海南省入侵杂草假高粱种子萌发特性研究
2021-06-03王玮倩李晓霞王亚黄乔乔
王玮倩, 李晓霞, 王亚, 黄乔乔*
海南省入侵杂草假高粱种子萌发特性研究
王玮倩1,2, 李晓霞2, 王亚2, 黄乔乔2*
(1. 海南大学植物保护学院, 海口 570228; 2. 农业农村部热带作物有害生物综合治理重点实验室, 中国热带农业科学院环境与植物保护研究所, 海口 571101)
为了解环境对假高粱()种子萌发的影响,对海南岛假高粱种子的萌发特性进行了研究。结果表明,假高粱种子的萌发率较高,均在85%以上,在昼夜温度35℃/25℃下的萌发时间最短(2.64 d);浓硫酸处理可提高种子萌发率;浓硫酸处理、赤霉素浸泡和研磨均能缩短种子萌发时间;种子在土壤表层的萌发率最高,随着埋藏深度的增加,种子萌发率下降,萌发时间增加。灾变区与非灾变区种子在不同温度、打破休眠方式、埋藏深度的萌发率和萌发时间均无显著差异。可见,环境在一定程度上影响了假高粱种子的萌发,高温和土壤表层的种子有较高的萌发率和较快的萌发速度。海南岛的假高粱种子萌发率较高且萌发生态幅较宽,是近年来其在海南突发成灾的重要原因之一,因此,在假高粱花果期之前加强控制,减少种子的成熟和传播。
入侵杂草;假高粱;种子; 萌发;海南
近几十年来,外来生物入侵成为全球关注和研究的热点,入侵生物对许多地域的自然生态系统、生物多样性及人类健康造成了严重危害[1-3]。生物入侵对世界各国的农业生产和经济也造成了巨大损失,而我国每年因生物入侵遭受的经济损失位居世界第一[4]。在入侵中国的外来有害生物中,外来植物的入侵尤为人们所重视[5-6]。
外来植物的入侵能力与其繁殖性状密切相关[7]。许多强入侵性的外来植物兼具有性和无性繁殖能力、种子产量大、种子萌发率高且萌发速度快、种子可在多种环境条件下保持较高的萌发率等繁殖性状[2,7-9]。对美国佛罗里达4种入侵植物和4种非入侵植物的研究表明,相对于非入侵植物,入侵植物的种子产量更大[8]。对比研究中国的乌桕()种群和入侵到美国的乌桕种群, 入侵地种群种子显著大于原产地种群[10]。外来植物在入侵扩散过程中进化出了更快的种子萌发速度[11]。入侵中国的飞机草()比原产地的生长能力更强[12-13]。入侵中国的大狼耙草()中央瘦果芒刺显著比外围的更长,更易附着在载体上快速占据新生境[14]。由于入侵植物常在干扰严重的生境成为优势种[15],较高的种子产量和种子萌发率、较快的种子萌发速度、较强的生态适应性可能使入侵植物迅速占据干扰造成的空缺生境。
假高粱(),又名石茅,隶属于禾本科(Gramineae)高粱属,原产于欧洲地中海地区,是一种危害大、繁殖快、难防除的多年生杂草,是世界农业地区最危险的10大恶性杂草之一[16], 也是我国禁止输入的检疫性有害生物[17]。前人对假高粱的生长习性[18]、竞争能力[19-20]、最佳防除时期[21]、相关除草剂的控草效果和作用机制[22-23]等方面进行了研究。魏守辉等[24]对江苏省南京市假高粱的种子萌发特性进行了研究,认为较高的温度和较低的埋藏深度可促进假高粱种子萌发。
