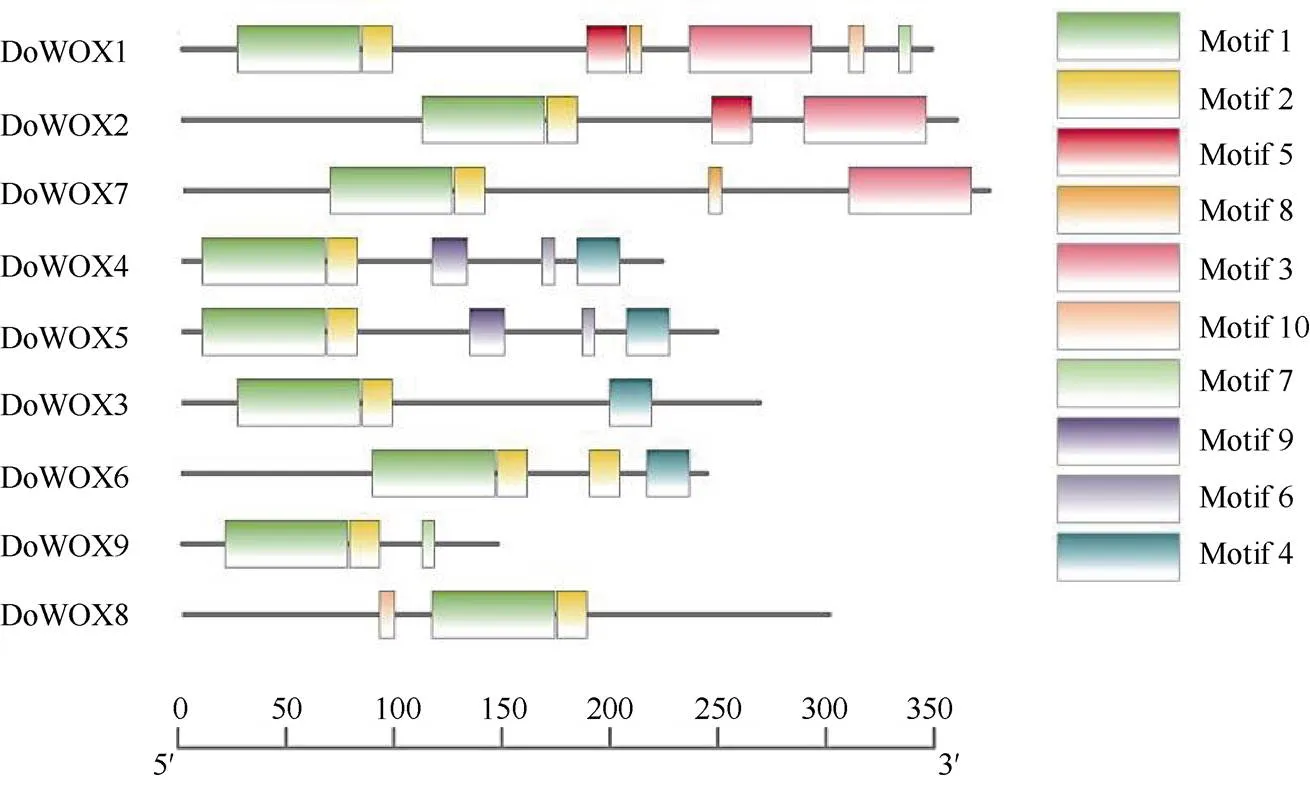
图3 DoWOXs蛋白的保守基序
对的启动子用PlantCARE进行分析(图5),结果表明,9个共含有13个与生长发育相关的顺式作用元件,不同基因间的顺式作用元件不同。其中,顺式作用元件meristem expre- ssion共有6个,含有3个,说明其可能在铁皮石斛分生组织的生长上扮演重要角色。与激素相关的顺式作用元件有auxin-responsive、gibbe- rellin-responsive、MeJA-responsive、abscisic acid responsive、salicylic acid responsive等,在植物和环境胁迫的适应中扮演重要角色;与逆境胁迫相关的非生物激素的顺式作用元件有drought-inducibility、defense and stress responsive、low-temperature responsive和anaerobic induction等,间接影响生长发育。这说明WOX家族成员在铁皮石斛生长发育过程中,参与胚胎发育、种子萌发、分生组织生长、根系发育和环境胁迫的调控等活动。
2.4 DoWOXs的表达
铁皮石斛类原球茎被认为是体胚[28],可增殖, 亦可分化成完整的植株,而WOX成员在维持植物的干细胞状态中起着重要的作用。采用qPCR技术分析了在铁皮石斛不同发育阶段中的表达模式(图6),结果表明,除外其余在类原球茎、丛生芽和小苗中有表达。