氮素形态对铁线莲光合特性及氮代谢的影响
2021-06-03马晓华胡青荻章彦君钱仁卷郑坚刘洪见
马晓华, 胡青荻, 章彦君, 钱仁卷, 郑坚, 刘洪见
氮素形态对铁线莲光合特性及氮代谢的影响
马晓华1*, 胡青荻1, 章彦君2, 钱仁卷1, 郑坚1*, 刘洪见1
(1. 浙江省亚热带作物研究所,浙江 温州 325005; 2. 国家林业和草原局竹子研究中心,杭州 310000)
为了解铁线莲的光合性能和氮素代谢的响应机制,对不同氮素形态配比下1 a生厚叶铁线莲()与天台铁线莲(ssp.)的生长、光响应曲线、A-Ci曲线和氮代谢相关酶活性进行了比较。结果表明,氮素形态配比显著影响铁线莲的生物量和叶绿素(Chl)含量,厚叶铁线莲在铵态氮(NH4+-N)与硝态氮(NO3--N)配比为1∶1时,生物量、Chl a、Chl b、Car含量达到最大;天台铁线莲的Chl a和Chl b含量在NH4+-N/NO3--N为1∶3时最高。在NH4+-N/NO3--N为1∶1时,厚叶铁线莲的光饱和点和天台铁线莲的光补偿点为最大值,且厚叶铁线莲叶片的Vcmax与Jmax值均显著高于其他处理。天台铁线莲的谷氨酰胺合成酶(GS)活性在NH4+-N/NO3--N为3∶1处理下最高,而硝酸还原酶(NR)和亚硝酸还原酶(NiR)活性在NH4+-N/NO3--N为1∶3时显著高于其他处理。可见,NH4+-N和NO3--N混合施用能有效促进2种铁线莲的生长和光合作用,加强氮素利用效率。厚叶铁线莲适宜的NH4+-N/NO3--N配比为1∶1,而天台铁线莲更适宜1∶3。
铁线莲;氮素形态;生物量;光合特性;氮代谢
植物的生长、发育代谢需要各种各样的营养, 而土壤不仅是植物生长的介质,也是植物发育和产出所需养分的主要供给者。肥料是改善土壤理化性质的重要物质,对提高土壤肥力,加强植物的营养供给,提高植物品质及增强植物抗逆性具有十分重要的作用[1]。氮素是限制植物生命活动的主要元素之一,是参与生物合成氨基酸、核酸、蛋白酶及其它含氮有机物的大量营养元素之一,在植物的生长发育中发挥极其重要的作用[2]。然而,植物对氮素的吸收利用不仅取决于植物自身生长代谢特点,还取决于生长环境中的各种氮源[3]。一般说来,土壤中的氮含量往往不能满足植物生长发育所需,而氮肥是生产中重要的土壤氮素补充手段,因此施氮肥是促进植株生长发育、维持养分平衡、保证高产、稳产的重要措施之一[4]。硝态氮(NO3--N)和铵态氮(NH4+-N)是无机氮素的两种形态,其不同的配比会直接影响土壤微生物活性进而影响土壤的养分状况[5], NO3--N和NH4+-N也是植物吸收的主要氮素形态,因此,二者的配比一直是氮肥施肥技术研究的热点。