1921年首次在海南采到假高粱标本(http://www. cvh.ac.cn/)。根据海南植物志的记载,1977年假高粱已遍布海南各地。假高粱在海南存在已有近100年,之前未见报道其对农业生产造成严重危害,但近几年尤其在南繁核心区的三亚、陵水、乐东等地开始暴发成灾,在弃/休耕地、路边、荒地等生境形成高密度的单优种群,并大面积入侵玉米()、高粱()、甘蔗()等作物田,引起海南省南繁管理局的高度重视并进行了大规模的灭除控制。假高粱入侵多年,为何在近期才开始在海南部分地区暴发成灾?此外,调查还发现假高粱在海南岛中部、东北部等地仍是零星发生并不造成严重危害。本研究比较了灾变和非灾变区的假高粱在种子萌发性状方面的差异,揭示了海南岛假高粱种子的萌发特性,为分析其在海南岛部分地区暴发成灾的机制提供科学依据。
1 材料和方法
1.1 材料
假高粱()种子于2019年5月在海南岛内11个地点采集(图1), 种子采集的生境均为弃/休耕地或路边。根据我们前期的调查,按照假高粱目前的成灾情况,将假高粱入侵区域分为南部灾变区、北部灾变区和非灾变区。假高粱灾变区样点的判定标准为发生危害面积大于0.333 hm2,覆盖度大于70%;非灾变区样点的判定标准是发生面积小于0.067 hm2,覆盖度小于50%。南部灾变区4个采样点分别为三亚、陵水、乐东和万宁;北部灾变区4个采样点分别为福山、金江、加来和澄迈;非灾变区3个采样点分别为儋州、雅星和海口。多次从儋州、琼中至定安、海口、文昌的调查均未见较大的假高粱种群,因此,非灾变区不是因为人工控制而成。
每个地点采集的假高粱种子均在室温下贮存,保持干燥通风。选择大小均一、颗粒饱满的黑褐色种子进行试验,以100粒为单位称量,重复3次, 取平均值,记录单粒种子质量。
1.2 方法
采用培养皿滤纸法[25]。所有的种子均用次氯酸钠消毒并用去离子水反复清洗。去除颖片的假高粱种子先用98%浓硫酸处理4 min,然后放置于直径90 mm、铺设双层滤纸的培养皿中,每皿20粒种子,加入5 mL去离子水,培养皿置于气候箱中。每隔24 h观察记录种子萌发情况,并随机移动培养皿位置。种子萌发以胚根伸出2 mm为标准,连续5 d无变化则试验结束。每处理重复4次。
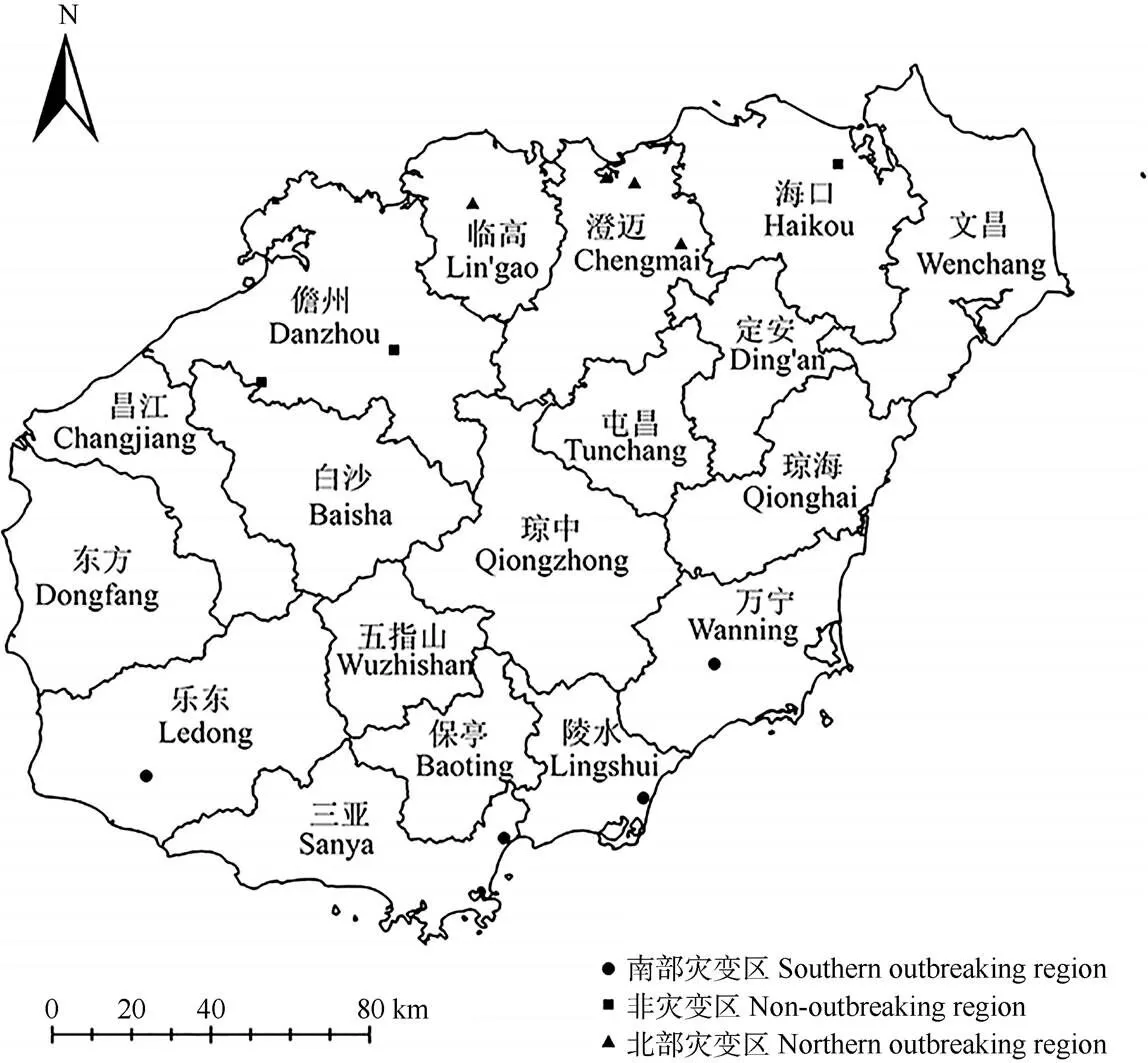
图1 海南岛内假高粱的采样点
温度 根据儋州气温记录(http://www.tianqi houbao.com/lishi/),气候箱的昼夜温差设置10℃, 气候箱昼夜温度分别设定为20℃/10℃、25℃/15℃、30℃/20℃、35℃/25℃,12 h/12 h (光/暗)。
