病毒诱导基因沉默在蔬菜作物上应用的研究进展
2021-05-31李杰罗江宏杨萍
李杰,罗江宏,杨萍
红河学院生命科学与技术学院/云南省高校滇南特色生物资源研究与利用重点实验室,云南蒙自 661100
我国蔬菜产业经过几十年的发展,在产值、出口量等方面均位居农作物首位,已成为中国农民增收、加快农村发展的支柱产业,但仍然存在蔬菜品质弱、产量低等问题。目前急需挖掘与蔬菜作物重要农艺性状相关的基因以培育优异、抗病和具有特色的品种[1-2]。随着高通量测序技术的发展,越来越多的蔬菜作物全基因组测序已完成,但对于单个基因的功能研究,还需要更有效和可靠的技术体系。基于植物病毒与其宿主在长期进化过程中形成的共生关系,将外源基因通过病毒载体导入植物体内,可为基因挖掘和基因功能研究提供技术支持[3]。病毒诱导的基因沉默(virus-induced gene silencing,VIGS)是利用RNA介导的植物天然抗病毒机制的一种技术,主要利用遗传免疫方式系统性沉默某一特定基因,再经过表型鉴定和基因表达来确定靶基因在植物生长发育中的作用及对环境变化产生的应激响应。用携带靶基因片段的病毒来侵染植物,植物的遗传免疫系统被激活后,植物体内与靶基因同源的RNA被特异性降解,从而发生基因沉默[4]。与传统的转基因、基因敲除、反义抑制等基因功能研究方法相比,VIGS技术试验周期短,不依赖转基因操作,具有低成本、高通量等优点[5]。近年来,VIGS技术的建立为蔬菜作物功能基因组学的研究提供了优越的条件,在基因功能的研究中得到了越来越广泛的应用,从而促进了我国蔬菜产业的可持续发展。
本文根据国内外相关研究的最新报道,重点对VIGS技术在蔬菜作物生长发育、次生代谢、生物与非生物胁迫等几个方面进行综述,分析VIGS技术存在的问题及解决方法,以期为VIGS技术的进一步应用提供依据。
1 VIGS技术介绍
1.1 VIGS技术机制
VIGS是一种利用植物防御病毒入侵的自然机制来研究植物基因功能的基因转录抑制技术,属于RNA干扰的一种。利用携带植物目的基因片段的病毒载体,通过有效的病毒侵染方式侵染植物,病毒载体在寄主体内复制和表达时,在RNA聚合酶的作用下,产生双链RNA(double strand RNA,dsRNA),dsRNA在细胞中被特异性核酸内切酶切割产生21—25 nt的小分子干扰RNA(small interfering RNA,siRNA),siRNA能诱导与其具有同源性的植物内源基因mRNA降解,可结合形成RNA诱导的沉默复合物(RNA-induced silencing complex,RISC)。一方面,RISC可以特异性地与细胞质中的同源RNA相互作用,使同源RNA降解,从而导致转录后基因沉默(PTGS)[6];另一方面,RISC特异性地与细胞核内的同源DNA相互作用,使其被甲基化等修饰,从而产生转录水平的基因沉默(transcriptional gene silencing,TGS)[7]。
1.2 VIGS的病毒载体及应用
VIGS是目前应用最广泛的研究植物功能基因的工具之一[8]。根据已有的研究报道,有多种病毒载体(RNA病毒、DNA病毒、卫星病毒)在VIGS上成功应用,其中 RNA病毒包括烟草花叶病毒(tobacco mosaic virus,TMV)、马铃薯X病毒(potato virus X,PVX)、烟草脆裂病毒(tobacco rattle virus,TRV)、番茄金色花叶病毒(tomato golden mosaic virus,TGMV)、甘蓝缩叶病毒(cabbage leaf curl virus,CbLCV)等[9]。但每种病毒载体诱导的沉默效率与宿主的选择有关。如 PVX病毒载体在番茄上的研究发现,需要先进行体外转录才能进行RNA注射侵染[10]。TRV病毒载体的沉默效率高,沉默持续性长且病毒症状轻,因而被广泛用于VIGS载体的构建[11]。苹果潜隐球形病毒(apple latent spherical virus,ALSV)因病症温和,且在番茄上不产生病毒症状,而经常用于番茄基因的沉默研究[12]。DNA病毒载体构建简单,试验操作难度低且无需体外转录。其中的甜菜曲顶头病毒(beet curly top virus,BCTV)和番茄卷曲叶病毒(tomato leaf curl virus,ToLCV)已成功用于番茄基因功能的研究[13-14]。卫星病毒基因组小,在宿主中复制速度快且易于遗传。其中的中国番茄黄化曲叶病毒(tomato yellow leaf curl China virus,TYLCV)和烟草曲茎病毒(tobacco curly shoot virus,TbCSV)可高效用于番茄基因的沉默[15-16]。研究者通过传统的“酶切-连接”方法成功构建了甜菜 WRKY转录因子家族成员BvWRKY23的RNA干扰(RNA interference,RNAi)表达载体[17]。