、、、和在类原球茎中的表达量最高,表明WOX在铁皮石斛的类原球茎发育中扮演重要角色。在丛生芽中的表达量最高,在小苗中最高,和在3个不同发育时期表达量均较高。此外,不同发育时期优势表达的不同,如在类原球茎特异表达,可能与其具有不同的启动子顺式作用元件有关。
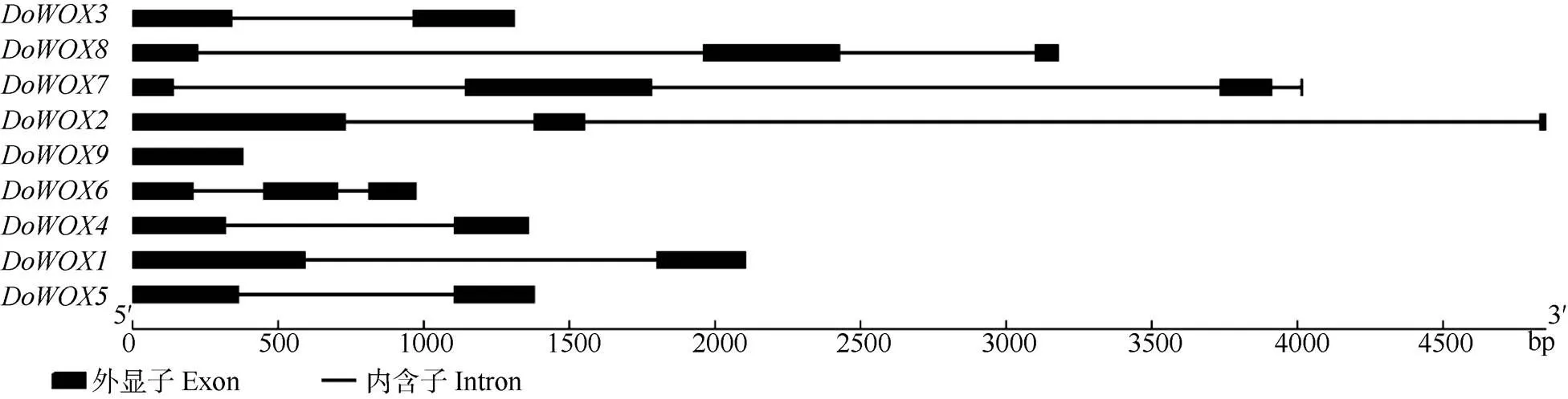
图4 DoWOXs基因结构
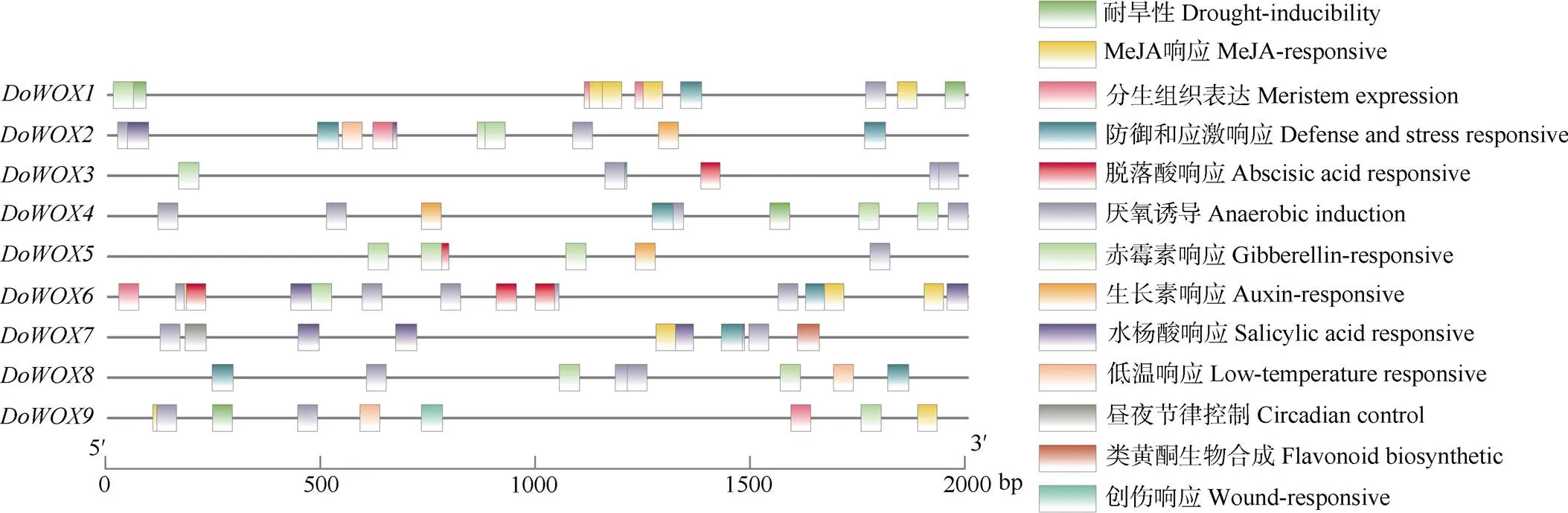
图5 DoWOXs启动子中的顺式作用元件
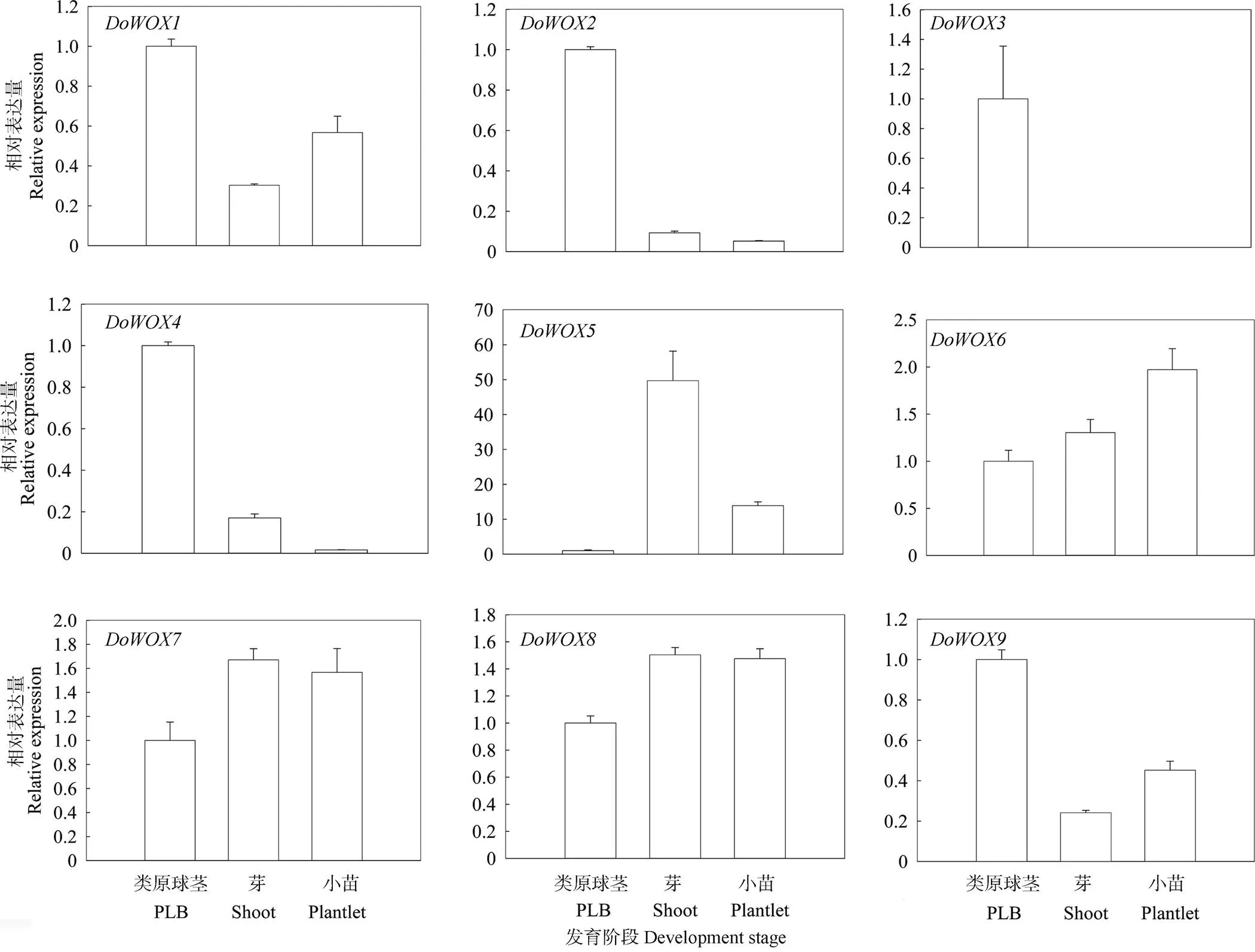
图6 DoWOXs基因在铁皮石斛不同发育时期的表达
开花是植物发育中非常重要的阶段,WOX参与植物花发育过程[18]。利用本实验室建立的铁皮石斛花发育数据库,获取了在小花蕾(S1)、中花蕾(S2)和全开花(S3)中的表达数据,利用百迈客云平台在线软件绘制热图(图7),可见,、的表达模式较为相似,在S1时期的表达量最高,其次为S2时期,S3时期最低,暗示这2个基因在花发育过程中功能相似。在3个时期的FPKM值均大于20,S2时期的表达量最高,而其他在3个时期的FPKM均小于5。

图7 DoWOXs在铁皮石斛不同开花期的表达谱。S1: 小花蕾; S2: 中花蕾; S3: 全开花; 红色: 上调; 绿色: 下调。