打破休眠方式 分别用研磨、98%浓硫酸处理4 min、55℃水浴加热120 min、1.01 mmol/L赤霉素浸泡24 h等4种打破休眠方式处理假高粱种子,设置空白对照。种子处理后置于昼夜温度为35℃/ 25℃、12 h/12 h (光/暗)的气候箱中培养。
埋藏深度 去除颖片的假高粱种子先用98%浓硫酸处理4 min,放置于育苗盘的孔穴中,孔穴直径为7 cm、高为10 cm,土壤为壤土和营养土按1∶1的体积比混合(壤土从儋州耕地收集)。每个孔穴20粒种子,埋藏深度分别为0、2、4、6 cm。育苗盘放在昼夜温度为35℃/25℃、12 h/12 h (光/暗)的气候箱中培养。
1.3 数据的统计分析
萌发率(%)=(萌发种子数/种子总数)×100%; 萌发时间(d)=每颗种子萌发所需天数之和/萌发种子总数。
灾变区之间种子质量的差异用单因素方差分析(One-Way ANOVA)。温度、打破休眠处理方式、埋藏深度对萌发率和萌发时间的影响采用双因素嵌套方差分析,其中温度、打破休眠处理方式、埋藏深度和灾变区(南部灾变区、北部灾变区、非灾变区)为固定因子,采样点嵌套于灾变区类型为随机因子。采用SPSS 19.0 (SPSS Inc., Chicago, IL, USA)对数据进行分析,采用最小显著差异法(LSD)进行多重比较。
2 结果和分析
2.1 种子质量
南部灾变区、北部灾变区和非灾变区假高粱种子的平均质量分别为0.247、0.290和0.276 g。不同地区间的种子质量差异显著(=39.929,<0.001), 南部灾变区的最小,北部灾变区的最大(图2)。
2.2 温度对种子萌发的影响
不同温度下假高粱种子的萌发率普遍较高,均在85%以上(表1, 图3)。灾变区和非灾变区种子的萌发率和萌发时间没有显著差异。温度对种子萌发率的影响显著,以20℃/10℃处理的萌发率最低。
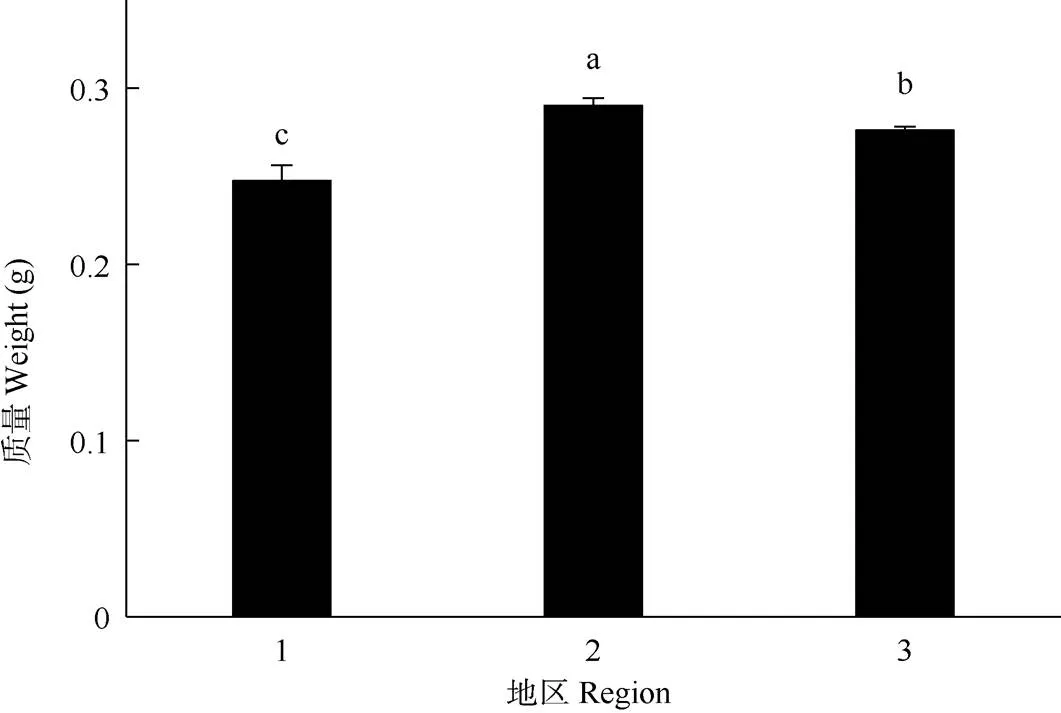
图2 种子质量。1: 南部灾变区; 2: 北部灾变区; 3: 非灾变区。柱上不同字母表示差异显著(P<0.05) (LSD检验)。下图同。
温度升高显著缩短假高粱种子萌发所需时间。在20℃/10℃下,种子萌发所需时间较长(7.3 d)。随着温度的升高,所需时间逐渐缩短,在35℃/25℃下萌发时间最短(2.64 d)。
2.3 打破休眠方式对种子萌发的影响
不同打破休眠方式对假高粱种子萌发率和萌发速度影响显著,灾变区和非灾变区的种子萌发率和萌发时间没有显著差异(表2, 图4)。浓硫酸处理的种子萌发率最高(96.23%),高于对照(81.53%)。研磨处理可能会破坏种子种皮,种子易发霉,萌发率较低(71.87%)。浓硫酸处理和研磨后的假高粱种子萌发平均所需时间分别为3.09和4.23 d,均低于对照(8.28 d),表明浓硫酸处理和研磨均能有效打破假高粱种子的休眠,缩短种子萌发时间。