铁线莲属()隶属毛茛科(Ranunculaceae),多为多年生草质或木质藤本植物,种类繁多,花型多变,花色艳丽,花期长, 被誉为“藤本皇后”。我国铁线莲属植物资源十分丰富,约有155种[6],其中不乏观赏性状优异的种质资源,其中就包含几种具有代表性的常绿铁线莲种。厚叶铁线莲(),适应性广、抗逆性强被广大园艺工作者青睐;天台铁线莲(ssp.)为浙江本地特有种,花色白、叶片革质、适应性强。随着近些年来观赏园艺的发展,铁线莲属植物已经走进千家万户,更由于常绿铁线莲终年常绿,观赏期长,因而更受青睐,市场需求量逐年递增。由于铁线莲的种类繁多,且部分种间差异较大,因而在实际生产管理中的水肥需求也显著不同。相同施肥量可能导致部分铁线莲叶片小而发黄,生长发育迟缓;而另一部分却出现徒长、枝叶繁茂,却迟迟不开花,因而明确适宜施肥量和肥料种类是铁线莲生产中亟待解决的问题。
研究表明,铁线莲属不同植物的生长管理存在显著差异[7]。氮素形态不同配比,显著影响冬小麦()[8]、水稻()[9]、番茄()[10]等植物叶片的叶绿素含量、光合作用效率和氮素的转化利用。NH4+-N可诱导番茄根中的谷氨酸脱氢酶(GOH)活性,但在水稻中却不然[11];也有研究表明,水稻等供应NH4+-N更有利于生长[9],然而NO3--N对小麦生长的影响优于NH4+-N[8]。通常,植株根系吸收的NO3--N大部分需要经过还原才能被利用,小部分储存在液泡中作为离子平衡和渗透调节物质,因此,氮素的形态差异可直接影响植物的元素吸收利用、光合作用,进一步影响其营养积累和生长发育,最终决定植物的生长周期。本试验旨在通过施用不同形态氮素配比的肥料,对2种常绿铁线莲叶片的光响应、A-Ci曲线、叶绿素含量、叶氮含量及光合氮利用率等生化指标进行测定,探讨铁线莲种苗期的光合特性和氮素利用差异,揭示常绿铁线莲种的施肥管理及氮素利用机制,对铁线莲植物苗木的培育提供理论依据。
1 材料和方法
1.1 材料
试验地设在浙江温州景山生态容器育苗基地(28°23′ N, 120°72′ E),以1 a生长势一致、生长健壮的厚叶铁线莲()和天台铁线莲(ssp)为试验材料。厚叶铁线莲和天台铁线莲均为攀援藤本植物,单株种植于育苗盆内,盆中放置攀爬架,盆底放置托盘,防止水肥流失。育苗容器规格为4.5 cm×6.7 cm×9.0 cm,基质为60%泥炭、20%蛭石和20%稻壳体积比复配,基质的容重为0.23 g/cm3,pH为5.62,有机质含量为278.36 g/kg,速效氮、磷和钾含量分别为0.15、0.055和0.27 g/kg。试验过程中基质保持水分充足(达田间持水量的75%)。
1.2 方法
采用氮素形态比例与铁线莲种类双因素试验,在保持各处理供氮水平一致的前提下,每盆施总氮8.50 g。设5个水平的氮素形态比(NH4+-N∶NO3--N):1∶0、3∶1、1:1、1∶3和0∶1,其中铵态氮(NH4+- N)由硫酸铵[(NH4)2SO4]提供,硝态氮(NO3--N)由硝酸钠(NaNO3)提供。2019年5月,选择苗龄相同、生长健壮、长势一致的铁线莲盆栽,将配置的氮肥溶于水中并浇入基质,一共10个处理,每处理5个重复。试验配制的氮肥除氮素形态配比外,其他元素含量均相同,用硫酸纳平衡各处理间的Na+差异。试验前适量控水以利于肥料在盆土中的扩散。每次浇水时用清水清洗盆栽下的托盘,并将水倒入盆中,以防止基质中养分流失,试验处理45 d后进行叶绿素含量、光响应曲线、CO2响应曲线、氮代谢过程关键酶活性等测定。