兰科植物的花具有较高的观赏价值主要源于其独特的花型,有3个瓣化的萼片,2个花瓣,1个唇瓣和1个合蕊柱[29]。进一步选取S2时期的花蕾进行不同组织的表达分析,结果表明(图8),和在花蕾的4个组织中均未表达;仅在唇瓣和合蕊柱中表达;其余6个在中花蕾的不同组织中均有表达,其中在唇瓣中表达较高,在萼片中表达最高,在花瓣中表达最高,和在合蕊柱中表达最高。这暗示家族成员在铁皮石斛花生长发育过程中具有不同的调控作用。此外,和在合蕊柱的表达量远高于其他花器官,可能与花形态发育密切相关。
3 结论和讨论
参与调控植物发育的基因在形态建成中起着关键的作用。WOX转录因子是参与植物发育的重要家族之一。本研究从铁皮石斛全基因组中鉴定出9个WOX转录因子(DoWOX1~DoWOX9),生物信息学和表达分析表明,WOX转录因子在铁皮石斛的生长发育尤其是类原球茎和花发育中起重要的调控作用。和在开花时期表达量的变化规律一致,且DoWOX4和DoWOX5氨基酸序列相似度高(图1),暗示这2个转录因子可能在调控花发育过程中的功能相似。此外,DoWOX4、DoWOX5和AtWOX3在系统进化树上的亲缘关系最近,AtWOX3在拟南芥花发育尤其是花表型构成中起作用[18]。
9个编码蛋白的同源异型结构域(HD)保守性高,具有典型的“螺旋-环-螺旋-转角-螺旋”结构,这与其他物种[30-31]的研究结果一致,表明该同源异型结构域在进化上具有较高的保守性。该同源异型结构域可以特异地和DNA序列结合而发挥重要的作用[32],如作为HOX蛋白的辅助因子影响植物、动物和真菌的发育[8]。