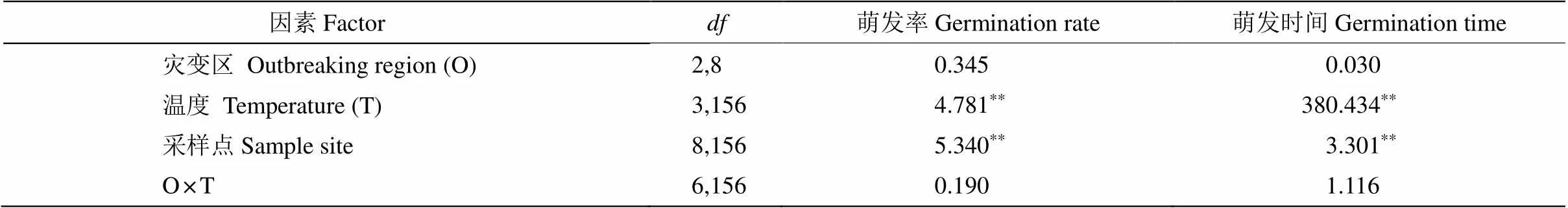
表1 灾变区、温度及其交互作用对假高粱种子萌发的影响(F值)
:0.05;:0.01。下表同。
:0.05;:0.01. The same is following Tables.
水浴加热处理后种子的萌发率为83.4%、萌发时间为7.74 d,均与对照相近,表明水浴加热对打破假高粱种子休眠的效果并不明显。赤霉素浸泡的种子萌发率为78.96%,与对照相近, 萌发时间为6.13 d,低于对照,表明赤霉素处理具有加快假高粱种子萌发的作用。
2.4 埋藏深度对种子萌发的影响
不同埋藏深度对假高粱种子的萌发率和萌发速度影响显著(表3, 图5)。种子在土壤表层的萌发率最高(83.93%),埋藏深度为2、4、6 cm时,萌发率大约为45%。种子萌发时间随着埋藏深度增加而增加,埋藏深度为6 cm的萌发时间显著大于其他埋藏深度。
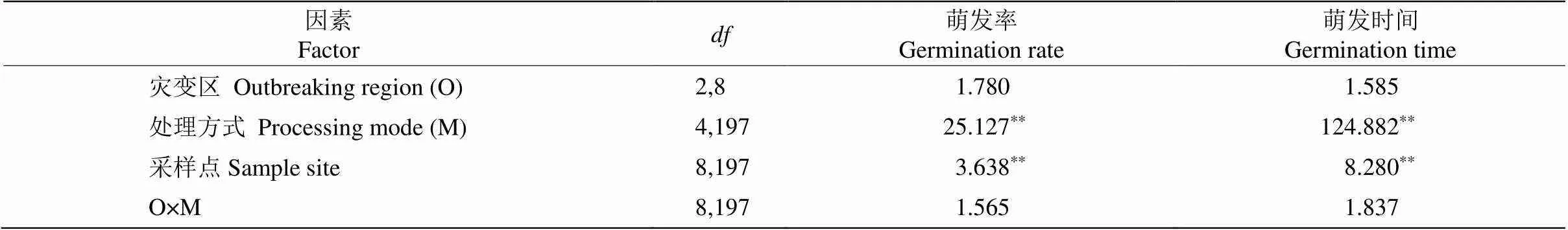
表2 灾变区、打破休眠处理方式及其交互作用对假高粱种子萌发的影响(F值)
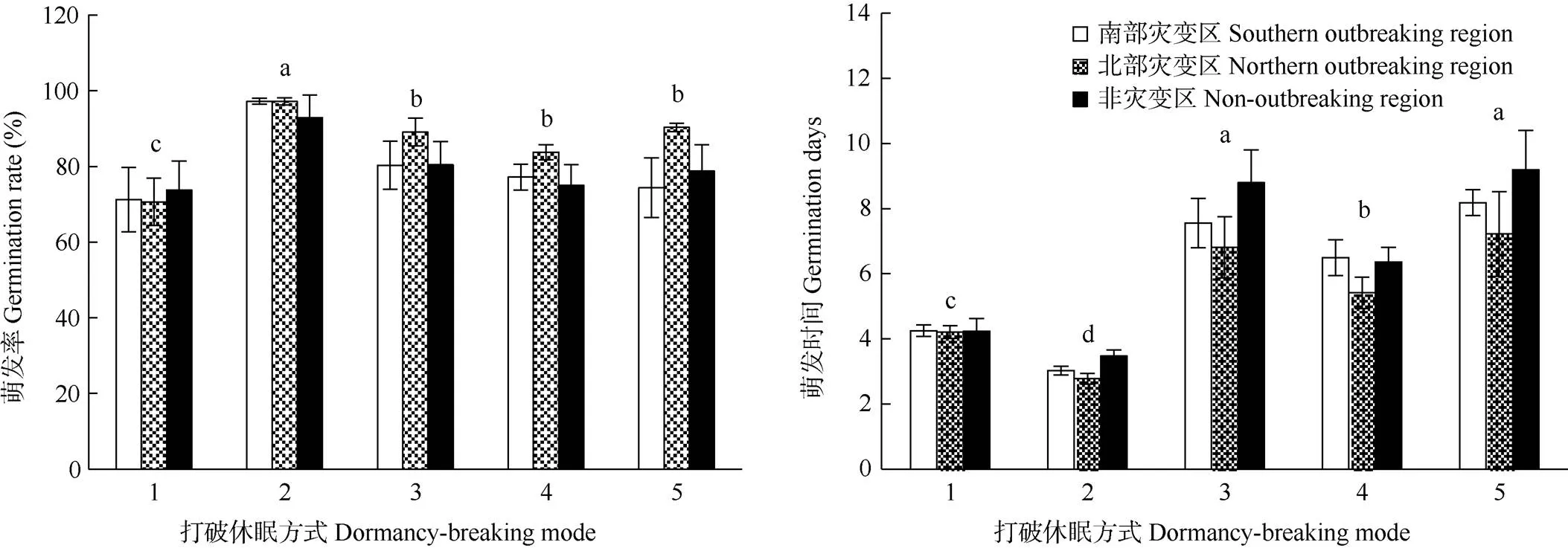
图4 打破休眠方式对假高粱种子萌发率和萌发时间的影响。1: 研磨; 2: 浓硫酸; 3: 水浴; 4: 赤霉素; 5: 对照。
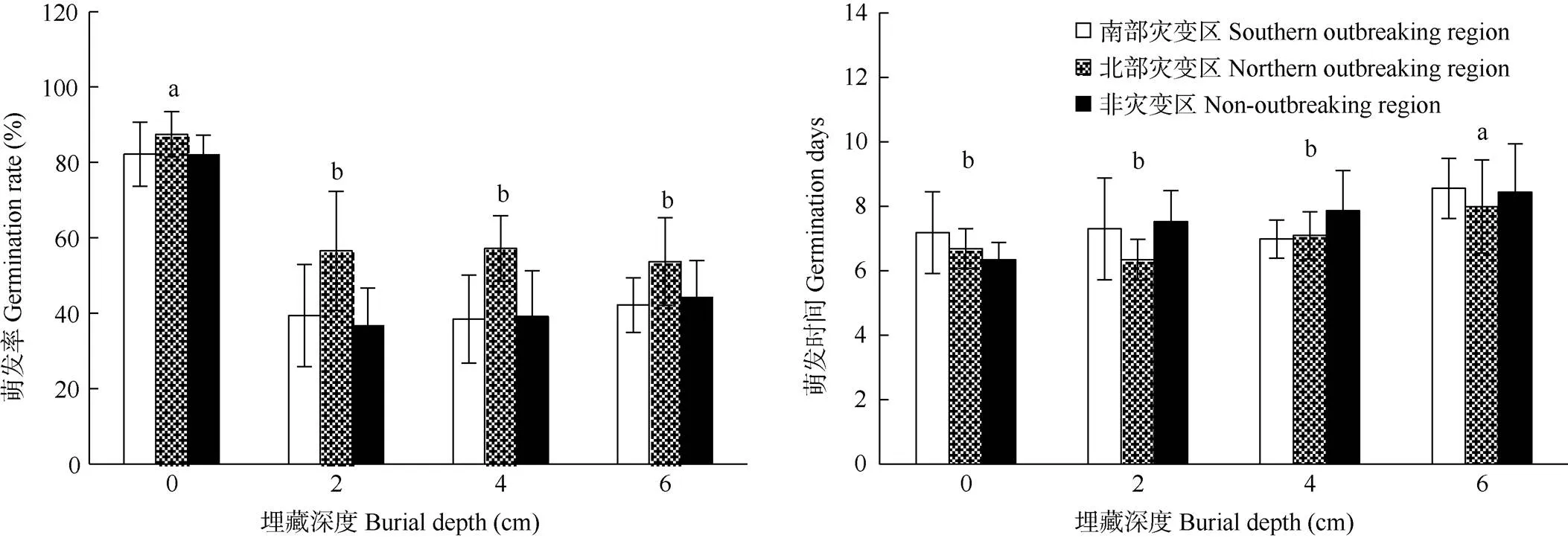
图5 埋藏深度对假高粱种子萌发率和萌发时间的影响
3 结论和讨论
在本研究中,南部灾变区假高粱的种子质量小于北部灾变区和非灾变区的。种子质量大可能有利于萌发、幼苗存活和竞争[26-29],而小种子可能更有利于扩散且数量多[30]。环境条件,如种群密度[31]、传粉环境[32]等也可能影响种子大小。假高粱种子质量可能是对当地环境适应的结果,南繁核心区的土地常在冬季被利用而在其他季节被搁置,因而南部灾变区假高粱种群可能受到频繁的干扰,较小和较多的种子利于其迅速占据干扰后的生境。
本研究结果表明,各采样点的假高粱种子萌发率普遍较高,温度、打破休眠方式、埋藏深度均可影响种子的萌发率和萌发速度,这与魏守辉等[24]对江苏省南京市假高粱种子的研究结果存在异同点。相同点是两省的假高粱种子在35℃/25℃下能较好萌发,最适萌发温度约为30℃;土壤埋藏对假高粱种子萌发的抑制作用明显,置于土壤表层的种子萌发率最高,随着埋藏深度的增加,种子萌发率逐渐降低。