试验结束后,选取完整植株,根和茎冲洗干净晾干,分别测定地下部分和地上部分生物量。比叶重采用熊庆娥等[12]的方法测量,采集同一位置长势相近的成熟叶片10枚,用蒸馏水冲洗并擦干,用叶面积仪测量叶面积,然后用烘箱烘干,称量干质量, 比叶重(mg/cm2)=叶片干质量/叶面积。
选择完整、长势健康的植株,用Li-6400便携式光合仪(美国Li-cor公司)的6400-22不透明簇状叶室测定铁线莲叶片的光响应和A-Ci曲线。光响应曲线测定时,设置光合有效辐射强度(PAR)梯度为1 600、1 200、1 000、800、600、400、200、150、100、50和0mol/(m2·s)。测定前用1 000mol/(m2·s)的光照强度进行光诱导。A-Ci曲线测定时,光照强度为1 000mol/(m2·s),在0、50、100、150、200、400、600、800、1 200、1 500和1 800mol/mol CO2浓度下测定净光合速率(Pn)[13]。
采集当年生枝条距顶端3~4片生长成熟、健康的叶片,用于叶绿素含量测定[14],用紫外分光光度计(UV2500,岛津)在波长665、649和470 nm处测吸光度;参考乔富廉[15]的方法测定叶片的谷氨酰胺合成酶(GS)活性;谷氨酸脱氢酶(GDH)采用Loulakakis等[16]的方法测定;谷氨酸合成酶(GOGAT)活性参考马振勇等[17]的方法测定;叶片的硝酸还原酶(NR)活性采用体内法进行测定[14];参照孙世芹等[18]的方法测定亚硝酸还原酶(NiR)活性,用g NO2-/(mg FW·h)表示;用紫外分光法测定铵态氮含量与硝态氮含量[19]。
1.3 数据分析
采用SPSS version 13.0 (SPSS Inc.,Standard Version)进行数据的统计分析,采用LSD多重比较分析处理间差异的显著,设定置信度为5%。
2 结果和分析
2.1 对铁线莲生长的影响
如图1所示,不同氮素形态配比显著影响铁线莲的生物量(<0.05)。厚叶铁线莲在铵态氮与硝态氮配比(NH4+-N∶NO3--N)为1∶1时生物量最高, 而天台铁线莲在配比为3∶1时最高(图1: A)。比叶重是衡量植物光合作用性能的一个重要参数,氮素形态配比也显著影响铁线莲的比叶重,厚叶铁线莲在配比为3∶1时比叶重最高;天台铁线莲则在1∶3时最大。
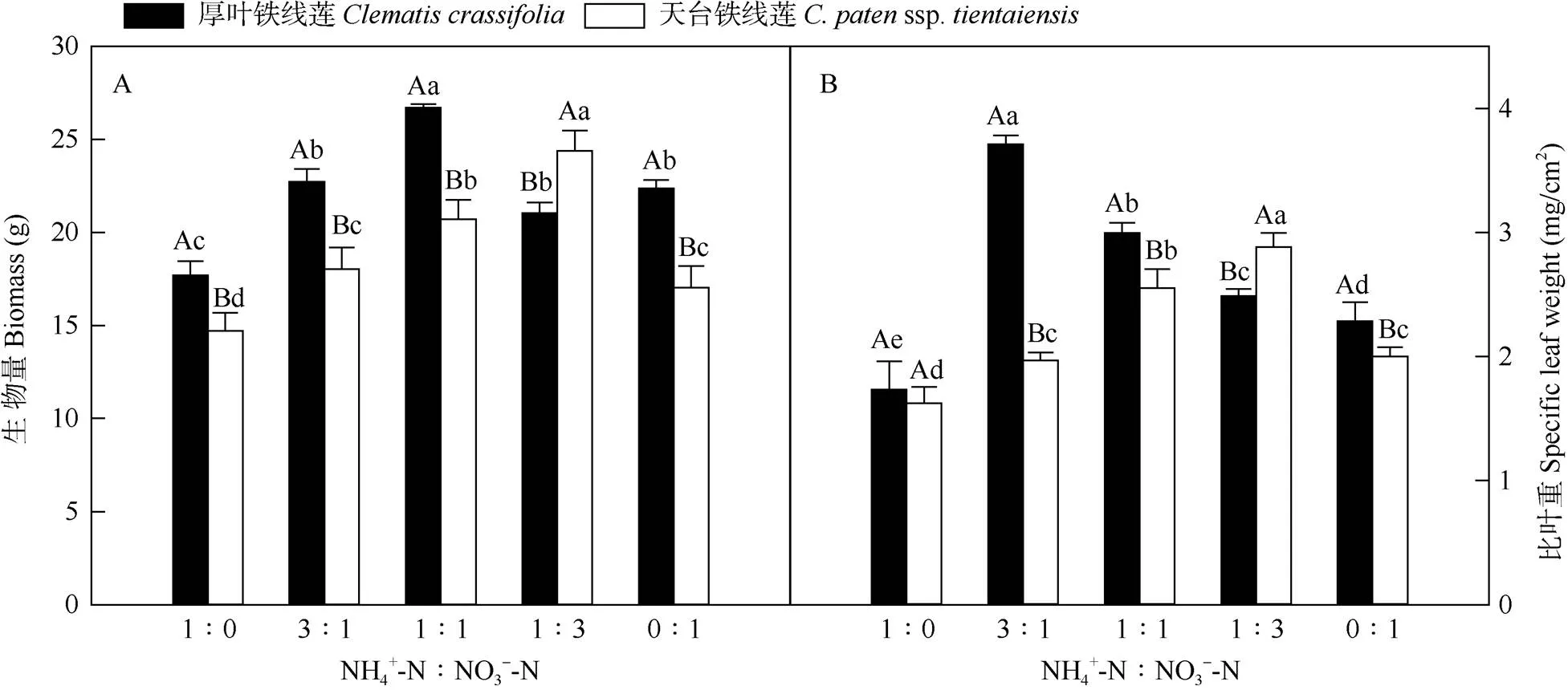
图1 厚叶铁线莲和天台铁线莲的生物量(A)和比叶重(B)变化。柱上不同大写和小写字母分别表示相同氮素配比下不同铁线莲间和同种铁线莲在不同氮素配比下差异显著(P<0.05)。下图同。
2.2 对叶片叶绿素含量的影响