基因结构分析表明, WUS支的成员基因结构较为保守。不同支系DoWOXs蛋白的保守基序在数量、位置上具有较高的相似性,这与系统进化树的结果一致。此外,Motif1和Motif2基序的保守程度更高。
基因的表达调控是一个复杂的过程,转录因子通过结合基因的顺式作用元件来调控基因的表达。的启动子上含有响应植物激素和逆境胁迫等相关的顺式作用元件,生长素可调控胚胎发育[33]、脱落酸影响种子休眠和萌发[34],茉莉酸甲酯(MeJA)参与植物防御反应[35],因此,可能参与植物生长发育和防御反应。
类原球茎、芽和小苗是铁皮石斛3个典型的生长发育阶段。5个在类原球茎中高表达, 说明在调控铁皮石斛类原球茎发育具有重要的作用,这与梁易等[24]的研究结果一致。类原球茎发育是铁皮石斛的特有阶段。在类原球茎中表达量最高,在芽和小苗中不表达,与同源性最高的的表达模式相似[36]。有大量研究表明,在植物的胚胎发育中扮演不同的角色,如
和影响拟南芥的胚胎模式[12],调控云杉()的体细胞胚胎发生[37]。在芽中表达最高,可能与铁皮石斛芽发育有关。参与调控植物芽的发育,如在拟南芥芽再生发育过程是必需的[38]、通过招募相关蛋白影响水稻芽的发育[39]。除外,其余在小苗中均有表达,说明可能调控铁皮石斛小苗发育。花发育是植物生殖阶段的重要过程,在小花蕾、中花蕾和全开花时期都有表达,说明在铁皮石斛花发育中起着重要的调控作用。调控花发育的作用在拟南芥、矮牵牛和烟草等植物中已有报道[17-20]。