不同之处是本研究结果表明,假高粱种子在不同温度下的萌发率均较高(85%以上),在相对较低的温度(20℃/10℃)也有较高的萌发率;而南京的假高粱种子仅在30℃恒温或35℃/25℃下萌发率可达到80%左右,在其他温度下(25℃和35℃)萌发率较低,说明海南省的假高粱种子可在更广的温度范围内保持较高的萌发率。此外,本研究结果表明,不经任何处理(对照)的种子萌发率也高达80%左右,明显高于之前的研究(24.44%),这说明海南省的假高粱种子颖片对其萌发率的抑制作用不明显。在土壤表层的假高粱种子萌发率为83.93%,比魏守辉等[24]报道的50% (萌发温度约为27℃)高出很多,说明海南省的假高粱有性繁殖能力更强,种子萌发率较高且有较宽的萌发生态幅,这应该是近年来假高粱在海南突发成灾的重要原因之一。因此,需在假高粱花果期前加强对其的控制,避免具有较高萌发率和较宽萌发生态幅的种子的成熟和传播。
各处理下灾变区与非灾变区假高粱种子的萌发率和萌发时间均没有显著差异,可能是海南不同地区的环境条件、土地利用和管理方式的差异造成了假高粱仅在部分地区暴发成灾。南繁区(南部灾变区)除冬季外,大量土地闲置,可能促进了假高粱暴发成灾。因此,需加强对闲置土地的管理,避免其成为假高粱重点入侵的生境并以此向外扩张。尽管不同灾变区间种子萌发率没有显著差异,但从埋藏深度的影响来看,南部灾变和非灾变区(集中分布于北部)种子的萌发率较为接近,均低于北部灾变区种子的萌发率,说明海南北部灾变区的假高粱可能正处于种群扩张阶段,并可能取代非灾变区的假高粱。虽然本研究表明灾变区与非灾变区之间假高粱的种子萌发率不存在显著差异,但我们还不能确定灾变区与非灾变区假高粱种群之间不存在遗传变异。灾变区和非灾变区的假高粱可能在其他方面有差异,如竞争能力、表型可塑性等。因此,在未来的研究中,比较灾变区和非灾变区假高粱在竞争能力、对环境条件变异的响应等的差异,进一步揭示假高粱成灾的机制。
[1] MACK R N, SIMBERLOFF D, LONSDALE W M, et al. Biotic invasions: Causes, epidemiology, global consequences, and control [J]. Ecol Appl, 2000, 10(3): 689-710. doi: 10.1890/1051-0761(2000)010 [0689:BICEGC]2.0.CO;2.
[2] HUANG Q Q, WU J M, BAI Y Y, et al. Identifying the most noxious invasive plants in China: Role of geographical origin, life form and means of introduction [J]. Biodiv Conserv, 2009, 18(2): 305-316. doi: 10.1007/s10531-008-9485-2.