不同氮素形态配比显著影响铁线莲叶片的叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量(图2)。在NH4+-N∶NO3--N配比为1∶1时, 厚叶铁线莲的Chl a、Chl b和Car含量最大;而天台铁线莲的Chl a和Chl b含量在1∶3时最大。天台铁线莲的Chl a/b在配比为1∶1时显著高于其他处理,厚叶铁线莲在配比为1∶3时显著高于其他处理,其余处理间的Chl a/b无显著差异。

图2 厚叶铁线莲和天台铁线莲叶片的叶绿素a (A)、叶绿素b (B)、类胡萝卜素(C)含量和叶绿素a/b (D)的变化
2.3 对光响应曲线的影响
不同氮素形态配比处理下,厚叶铁线莲与天台铁线莲叶片Pn的变化趋势基本一致。PAR为0~ 800mol/(m2·s)时,叶片Pn随着PAR的升高快速上升; 当PAR>800mol/(m2·s)时逐渐趋于缓慢,最后达到光饱和(图3)。不同氮素形态配比下, 2种铁线莲的光饱和点与光补偿点发生显著变化, 其中厚叶铁线莲的光饱和点在配比为1∶1时最高, 而光补偿点在1∶3时最高;天台铁线莲的光饱和点出现在3∶1时,而光补偿点最高值出现在1∶1处理条件下。厚叶铁线莲与天台铁线莲叶片的Pmax分别在配比为0∶1与1∶3时最大(表1)。
2.4 对A-Ci曲线的影响
在不同氮素形态配比下铁线莲叶片的Pn随Ci升高的变化趋势基本一致。Ci≤1 000mol/mol时, Pn随Ci的升高而快速上升(图4)。从表2可见,2种铁线莲的CO2补偿点在不同氮素形态配比下差异显著,但CO2补偿点均在配比为1∶0时最高,在配比为0∶1时最低。在配比为1∶1时,厚叶铁线莲叶片的Vcmax与Jmax均显著高于其他处理,而天台铁线莲的Vcmax在配比为1∶0时显著低于其他处理,且其他处理间差异不显著;Jmax在配比为1∶3和0∶1时显著高于其他处理。
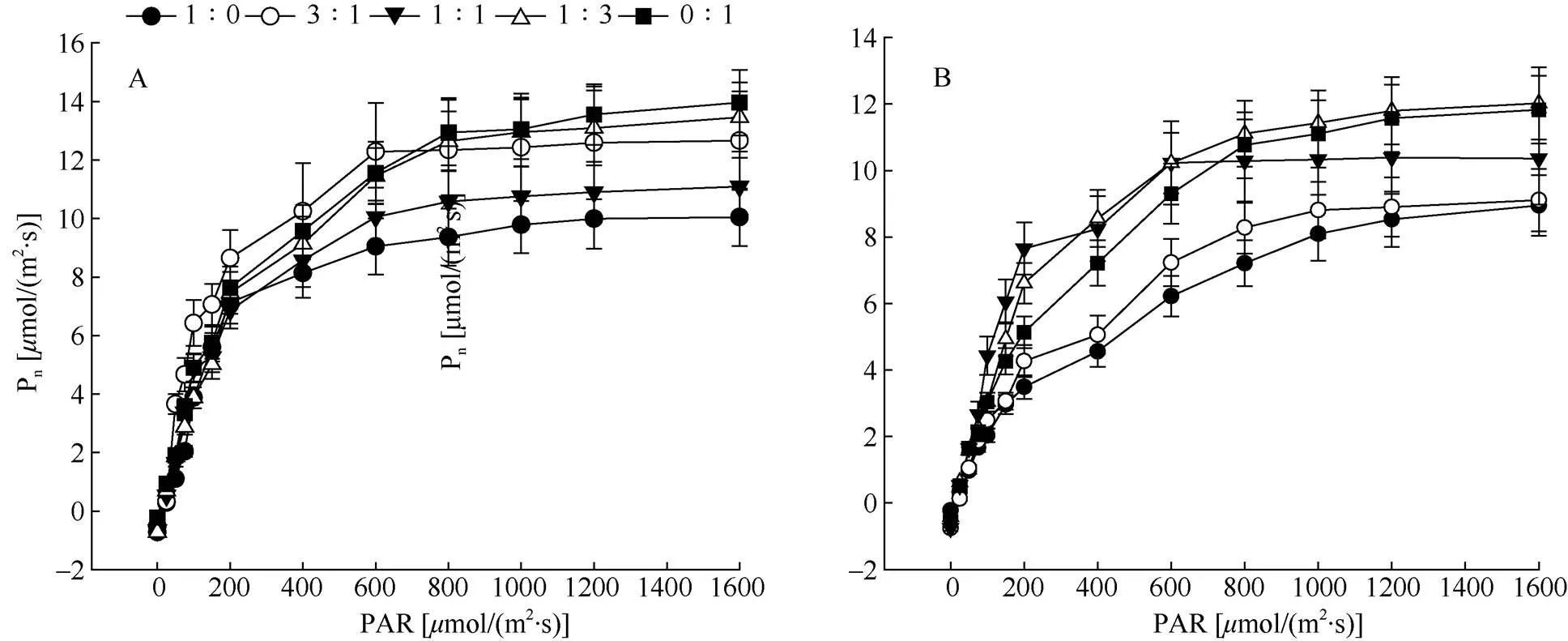
图3 厚叶铁线莲(A)和天台铁线莲(B)的光响应曲线。Pn: 净光合速率; PAR: 光合有效辐射。
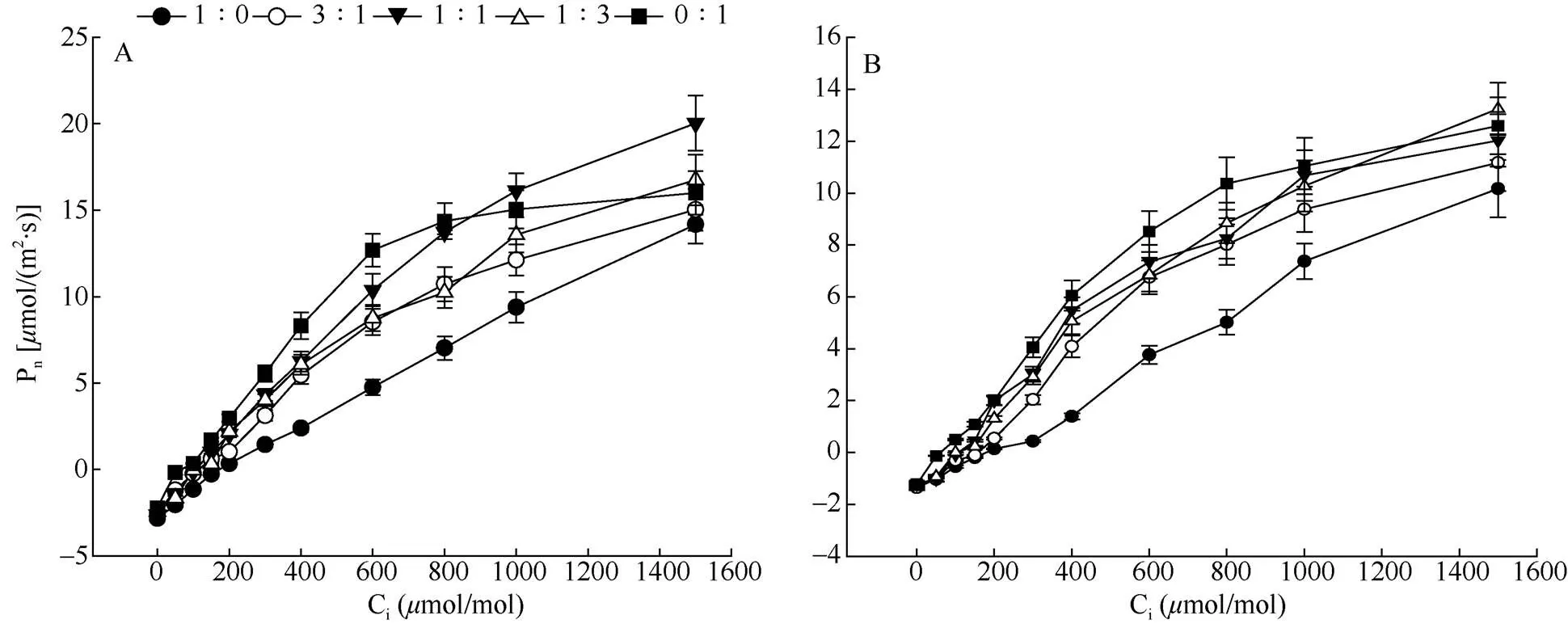
图4 厚叶铁线莲(A)和天台铁线莲(B)的A-Ci曲线。Ci: 胞间CO2浓度。
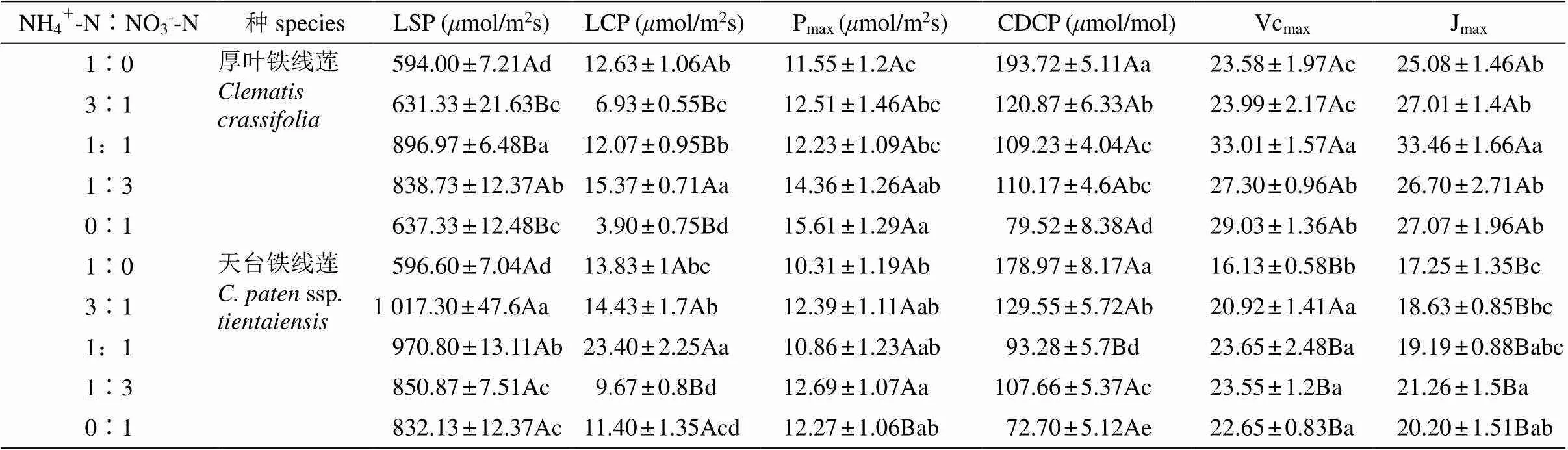
表1 厚叶铁线莲和天台铁线莲光响应曲线与A-Ci曲线的特征参数
LSP: 光饱和点; LCP: 光补偿点; Pmax: 饱和最大光合速率; CDCP: CO2补偿点; Vcmax: 最大RuBP羧化速率; Jmax: RuBP再生的光合电子传递速率。
LSP: Light saturation point; LCP: Light compensation point; Pmax: Maximum net photosynthetic rate; CDCP: CO2compensation point; Vcmax: Maximum RuBP carboxylation rate; Jmax: Photosynthetic electron transfer rate of RuBP regeneration.