1.3 VIGS技术进展
KUMAGAI等[18]首次以八氢番茄红素脱氢酶(phytoene desaturase,PDS)cDNA片段完成 TMVVIGS载体构建。经过 3年的发展,RUIZ等[19]利用PDS构建了 PVX-VIGS载体。VALENTINE等[20]以TRV病毒为载体完成了 TRV-VIGS体系的构建,TRV-VIGS体系后来常用于蔬菜作物基因沉默载体的构建。近几年,WANG等[21]首次利用大麦条纹花叶病毒(barley stripe mosaic virus,BSMV)建立了BSMVVIGS体系用于大麦基因功能的研究。张怡等[22]成功构建了大豆线虫肌钙蛋白(troponin C,tnc)和酰胺样多肽(FMRFamide-like peptides,flp)基因的 TRV-VIGS体系。随着不同病毒载体 VIGS体系研究的深入,PANDEY等[23]研究出复合病毒体系,将其他植物病毒的miRNA导入到另外一种病毒中,形成外源mirRNAs来侵染植株,开发了MIR-VIGS方法,标志着双病毒载体应用VIGS体系的产生。赵祯等[24]运用Gateway技术成功构建了茄子(Solanumaculeatissimum)TRV-VIGS表达载体,沉默了叶片和果实的SmMsrA,表明基因克隆技术与VIGS技术的结合使用,减少了酶切环节,提高了载体构建的效率。最近,宋恬等[25]首次在扁桃花器官上建立了以TRV为载体的VIGS沉默体系,成功用携带pTRV2-AcCBF1的农杆菌侵染扁桃花器官且出现沉默效果。邱润霜等[26]以番茄斑萎病毒(tomato spotted wilt orthotospovirus,TSWV)N构建到pTRV-PTV00载体上,完成了TSWV介导的NVIGS载体的构建,开阔了构建病毒内源基因沉默体系的新思路。
2 VIGS技术的优点
VIGS研究基因功能的优点在于它构建简易、周期性短、成本较低。一般构建重组载体到侵染植物进行功能鉴定只需几周时间,因此可以大规模进行基因组序列和 EST序列的功能鉴定[27]。VIGS对于难获突变体作物的研究优势突出。当植物基因组较大时,利用靶基因家族的保守DNA区域,VIGS通过沉默一个基因家族的多个成员来避免功能冗余的问题,当涉及到大的蛋白质家族或基因重复时,各种点突变和插入不能产生适当的表型[28],而 VIGS可使序列差异为10%—20%的同源基因沉默[29]。利用VIGS沉默目标基因无需靶基因的完整编码序列,只需其中的一段 cDNA片段即可实现靶基因沉默[30]。而RNAi技术在应用时,dsRNA的合成成本过高,不利于产品的推广[31]。CRISPR/Cas9技术实现基因编辑有赖于PAM序列,在附近无PAM序列的区域时无法进行编辑[32]。VIGS具备将 cDNA文库整合到病毒载体上进行高通量基因筛选分析的优势[33]。VIGS技术可用于不同蔬菜物种间比较基因组学的研究[34]。
3 VIGS在茄果类蔬菜作物中的应用
蔬菜作物分子学研究领域已进入后基因组时代[35]。应用 VIGS技术鉴定茄果类蔬菜的基因功能涉及物质合成调控、生物和非生物应激反应等方面。根据国内外近3年的最新文献,重点介绍在茄果类蔬菜果实发育、激素调节和植物与病原微生物相互作用及非生物胁迫应答等方面利用 VIGS技术的研究进展(表1)。

表1 利用VIGS研究的茄果类蔬菜基因及其表型Table 1 List of genes silenced by VIGS and the phenotype observed in solanaceous vegetable
3.1 VIGS在茄果类蔬菜物质合成调控中的研究应用
糖类的合成与转运在植物新陈代谢与物质循环中起着关键作用,同时也受到多方面的调控。利用VIGS技术,目前已经系统研究了茄果类蔬菜中糖类合成途径中的一些关键调控因子的功能。通过基因编辑技术,研究基因表达的激活(activation)和抑制(repressor)[57]。例如在番茄酰基糖代谢中间产物直链脂肪酸(straight-chain fatty acid,SCFA)合成途径KASII和KASIII功能的研究中,沉默该基因后,SCFA含量减少了 40%,同时沉默植株中KASII和KASIII表达量显著降低[36]。那么糖类是如何由新叶向老叶转运呢?最新的一项研究通过 VIGS技术沉默SWEET家族基因SISWEET1a发现,沉默番茄植株的幼叶中己糖积累量减少了50%,在成熟叶片中增加了2倍多,证明SISWEET1a在番茄叶片合成的糖分向幼叶转运的过程中起着关键作用,挖掘出了SISWEET1a一个新的功能[37],说明基因编辑技术能对家族基因中单个基因准确定位,并进行稳定改造,挖掘其基因功能[58]。