除和外,其余均在中花蕾不同组织中表达,说明在中花蕾时期和可能和其他存在功能冗余的情况,具体功能还需进一步研究。合蕊柱是兰科植物花的独特结构,和在合蕊柱中的表达量远远高于其他组织,推测两者调控铁皮石斛花的发育,但具体功能有待进一步研究。同一亚家族成员存在一些相似的功能,有研究表明WUS分支的其他成员存在不同程度替代WUS在芽和花干细胞维持方面的功能[16],这为寻找WOX家族的共性提供一个突破口。
植物组织培养技术是目前解决铁皮石斛种苗较难繁殖的主要方式,干细胞的形成与分化是组织培养中的重要环节,因此,研究铁皮石斛干细胞的相关分子机理具有十分重要的意义。兰科植物的花是重要的生殖器官,而且具有较高的观赏价值,研究兰科植物花器官发育对了解其花形成机制及品种选育提供了重要的理论基础。本研究为进一步挖掘铁皮石斛WOX转录因子功能奠定了基础。
[1] SCHERES B. Stem-cell niches: Nursery rhymes across kingdoms [J]. Nat Rev Mol Cell Biol, 2007, 8(5): 345-354. doi: 10.1038/nrm2164.
[2] LAUX T. The stem cell concept in plants: A matter of debate [J]. Cell, 2003, 113(3): 281-283. doi: 10.1016/s0092-8674(03)00312-x.