2.5 对氮代谢相关酶活性的影响
氮素配比显著影响了铁线莲叶片的GS、GDH、GOGAT活性(表2)。同一氮素配比下厚叶铁线莲与天台铁线莲的GS、GDH、GOGAT活性差异显著(图5)。在配比为1∶3时,厚叶铁线莲的GS、GDH和GOGAT活性显著高于其他氮素配比;而在配比为1∶0时,GS、GDH和GOGAT活性最低,配比为1∶3时的3种酶活性分别是配比为1∶0时的3.56、2.45和6.4倍。天台铁线莲的GS活性在配比为3∶1时最高,显著高于其他配比;GDH活性在配比为1∶3和3∶1时的差异不显著,但显著高于其他配比; GOGAT活性在配比为1∶1时最高,显著高于其他配比。
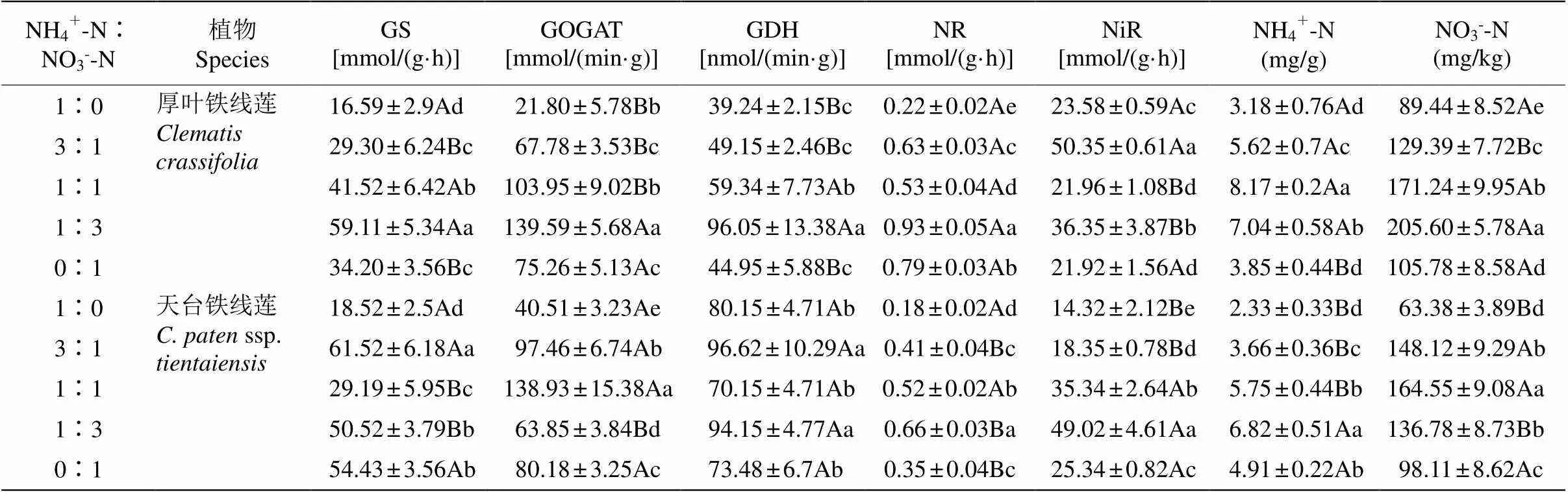
表2 厚叶铁线莲和天台铁线莲的GS、GOGAT、GDH、NR、NiR活性及铵态氮和硝态氮含量
GS: 谷氨酰胺合成酶; GOGAT: 谷氨酸合成酶; GDH: 谷氨酸脱氢酶; NR: 硝酸还原酶; NiR: 亚硝酸还原酶; NH4+-N: 铵态氮; NO3--N: 硝态氮。
GS: Glutamine synthetase; GOGAT: Glutamate synthase; GDH: Glutamate dehydrogenase; NR: Nitrate reductase; NiR: Nitrite reductase; NH4+-N: Ammonium nitrogen;NO3--N: Nitrate nitrogen.
2.6 对硝酸还原酶和亚硝酸还原酶活性的影响
厚叶铁线莲与天台铁线莲在不同氮素配比下的硝酸还原酶(NR)和亚硝酸还原酶(NiR)活性差异十分显著(表2)。厚叶铁线莲与天台铁线莲的NR在铵态氮与硝态氮为1∶3时最大,显著高于其他处理。在配比为3∶1时,厚叶铁线莲的NiR最高, 显著高于其他处理组;而天台铁线莲在配比为1∶3时最大,显著高于其他处理。
2.7 不同氮素处理对铁线莲铵态氮和硝态氮含量的影响
在不同氮素配比下,厚叶铁线莲和天台铁线莲叶片内铵态氮与硝态氮含量差异十分显著(表2)。厚叶铁线莲叶片的铵态氮含量在配比为1∶1时显著高于其他处理;而硝态氮含量则在配比为3∶1时显著高于其他处理。天台铁线莲则相反,在配比为3∶1时铵态氮含量最高,且显著高于其他处理;而在配比为1∶1时硝态氮含量最大,显著高于其他处理。
3 结论和讨论
植物适应生长环境的重要表现之一是对氮素的催化、同化和利用能力。氮素形态的变化会直接对植株产生不同的生理效应,从而导致植物的生长发育不同。大量研究表明铵态氮和硝态氮都可以为植物的生长发育提供足够的氮源,但二者共同配合的情况下可能更好[20]。本研究表明,厚叶铁线莲与天台铁线莲的生物量在铵态氮与硝态氮共同作用下显著高于单一氮源处理,这与高志等[21]对玉米()的研究结果相似。但本研究亦表明,两种铁线莲的最适铵态氮与硝态氮配比不同。以铵态氮作为单一氮源时,植物需要消耗较大能量进行氮素转化,而硝态氮则相反。二者配比能更好的促进植物生长,主要是因为硝态氮的吸收可以减少植物的能量消耗,同时可以吸收大量的阳离子,这些阳离子可以增加细胞的渗透势,从而促进细胞的伸长和植株的生长[22]。
叶绿素是植物吸收转化光能的主要功能分子,光合作用是植物最基本的物质代谢和能量代谢,是植物生长发育的基础,叶绿素含量直接影响植物的光合速率及光合产物的产生[23]。本研究表明,在不同的氮素配比下,厚叶铁线莲与天台铁线莲叶片的叶绿素含量差异显著,并分别在配比为1∶1与1∶3时最高,表明不同氮源的供应可以影响叶绿素的含量和比例,从而影响铁线莲对光能的吸收和转化,进而改变光化学效率和光合产物的积累,但不同种的铁线莲的适宜氮素配比不同。光响应曲线是了解植物的基本光合特性和光化学效率的重要途径之一[24]。LCP和LSP的大小直接反映植物对光的利用能力,体现植物最基本的光需求和光需特性。本研究结果表明,不同氮素配比下厚叶铁线莲和天台铁线莲的LCP、LSP和Pmax均显著高于单一氮源处理,这与邢素芝等[25]的研究结果相似,表明铵态氮与硝态氮相互作用改变了铁线莲的氮素利用效率, 影响了叶绿素的含量和形态建成,使其更有利于吸收利用光能,进而提高铁线莲的光合速率,促进生长。CO2作为光合反应的基本原料,其浓度变化直接影响植物的光合效率。朱宣全等[26]的研究表明, 氮素的施用对植物的A-Ci曲线有调控作用。本研究结果也表明,不同氮素配比下铁线莲的CO2补偿点差异显著,表明在氮素形态配比影响铁线莲的呼吸消耗,且在铵态氮为单一氮源下, 2种铁线莲的光补偿点最高,说明呼吸消耗最大,这可能与铵态氮转化过程中的能量消耗有关,能耗的增加在一定程度上减弱了植物生长。Vcmax表示叶片在单位时间单位面积固定的最大CO2量,它决定了植物的Pmax、光呼吸等过程[27-28]。本研究表明, 混合氮源下铁线莲的Vcmax、Jmax显著高于单一氮源, 表明铵态氮与硝态氮混合施用促进了铁线莲的相关酶合成,有利于增强铁线莲叶片光合电子传递相关酶的催化效率, 有效促进光合作用,进而增强植物生长。