[3] VAN KLEUNEN M, DAWSON W, ESSL F, et al. Global exchange and accumulation of non-native plants [J]. Nature, 2015, 525(7567): 100- 103. doi: 10.1038/nature14910.
[4] PAINI D R, SHEPPARD A W, COOK D C, et al. Global threat to agriculture from invasive species [J]. Proc Natl Acad Sci USA, 2016, 113(27): 7575-7579. doi: 10.1073/pnas.1602205113.
[5] LI Q, CHEN Q, HE F R, et al. Arbuscular mycorrhizal fungi promote the growth ofand the absorption of insoluble phosphorus [J]. J Trop Subtrop Bot, 2020, 28(4): 339-346. doi: 10. 11926/jtsb.4170. 李琴, 陈琪, 贺芙蓉, 等. 丛枝菌根真菌促进南美蟛蜞菊生长及对难溶磷的吸收 [J]. 热带亚热带植物学报, 2020, 28(4): 339-346. doi: 10.11926/jtsb.4170.
[6] HUANG Q Q, SHEN Y D, LI X X, et al. Research progress on the distribution and invasiveness of alien invasive plants in China [J]. Ecol Environ Sci, 2012, 21(5): 977-985. doi: 10.3969/j.issn.1674-5906.2012. 05.032. 黄乔乔, 沈亦德, 李晓霞, 等. 外来入侵植物在中国的分布及入侵能力研究进展 [J]. 生态环境学报, 2012, 21(5): 977-985. doi: 10. 3969/j.issn.1674-5906.2012.05.032.
[7] VAN KLEUNEN M, WEBER E, FISCHER M. A meta-analysis of trait differences between invasive and non-invasive plant species [J]. Ecol Lett, 2010, 13(2): 235-245. doi: 10.1111/j.1461-0248.2009.01418.x.