[3] LUO L J, ZENG J, TIAN Z X, et al. Plant development: From cells to individuals [J]. Chin Sci Bull, 2016, 61(33): 3532-3540. doi: 10.1360/ N972016-00906.罗林杰, 曾健, 田朝霞, 等. 植物的发育: 从细胞到个体[J]. 科学通报, 2016, 61(33): 3532-3540. doi: 10.1360/N972016-00906.
[4] SANG Y L, CHENG Z J, ZHANG X S. Plant stem cells andorganogenesis [J]. New Phytol, 2018, 218(4): 1334-1339. doi: 10.1111/ nph.15106.
[5] SCHOOF H, LENHARD M, HAECKER A, et al. The stem cell popu- lation ofshoot meristems is maintained by a regulatory loop between theandgenes [J]. Cell, 2000, 100(6): 635-644. doi: 10.1016/S0092-8674(00)80700-X.
[6] YU Y M, LEE E K, HONG S M, et al. Plant stem cell line derived from quiescent center and method for isolating the same: EP, 20080832681 [P]. 2010-05-26.
[7] GEHRING W J. Homeo boxes in the study of development [J]. Science, 1987, 236(4806): 1245-1252. doi: 10.1126/science.2884726.
[8] VAN DER GRAAFF E, LAUX T, RENSING S A. The WUS homeobox- containing (WOX) protein family [J]. Genome Biol, 2009, 10(12): 248. doi: 10.1186/gb-2009-10-12-248.
[9] LIAN G B, DING Z W, WANG Q, et al. Origins and evolution of WUSCHEL-related Homeobox protein family in plant kingdom [J]. Sci World J, 2014, 2014: 534140. doi: 10.1155/2014/534140.
[10] MUKHERJEE K, BROCCHIERI L, BÜRGLIN T R. A comprehensive classification and evolutionary analysis of plant Homeobox genes [J]. Mol Biol Evol, 2009, 26(12): 2775-2794. doi: 10.1093/molbev/msp201.
[11] SARKAR A K, LUIJTEN M, MIYASHIMA S, et al. Conserved factors regulate signalling inshoot and root stem cell organizers [J]. Nature, 2007, 446(7137): 811-814. doi:10.1038/nature 05703.
[12] HAECKER A, GROß-HARDT R, GEIGES B, et al. Expression dyna- mics ofgenes mark cell fate decisions during early embryonic patterning in[J]. Development, 2004, 131(3): 657-668. doi:10.1242/dev.00963.
[13] HAMANN T, BENKOVA E, BÄURLE I, et al. Thegene encodes an auxin response protein inhibiting MONOPTEROS-mediated embryo patterning [J]. Genes Dev, 2002, 16(13): 1610-1615. doi:10.1101/gad.229402.
[14] SHENG L H, HU X M, DU Y J, et al. Non-canonical-mediated root branching contributes to plasticity inroot system architecture [J]. Development, 2017, 144(17): 3126-3133. doi:10.1242/ dev.152132.
[15] SUER S, AGUSTI J, SANCHEZ P, et al.imparts auxin respon- siveness to cambium cells in[J]. Plant Cell, 2011, 23(9): 3247-3259. doi:10.1105/tpc.111.087874.
[16] DOLZBLASZ A, NARDMANN J, CLERICI E, et al. Stem cell regu- lation bygenes [J]. Mol Plant, 2016, 9(7): 1028- 1039. doi:10.1016/j.molp.2016.04.007.
[17] LAUX T, MAYER K F X, BERGER J, et al. Thegene is required for shoot and floral meristem integrity in[J]. Development, 1996, 122(1): 87-96.
[18] COSTANZO E, TREHIN C, VANDENBUSSCHE M. The role ofgenes in flower development [J]. Ann Bot, 2014, 114(7): 1545- 1553. doi:10.1093/aob/mcu123.