GS是植物氮或氨同化途径“GS/GOGAT途径”的关键酶之一,其活性直接反映氮素的同化能力[29]。GDH在氨的再同化过程中参与氨基酸降解的氧化脱氨作用,对于谷氨酸的催化合成具有重要作用[29]。本研究结果表明,铵态氮与硝态氮的混合施用可以有效增强天台铁线莲和厚叶铁线莲叶片的GS、GOGAT和GDH活性,表明2种氮源混合施用使铁线莲的GS、GOGAT和GDH活性提高,同时启用GS/GOGAT氮同化途径,快速转化为有机氮,供其吸收利用。NR是硝酸盐同化的起始酶和限速酶, 其活性很大程度上受到木质部运送至叶片的硝态氮流量的限制,且直接影响植物对无机氮的转化利用[30]。本研究结果表明,混合氮源下铁线莲的NR与NiR活性显著高于单一氮源,可能是硝态氮作为单一氮源时会产生高浓度的NO3-,从而抑制根部NO3-向叶片中的运输,而铵态氮作为单一氮源时, NH4+会作为NO3-还原反应的终产物产生反馈机制,导致NR与NiR活性降低。植物体内氮素的形态及其含量也直接体现其氮素的存储和代谢利用水平, 研究表明,铁线莲叶片中同时存在铵态氮与硝态氮,但硝态氮含量较高,且施用混合氮源时的铵态氮与硝态氮含量显著高于单一氮源,这与刘春娜等[29]的研究结果相似,表明植物硝态氮吸收能量主要来源于光合器官,因为叶片可能是硝态氮的主要利用部位。氮素形态的不同影响植物同化吸收氮素的能量消耗和形态建成,供应混合氮素可大大减少其同化利用过程中的能耗,并影响植物的色素含量及形态建成,促使其向更有利于增强光合利用效率方面的生长发育。有研究表明[31],铵态氮的大量积累通常会抑制铵态氮的吸收,而植物对硝态氮的吸收可能会缓解过多吸收NH4+-N而产生铵毒。
适宜的铵态氮与硝态氮混合营养供应可显著提高厚叶铁线莲与天台铁线莲的生长、光合色素含量、最大净光合速率和氮代谢相关酶NR、GS、DDH等活性,增强氮素的转化利用能力。本研究结果表明,不同氮素形态配比显著增加厚叶铁线莲与天台铁线莲的生物量,提高叶片的Pmax、Vcmax、Jmax, NR和GS活性及NH4+-N和NO3--N含量也升高。但两种植物的最适宜铵态氮与硝态氮配比不同,厚叶铁线莲适宜的NH4+-N∶NO3--N为1∶1,而天台铁线莲更适宜于1∶3。今后,在栽培生产中,可根据铁线莲对氮素形态需求的差异有针对性地施用氮肥, 从而有效控制生长,为铁线莲的大规模生产提供良好的基础。
[1] YU F H, JIANG J L, SUN L. Research progress on effects of different fertilizers on crop growth [J]. Mod Agric Sci Technol, 2018(4): 9-10. doi: 10.3969/j.issn.1007-5739.2018.04.006.于富豪, 姜金玲, 孙璐. 不同肥料对作物生长的影响研究进展 [J]. 现代农业科技, 2018(4): 9-10. doi: 10.3969/j.issn.1007-5739.2018.04.006.
[2] Zou W T, Li H Z, Lin W J, et al. Effects of the content of available nitrogen in water on the growth and enzymatic activity of[J]. J Anhui Agric Univ, 2011, 38(6): 952-956.邹文桐, 李海珍, 林文娇, 等. 氮素水平对宽杆芥菜幼苗生长及酶活性的影响 [J]. 安徽农业大学学报, 2011, 38(6): 952-956.
[3] ROTHSTEIN D E, CREGG B M. Effects of nitrogen form on nutrient uptake and physiology of Fraser fir () [J]. For Ecol Manag, 2005, 219(1): 69-80. doi: 10.1016/j.foreco.2005.08.043.
[4] XING Y, MA X H. Research progress on effect of nitrogen form on plant growth [J]. J Agric Sci Technol, 2015, 17(2): 109-117. doi: 10. 13304/j.nykjdb.2014.574.邢瑶, 马兴华. 氮素形态对植物生长影响的研究进展 [J]. 中国农业科技导报, 2015, 17(2): 109-117. doi: 10.13304/j.nykjdb.2014.574.
[5] ABUZINADAH R A, READ D J. The role of proteins in the nitrogen nutrition of ectomycorrhizal plants: IV. The utilization of peptides by birch (L.) infected with different mycorrhizal fungi [J]. New Phytol, 1989, 112(1): 55-60. doi: 10.1111/j.1469-8137.1989.tb 00308.x.
[6] WANG W C, LI L Q. A new system of classification of the genus(Ranunculaceae) [J]. Acta Phytotaxon Sin, 2005, 43(5): 431- 488. doi: 10.1360/aps040130. 王文采, 李良千. 铁线莲属一新分类系统 [J]. 植物分类学报, 2005, 43(5): 431-488. doi: 10.1360/aps040130.