[8] BURNS J H. Demographic performance predicts invasiveness of species in the Commelinaceae under high-nutrient conditions [J]. Ecol Appl, 2008, 18(2): 335-346. doi: 10.1890/07-0568.1.
[9] RUPRECHT E, FENESI A, NIJS I. Are plasticity in functional traits and constancy in performance traits linked with invasiveness? An experimental test comparing invasive and naturalized plant species [J]. Biol Invas, 2014, 16(7): 1359-1372. doi: 10.1007/s10530-013- 0574-0.
[10] ZHANG J L, SIEMANN E, TIAN B L, et al. Differences in seed properties and germination between native and introduced populations of[J]. J Plant Ecol, 2020, 13(1): 70-77. doi: 10. 1093/jpe/rtz048.
[11] TABASSUM S, LEISHMAN M R. Have your cake and eat it too: Greater dispersal ability and faster germination towards range edges of an invasive plant species in eastern Australia [J]. Biol Invas, 2018, 20 (5): 1199-1210. doi: 10.1007/s10530-017-1620-0.
[12] QIN R M, ZHENG Y L, VALIENTE-BANUET A, et al. The evolution of increased competitive ability, innate competitive advantages, and novel biochemical weapons act in concert for a tropical invader [J]. New Phytol, 2013, 197(3): 979-988. doi: 10.1111/nph.12071.
[13] ZHENG Y L, FENG Y L, ZHANG L K, et al. Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader [J]. New Phytol, 2015, 205(3): 1350-1359. doi: 10.1111/nph.13135.
[14] ZHOU C Q, TANG S C, PAN Y M, et al. Effects of light and tem- perature on germination of heteromorphic achenes ofL. [J]. J Trop Subtrop Bot, 2015, 23(6): 662-668. doi: 10.11926/j.issn. 1005-3395.2015.06.010. 周超群, 唐赛春, 潘玉梅, 等. 光照和温度对入侵植物大狼耙草异型瘦果萌发的影响 [J]. 热带亚热带植物学报, 2015, 23(6): 662-668. doi: 10. 11926/j.issn.1005-3395.2015.06.010.
[15] DAVIS M A, GRIME J P, THOMPSON K. Fluctuating resources in plant communities: A general theory of invasibility [J]. J Ecol, 2000, 88(3): 528-534. doi: 10.1046/j.1365-2745.2000.00473.x.
[16] MONAGHAN N. The biology of Johnson grass () [J]. Weed Res, 1979, 19(4): 261-267. doi: 10.1111/j.1365-3180.1979. tb01536.x.
[17] ZHANG Q, LIN J C, QIANG S. Analysis of the importance of quarantining based on the port checking out ratio for Johnson grass [J]. J Anhui Agric Sci, 2004, 32(3): 448-451. doi: 10.13989/j.cnki.0517- 6611.2004.03.028. 张强, 林金成, 强胜. 检疫口岸假高粱检出率分析及其防治 [J]. 安徽农业科学, 2004, 32(3): 448-451. doi: 10.13989/j.cnki.0517-6611. 2004.03.028.
[18] ROSALES-ROBLES E, CHANDLER J M, WU H I, et al. A model to predict the influence of temperature on rhizome Johnson grass () development [J]. Weed Sci, 2003, 51(3): 356-362. doi: 10.1614/0043-1745(2003)051[0356:AMTPTI]2.0.CO;2.
[19] WOOD M L, MURRAY D S, BANKS J C, et al. Johnson grass () density effects on cotton () harvest and economic value [J]. Weed Technol, 2002, 16(3): 495-501. doi: 10.1614/0890-037X(2002)016[0495:JSHDEO]2.0.CO;2.
[20] MITSKAS M B, TSOLIS C E, ELEFTHEROHORINOS I G, et al. Interference between corn and Johnsongrass () from seed or rhizomes [J]. Weed Sci, 2003, 51(4): 540-545. doi: 10.1614/ 0043-1745(2003)051[0540:IBCAJS]2.0.CO;2.
[21] GHOSHEH H Z, HOLSHOUSER D L, CHANDLER J M. The critical period of Johnson grass () control in field corn () [J]. Weed Sci, 1996, 44(4): 944-947. doi: 10.1017/S00431745 00094960.
[22] HUANG S G, CAI R J, ZHONG X S, et al. Tests of several kinds of herbicides for controlling(L.) [J]. Plant Quarantine, 2002, 16(6): 341-344. doi: 10.19662/j.cnki.issn1005-2755.2002.06.008. 黄胜光, 蔡荣金, 钟学伸, 等. 几种除草剂防除假高粱药效试验 [J]. 植物检疫, 2002, 16(6): 341-344. doi: 10.19662/j.cnki.issn1005-2755. 2002.06.008.
[23] FERRELL J A, EARL H J, VENCILL W K. The effect of selected herbicides on CO2assimilation, chlorophyll fluorescence, and stomatal conductance in Johnson grass (L.) [J]. Weed Sci, 2003, 51(1): 28-31. doi: 10.1614/0043-1745(2003)051[0028:TEOSHO] 2.0.CO;2.