[19]VANDENBUSSCHE M, HORSTMAN A, ZETHOF J, et alDifferential recruitment oftranscription factors for lateral development and organ fusion in petunia and[J]. Plant Cell, 2009, 21(8): 2269-2283. doi: 10.1105/tpc.109.065862.
[20] MCHALE N A, MARCOTRIGIANO M.is required for dorso- ventrality and lateral growth of the leaf blade in[J]. Development, 1998, 125(21): 4235-4243.
[21] Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China, Vol. 2 [M]. Beijing: China Medical Science Press, 2015: 282-283.国家药典委员会. 中华人民共和国药典, 第2部 [M]. 北京: 中国医药科技出版社, 2015: 282-283.
[22] ZHANG M Z, HE C M, WANG H B, et al. Advances in active polysaccharides in medicinal plants of Orchidaceae [J]. J Trop Subtrop Bot, 2019, 27(5): 611-622. doi: 10.11926/jtsb.4073.张明泽, 何春梅, 王浩斌, 等. 兰科药用植物活性多糖研究进展 [J]. 热带亚热带植物学报, 2019, 27(5): 611-622. doi: 10.11926/jtsb.4073.
[23] RAMKUMAR T R, KANCHAN M, UPADHYAY S K, et al. Identification and characterization of WUSCHEL-related homeobox() gene family in economically important orchid speciesand[J]. Plant Gene, 2018, 14: 37-45. doi:10.1016/j.plgene.2018.04.004.
[24] LIANG Y. Expression analysis ofandinduring protocorm development[D]. Chengdu: Southwest Jiaotong University, 2018: 1-61.梁易. 铁皮石斛原球茎发育过程中与的表达分析 [D]. 成都: 西南交通大学, 2018: 1-61.
[25] XIANG L, LI B J, QIN D H, et al. Advances in molecular biology of orchid floral development [J]. Chin J Cell Biol, 2011, 33(5): 554-563.向林, 李伯钧, 秦德辉, 等. 兰花花发育的分子生物学研究进展 [J]. 中国细胞生物学学报, 2011, 33(5): 554-563.
[26] ZHANG G Q, LIU K W, LI Z, et al. Thegenome and the evolution of orchids [J]. Nature, 2017, 549(7672): 379-383. doi:10. 1038/nature23897.
[27] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod [J]. Methods, 2001, 25(4): 402-408. doi:10.1006/meth.2001.1262.
[28] LEE Y I, HSU S T, YEUNG E C. Orchid protocorm-like bodies are somatic embryos [J]. Amer J Bot, 2013, 100(11): 2121-2131. doi:10. 3732/ajb.1300193.
[29] RUDALL P J, BATEMAN R M. Roles of synorganisation, zygo- morphy and heterotopy in floral evolution: The gynostemium and labellum of orchids and other lilioid monocots [J]. Biol Rev, 2002, 77(3): 403-441. doi:10.1017/S1464793102005936.
[30] WANG M M, LIU M M, RAN F, et al. Global analysis oftranscription factor gene family inreveals their stress- and hormone-responsive patterns [J]. Int J Mol Sci, 2018, 19(11): 3470. doi: 10.3390/ijms19113470.
[31] LI X X, LIU C, LI W, et al. Genome-wide identification, phylogenetic analysis and expression profiling of thefamily genes in[J]. Hereditas, 2016, 38(5): 444-460. doi: 10.16288/j.yczz.15-499.李晓旭, 刘成, 李伟, 等. 番茄转录因子家族的鉴定及其进化、表达分析 [J]. 遗传, 2016, 38(5): 444-460. doi:10.16288/j.yczz.15-499.
[32] GEHRING W J, MÜLLER M, AFFOLTER M, et al. The structure of the homeodomain and its functional implications [J]. Trends Genet, 1990, 6: 323-329. doi:10.1016/0168-9525(90)90253-3.
[33] SU Y H, ZHAO X Y, LIU Y B, et al. Auxin-inducedexpression is essential for embryonic stem cell renewal during somatic embryo- genesis in[J]. Plant J, 2009, 59(3): 448-460. doi:10.1111/ j.1365-313X.2009.03880.x.