[7] GUAN K Y, LI Z J, LI J X, et al. A preliminary study on the intro- duction and cultivation of[J]. Acta Bot Yunnan, 2002, 24(3): 392-397. doi: 10.3969/j.issn.2095-0845.2002.03.014.管开云, 李志坚, 李景秀, 等. 铁线莲属植物的引种栽培研究初报 [J]. 云南植物学报, 2002, 24(3): 392-397. doi: 10.3969/j.issn.2095- 0845.2002.03.014.
[8] HUANG M, DUAN Y Q, LI Y J, et al. Effects of different nitrogen form ratios on the nitrogen accumulation and utilization of winter wheat [J]. J Soil Water Conserv, 2013, 27(6): 290-293,304. doi: 10. 3969/j.issn.1009-2242.2013.06.056.黄明, 段有强, 李友军, 等. 不同氮素形态配比对冬小麦氮素积累和利用的影响 [J]. 水土保持学报, 2013, 27(6): 290-293, 304. doi: 10.3969/j.issn.1009-2242.2013.06.056.
[9] TAN J K, ZHANG Y L, SHEN Q R, et al. Effects of different NH4+-N/ NO3--N ratios on water use efficiency and its biological characteristics of rice at early stage [J]. J Nanjing Agric Univ, 2002, 25(3): 49-52. doi: 10.3321/j.issn:1000-2030.2002.03.012.谈建康, 张亚丽, 沈其荣, 等. 不同形态氮素比例对水稻苗期水分利用效率及其生物效应的影响 [J]. 南京农业大学学报, 2002, 25(3): 49-52. doi: 10.3321/j.issn:1000-2030.2002.03.012.
[10] LIU R, SHI F, LIU W C, et al. Effect of nitrogen forms on cell ultra- structure and photosynthesis of tomato under salinity [J]. Acta Hort Sin, 2015, 34(3): 471-479. doi: 10.16420/j.issn.0513-353x.2014-0821.刘冉, 石峰, 刘伟成, 等. 不同形态氮素对盐胁迫下番茄细胞超微结构与光合作用的影响 [J]. 园艺学报, 2015, 34(3): 471-479. doi: 10.16420/j.issn.0513-353x.2014-0821.
[11] Liu Y H. Studies in mechanism of ammonium toxicity in tomato (Mill.) under different light intensities [D]. Hangzhou: Zhejiang University, 2004: 1-56. 刘永华. 不同光强下番茄铵毒害机理的研究 [D]. 杭州: 浙江大学, 2004: 1-56.
[12] XIONG Q E. Plant Physiology [M]. Chengdu: Sichuan Science and Technology Press, 2003: 122-128. 熊庆娥. 植物生理学实验教程[M]. 成都: 四川科学技术出版社, 2003: 122-128.
[13] LONG S P, BERNACCHI C J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Proce- dures and sources of error [J]. J Exp Bot, 2003, 54(392): 2393-2401. doi: 10.1093/jxb/erg262.
[14] GAO J F. Experimental Guidance for Plant Physiology [M]. Beijing: Higher Education Press, 2006: 74-77. 高俊凤. 植物生理学实验指导 [M]. 北京: 高等教育出版社, 2006: 74-77.
[15] QIAO F L. Determination Technique of Plant Physiology Experiment Analysis [M]. Beijing: China Agricultural Science and Technology Press, 2002: 78-82. 乔富廉. 植物生理学实验分析测定技术 [M]. 北京: 中国农业科学技术出版社, 2002: 78-82.
[16] LOULAKAKIS K A, ROUBELAKIS-ANGELAKIS K A. Intracellular localization and properties of NADH-glutamate dehydrogenase fromL.: Purification and characterization of the major leaf isoenzyme [J]. J Exp Bot, 1990, 41(10): 1223-1230. doi: 10.1093/jxb/ 41.10.1223.
[17] MA Z Y, SHAO L H, LIU L F, et al. Corn leaves glutamate synthase (GOGAT) purification and characteristic research[J]. J Maize Sci, 2009, 17(5): 82-84. 马振勇, 邵立红, 刘利锋, 等. 玉米叶片谷氨酸合酶(GOGAT)的纯化和特性研究 [J]. 玉米科学, 2009, 17(5): 82-84.
[18] SUN S Q, YAN X F. Determination method of nitrate reductase activity in the leaves of[J]. J NE For Univ, 2004, 32 (3): 83-84. doi: 10.3969/j.issn.1000-5382.2004.03.027.孙世芹, 阎秀峰. 喜树叶片硝酸还原酶活性的测定方法 [J]. 东北林业大学学报, 2004, 32(3): 83-84. doi: 10.3969/j.issn.1000-5382. 2004.03.027.
[19] Shanghai Institute of Plant Physiology, Chinese Academy of Sciences. Guide to Modern Plant Physiology Experiments [M]. Beijing: Science Press, 1999: 261-262. 中国科学院上海植物生理研究院, 上海市植物生理学会. 现代植物生理学实验指南[M]. 北京: 科学出版社, 1999: 261-262.
[20] XU J L, BIE Z L, ZHANG S L. Effects of nitrogen form ratios on the lettuce growth and activity of protective enzymes [J]. J Huazhong Agric Univ, 2005, 24(3): 290-294. doi: 10.3321/j.issn:1000-2421.2005. 03.018.徐加林, 别之龙, 张盛林. 不同氮素形态配比对生菜生长、品质和保护酶活性的影响 [J]. 华中农业大学学报, 2005, 24(3): 290-294. doi: 10.3321/j.issn:1000-2421.2005.03.018.
[21] GAO Z, XU Y C, Shen Q R, et al. Compound fertilizers with different nitrogen forms on the growth and nitrogen use efficiency of maize seedlings [J]. Acta Agric Boreali-Sin, 2005, 20(6): 68-72. doi: 10. 3321/j.issn:1000-7091.2005.06.018.高志, 徐阳春, 沈其荣, 等. 不同氮素形态配比的复混肥对玉米苗期生长及氮肥利用效率的影响 [J]. 华北农学报, 2005, 20(6): 68-72. doi: 10.3321/j.issn:1000-7091.2005.06.018.