[24] ZHENG L, FENG Y L. Allelopathic effects ofSpreng. on seed germination and seedling growth in ten herbaceous species [J]. Acta Ecol Sinica, 2005, 25(10): 2782-2787. doi: 10.3321/j.issn:1000-0933.2005.10.046. 郑丽, 冯玉龙. 紫茎泽兰叶片化感作用对10种草本植物种子萌发和幼苗生长的影响 [J]. 生态学报, 2005, 25(10): 2782-2787. doi: 10. 3321/j.issn:1000-0933.2005.10.046.
[25] WEI S H, ZHANG C X, LI C H, et al. Seed germination behavior of Johnson grass () [J]. Sci Agric Sin, 2008, 41(1): 116-121. doi: 10.3864/j.issn.0578-1752.2008.01.015. 魏守辉, 张朝贤, 黎春花, 等. 外来恶性杂草假高粱种子萌发特性研究 [J]. 中国农业科学, 2008, 41(1): 116-121. doi: 10.3864/j.issn. 0578-1752.2008.01.015.
[26] LARIOS E, BÚRQUEZ A, BECERRA J X, et al. Natural selection on seed size through the life cycle of a desert annual plant [J]. Ecology, 2014, 95(11): 3213-3220. doi: 10.1890/13-1965.1.
[27] GUO Y Y, LIU H J, YU H Y, et al. Effects of seed size and drought stress on growth ofseedlings [J]. J NW Agric For Univ (Nat Sci), 2016, 44(8): 143-147. doi: 10.13207/j.cnki.jnwafu. 2016.08.021. 郭有燕, 刘宏军, 余宏远, 等. 种子大小和干旱胁迫对文冠果幼苗生长的影响 [J]. 西北农林科技大学学报(自然科学版), 2016, 44(8): 143-147. doi: 10.13207/j.cnki.jnwafu.2016.08.021.
[28] TREMAYNE M A, RICHARDS A J. Seed weight and seed number affect subsequent fitness in outcrossing and selfingspecies [J]. New Phytol, 2000, 148(1): 127-142. doi: 10.1046/j.1469-8137.2000. 00738.x.
[29] MAO P L, GUO L M, GAO Y X, et al. Effects of seed size and sand burial on germination and early growth of seedlings for coastalParl. in the northern Shandong Peninsula, China [J]. Forests, 2019, 10(3): 281. doi: 10.3390/f10030281.
[30] HAMILTON M A, MURRAY B R, CADOTTE M W, et al. Life- history correlates of plant invasiveness at regional and continental scales [J]. Ecol Lett, 2005, 8(10): 1066-1074. doi: 10.1111/j.1461-0248. 2005.00809.x.
[31] HUANG F F, PENG S L, CHEN B M, et al. Rapid evolution of dispersal-related traits during range expansion of an invasive vine[J]. Oikos, 2015, 124(8): 1023-1030. doi: 10.1111/ oik.01820.
[32] HUANG Q Q, BURD M, FAN Z W. Resource allocation and seed size selection in perennial plants under pollen limitation [J]. Amer Nat, 2017, 190(3): 430-441. doi: 10.1086/692543.
Seed Germination Traits of Invasive Weedin Hainan Island
WANG Wei-qian1,2, LI Xiao-xia2, WANG Ya2, HUANG Qiao-qiao2*
(1. College of Plant Protection, Hainan University,Haikou 570228, China; 2. Key Laboratory of Integrated Pest Management on Tropical Crops, Ministry of Agriculture and Rural Affairs, Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences,Haikou 571101, China)
In order to reveal the effect of environment condition on seed germination of, its germination characters were studied in Hainan Island. The results showed that the germination rate ofseeds was high with more than 85%, and the germination time was the shortest (2.64 d) under day and night temperature at 35℃/25℃. The seeds treated with 98% concentrated sulfuric acid could improve germination rate. The seed germination time was shortened treated with concentrated sulfuric acid, gibberellin and grinding. The seeds on the soil surface had the highest germination rate, and germination rate decreased and germination time increased with burial depth. Among different treatments, such as temperature, dormancy breaking and burial depth, there were no significant differences in germination rate and germination time ofseeds between outbreaking and non-outbreaking regions. Therefore, environmental conditions could influence germination ofseeds, and the seeds under high temperature and on the soil surface had high germination rate and short germination time. Seeds ofin Hainan Island had high germination rate and wide ecological breadth of germination, which was one of important reasons for its sudden disaster in Hainan in recent years. So, the strengthen control before flowering of, reduce seed maturation and dissemination.
Invasive weed;; Seed; Germination; Hainan
10.11926/jtsb.4302
2020-09-08
2020-10-16
海南省基础与应用基础研究计划(自然科学领域)高层次人才项目(2019RC280); 海南省重点研发计划项目(ZDYF2019055); 中国热带农业科学院基本科研业务费专项(1630042019014)资助
This work was supported by the Project for High-level Talents of Basic and Applied Basic Research (Natural Science) in Hainan (2019RC280), the Project for Key Research and Development in Hainan (Grant No. ZDYF2019055), and the Project for Central Public-interest Scientific Institution Basal Research in Chinese Academy of Tropical Agricultural Sciences (Grant No. 1630042019014).
王玮倩(1994~ ), 女, 在读硕士, 主要从事植物入侵生态学研究。E-mail: 1310744381@qq.com
Corresponding author. E-mail: huangqq@catas.cn