[34] DU Z Y, CHEN M X, CHEN Q F, et al.acyl-CoA-binding protein ACBP1 participates in the regulation of seed germination and seedling development [J]. Plant J, 2013, 74(2): 294-309. doi:10.1111/ tpj.12121.
[35] DU M M, ZHAI Q Z, DENG L, et al. Closely related NAC trans- cription factors of tomato differentially regulate stomatal closure and reopening during pathogen attack [J]. Plant Cell, 2014, 26(7): 3167- 3184. doi:10.1105/tpc.114.128272.
[36] PÉRET B, DE RYBEL B, CASIMIRO I, et al.lateral root development: An emerging story [J]. Trends Plant Sci, 2009, 14(7): 399-408. doi:10.1016/j.tplants.2009.05.002.
[37] PALOVAARA J, HAKMAN I. Conifer-related homeodomain transcription factors, developmental consideration and expression dynamic ofduringsomatic embryogenesis [J]. Plant Mol Biol, 2008, 66(5): 533-549. doi:10.1007/s11103-008-9289-5.
[38] ZHANG T Q, LIAN H, ZHOU C M, et al. A two-step model for de novo activation ofduring plant shoot regeneration [J]. Plant Cell, 2017, 29(5): 1073-1087. doi:10.1105/tpc.16.00863.
[39]CHENG S F, TAN F, LU Y, et al. WOX11 recruits a histone H3K27me3 demethylase to promote gene expression during shoot development in rice [J]. Nucl Acids Res, 2018, 46(5): 2356-2369. doi:10.1093/nar/ gky017.
Identification and Analysis of WOX Transcription Factor in
ZENG Dan-qi1,2, ZHANG Ming-ze1, HE Chun-mei1, WANG Hao-bin1,2, YU Zhen-ming1, SI Can1,2, ZHAO Cong-hui1,2, LI Dong-mei3, DUAN Jun1*
(1. Guangdong Provincial Key Laboratory of Applied Botany, Key Laboratory of South China Agricultural Plant Molecular Analysis and Gene Improvement, South China Botanical Garden, Chinese Academy of Sciences,Guangzhou 510650, China; 2. University of Chinese Academy of Sciences,Beijing 100049, China; 3. Shunde Polytechnic,Foshan 528300, Guangdong, China)
In order to understand the function of transcription factor WOX in, the WOX family members inwere identified by whole genome analysis techniques, the bioinformatics of WOXs and expression patterns ofwere analyzed. The results showed that there were ninetranscription factors ingenome, named DoWOX 1to DoWOX 9, respectively. The most ofcontained 2-3 exons, whilehad only one exon. The promoters ofcontained-regulatory elements related to hormone induction, stress responsive, plant growth and development. The expression of,,,andwere the highest at protocorm-like body (PLB) stage by qPCR, which ofwas the highest at small flower bud stage, and then decreased. In addition, the expressions of,andwere the highest in columns, and that ofwas the highest in lips. Therefore,might be involved in regulating the growth and development of,and play an important role in maintenance of PLBs state and flower development.
; WOX transcription factor; Bioinformatics analysis; Gene expression; qPCR
10.11926/jtsb.4294
2020-08-18
2020-09-23
中国科学院华南农业植物分子分析与遗传改良重点实验室开放课题(KF202008);广州市科技计划项目(201707010345)资助
This work was supported by the Open Project of Key Laboratory of South China Agricultural Plant Molecular Analysis and Genetic Improvement, South China Botanical Garden, Chinese Academy of Sciences (Grant No. KF202008); and the Project for Science and Technology Planning in Guangzhou (Grant No. 201707010345).
曾丹琦(1997~ ),女,硕士研究生,从事铁皮石斛生物技术育种研究。E-mail: ZengDanqi20@scbg.ac.cn
Corresponding author. E-mail: duanj@scib.ac.cn