[22] WANG G Y. Effects of different nitrogen forms and combined with plant hormone on growth, nutrient uptake and their distribution in tobacco plants [D]. Baoding: Hebei Agricultural University, 2002: 19-28.王国英. 不同形态氮素及植物激素对烟草生长、养分吸收和分配的影响 [D]. 保定: 河北农业大学, 2002: 19-28.
[23] ZHANG J Z, LIN G Z, LIN Z F. et al. Response of photosynthesis to growth light intensity in some south subtropical woody plants [J]. J Trop Subtrop Bot, 2005, 13(5): 413-418. doi: 10.3969/j.issn.1005-3395. 2005.05.009.张进忠, 林桂珠, 林植芳, 等. 几种南亚热带木本植物光合作用对生长光强的响应 [J]. 热带亚热带植物学报, 2005, 13(5): 413-418. doi: 10.3969/j.issn.1005-3395.2005.05.009.
[24] SHARP R E, MATTHEWS M A, BOYER J S. Kok effect and the quantum yield of photosynthesis: Light partially inhibits dark respi- ration [J]. Plant Physiol, 1984, 75(1): 95-101. doi: 10.1104/pp.75.1.95.
[25] XING S Z, WANG J F, LI X L, et al. Different nitrogen fertilizers and ratios effect on growth, safety and quality of spinach [J]. J Plant Nutri Fertil, 2015, 21(2): 527-534. doi: 10.11674/zwyf.2015.0229.邢素芝, 汪建飞, 李孝良, 等. 氮肥形态及配比对菠菜生长和安全品质的影响 [J]. 植物营养与肥料学报, 2015, 21(2): 527-534. doi: 10.11674/zwyf.2015.0229.
[26] ZHU X Q, JIA M, SHI P Y, et al. Fitting analysis of CO2response curve of tobacco under different nitrogen fertilizer levels [J]. J S Agric, 2020, 51(3): 537-544. doi: 10.3969/j.issn.2095-1191.2020.03.007.朱宣全, 贾孟, 史普酉, 等. 不同氮肥水平下烟草CO2响应曲线的拟合分析 [J]. 南方农业学报, 2020, 51(3): 537-544. doi: 10.3969/j. issn.2095-1191.2020.03.007.
[27] FARQUHAR G D, VON CAEMMERER S, BERRY J A. A bioche- mical model of photosynthetic CO2assimilation in leaves of C3species [J]. Planta, 1980, 149(1): 78-90. doi: 10.1007/BF00386231.
[28] WULLSCHLEGER S D. Biochemical limitations to carbon assimi- lation in C3plants: A retrospective analysis of the/Ccurves from 109 species [J]. J Exp Bot, 1993, 44(5): 907-920. doi: 10.1093/jxb/44.5.907.
[29] LIU C N. Nutrition effects of different nitrogen onseedlings [D]. Harbin: Northeast Forestry University, 2010: 43-50. 刘春娜. 落叶松幼苗对不同形态氮的营养响应 [D]. 哈尔滨: 东北林业大学, 2010: 43-50.
[30] LIN J F, LIN Y Z, QIU W C, et al. Comprehensive evaluation analysis on cultivation effects of lettuce under different nitrogen levels [J]. J Trop Subtrop Bot, 2013, 21(5): 432-438. doi: 10.3969/j.issn.1005-3395. 2013.05.009.林俊芳, 林义章, 丘武昌, 等. 不同氮素水平对油麦菜栽培效果的综合评价分析 [J]. 热带亚热带植物学报, 2013, 21(5): 432-438. doi: 10.3969/j.issn.1005-3395.2013.05.009.
[31] YE L S, CHEN S L. Effects of nitrogen forms and ratios on photo- synthetic characters and enzyme activities in nitrogen metabolism of[J]. Chin J Ecol, 2016, 35(9): 2355-2360. doi: 10.13292/j.1000-4890.201609.025.叶莉莎, 陈双林. 氮素形态及配比对雷竹光合特性和氮代谢酶活性的影响 [J]. 生态学杂志, 2016, 35(9): 2355-2360. doi: 10.13292/j. 1000-4890.201609.025.
Effect of Nitrogen Forms on Photosynthetic Characteristics and Nitrogen Metabolismof
MA Xiao-hua1*, HU Qing-di1, ZHANG Yan-jun2, QIAN Ren-juan1, ZHENG Jian1*, LIU Hong-jian1
(1. Zhejiang Institute of Subtropical Crops,Wenzhou 325005, Zhejiang, China; 2. Bamboo Research Center, National Forestry and Grassland Administration, Hangzhou 310000, China)
In order to understand the photosynthetic characteristics and response mechanism of nitrogen metabolism in, the growth, light response curve, A-Ci curve and nitrogen metabolism related enzyme activities of 1-year-oldandssp.were studied under different ratios of NH4+-N∶NO3--N. The results showed that the ratio of NH4+-N∶NO3--N significantly affects the biomass and chlorophyll content of clematis, and the biomass, Chl a, Chl b, and Car contents ofreach the maximum under the ratio at 1∶1, while the Chl a and Chl b contents ofsspreach the maximum under the ratio at 1∶3. Under the ratio of 1∶1, the light saturation point (LSP) ofand the light compensation point (LCP) ofssp.were the maximum, and the Vcmaxand Jmaxvalues ofleaves were significantly higher than those under other ratios. The glutamine synthetase (GS) activity ofssp.was the highest under the ratio of 3∶1, while NR and nitrite reductase (NiR) activities under the ratio at 1∶3 were significantly higher than those under other ratioes. Therefore, the mixed application of NH4+-N and NO3--N could effectively promote the growth and photosynthesis, and enhance the nitrogen utilization efficiency of two.was suitable for 1∶1 of NH4+-N to NO3--N, whilessp.was more suitable for 1∶3.
; Nitrogen form; Biomass; Photosynthetic characteristics; Nitrogen metabolism
10.11926/jtsb.4281
2020-07-02
2020-09-14
浙江省公益研究计划项目(LGN18C150006); 浙江省科技计划项目(2019C02036); 温州市重大科技创新攻关项目(ZS2020002)资助
This work was supported by the Program for Public Welfare Research in Zhejiang (Grant No. LGN18C150006); the Program for Science and Technology of Zhejiang (Grant No. 2019C02036); and the Project for Major Scientific and Technological Innovation in Wenzhou (Grant No. ZS2020002).
马晓华,女,助理研究员,主要从事观赏植物生理生化与育种研究。E-mail: maxiaohua1120@126.com
Corresponding author. E-mail: zjyzs@126.com; maxiaohua1120@126.com