辣椒素是辣椒果实风味品质的重要指标,并且在其生长中起着防虫防病的自我防御作用。茉莉酸(jasmonic acids,JAs)在调控辣椒素的代谢信号转导中起着非常重要的作用,其调控因子R2R3-MYB的转录因子CaMYB108主要在辣椒的果实和花中表达,但是CaMYB108的下游信号尚未确定。利用VIGS技术沉默CaMYB108后发现辣椒素生物合成基因CBGs表达量降低,辣椒素含量减少,花药开裂延迟,同时花粉活力降低。通过双荧光素酶报告基因检测发现,CaMYB108靶向启动CBG。另外,茉莉酸甲酯诱导了CaMYB108和CBGs的表达,从而证实了CaMYB108参与辣椒植株雄蕊的发育以及辣椒素的合成[38]。以上研究都是基于TRV载体诱导的VIGS方法,而另外一种ALSV载体侵染植株后虽然没有明显的外观特征,但其部分基因序列经过改造后也常用于VIGS体系的构建。有研究者发现,利用ALSV病毒侵染的藜麦叶片摩擦接种到辣椒上的方式并不能实现侵染,但使用ALSV病毒浓缩液则侵染成功,且沉默效率达到90%。在ALSV介导的VIGS体系中,将氨基酸转移酶基因(pAMT)导入 ALSV病毒载体的方式成功侵染了辣椒植株,且侵染率在 80%—90%,pAMT沉默植株的辣椒素含量和辣椒素酯的积累减少。以上研究结果表明,ALSV病毒能够用于构建VIGS体系来研究辣椒素积累的调控机制,从而可以在辣椒育种上通过基因改造来提高辣椒素的含量[39]。
番茄果实中类胡萝卜素的积累受环境和激素的影响较大,在调控番茄果实类胡萝卜素代谢中,螺旋-环-螺旋转录因子(Helix-Loop-Helix,SLAR)影响类胡萝卜素的合成。为了挖掘SLAR的功能,研究者利用VIGS技术沉默普通番茄‘M82’和樱桃番茄‘Micro Tom’的SLAR,发现两个品种具有相同的表型,而且沉默后番茄红素、总类胡萝卜素和叶绿素含量明显减少,验证了SLAR直接参与番茄类胡萝卜素的生物合成过程[40]。有研究者利用CRISPR/Cas9技术分别以八氢番茄红素合成酶1(phytoene synthase 1,PSY1)、MYB12和花色苷2(Anthocyanin 2,ANT2)为靶位点,成功培育出黄色、粉红色和紫色番茄[59],但构建载体时需要准确的靶基因序列,且容易脱靶。糖苷生物碱属于有毒物质,存在于许多茄科植物中,其合成受遗传和环境的影响,尤其是光环境。叶绿素和类胡萝卜素的生物合成也依赖光信号转导途径,同时在糖苷生物碱合成过程中具有相同的中间物质。研究者通过VIGS沉默PDS和镁螯合酶基因CHLI与CHLH,发现沉默茄子叶片中叶绿素和类胡萝卜素含量明显降低,通过高效液相色谱和代谢物全谱分析,发现沉默植株糖苷生物碱含量显著降低,参与糖苷生物碱合成和其他代谢物合成的基因下调表达,而且发现光合色素的积累会影响甾族类糖苷生物碱在茄子叶片中的合成[41]。在茄子果实成熟过程中,查尔酮、花青素等类黄酮物质在其角质层和表皮细胞中有所积累,类黄酮在茄子果实成熟过程中的调控机制和影响因素也利用 VIGS技术得到了解答。研究者利用VIGS技术沉默茄子查尔酮合成酶基因(SmCHS),发现沉默植株茄子果实颜色变浅,花青素含量降低,表皮细胞的大小和形状都发生了变化,且沉默植株茄子果实的向重力性反应减轻,茄子果实弯曲,这一表型不仅说明CHS调控茄子果实中类黄酮的积累,还证明了表皮细胞的延展性依赖CHS进行表达,从而揭示了CHS的新功能[42]。许志茹等[60]利用GATEWAY技术构建了芜菁(Brassicarapa)二氢黄酮醇4-还原酶(dihydroflavonol 4-reductase,DFR)基因过表达的RNAi载体,为鉴定DFR在依光型和非依光型花青素合成过程中的功能研究奠定了基础。同样是色素相关基因功能的研究,VIGS体系不需要构建植物遗传转化体系,大大缩短了研究进程。生长素作为植物适应环境变化和植物组织代谢的关键激素,其在植物离层区域如何调控植物组织离层尚不清楚。相关研究者对此展开了研究,在对番茄生长素共轭水解酶相关的5个基因功能的研究中发现,IAA-Ile参与生长素共轭水解过程,并作为候选基因存在于离层区。利用环己酰亚胺(cycloheximide,CHX)处理发现从头合成生长素共轭水解酶抑制组织离层。利用 VIGS技术沉默番茄SlILL1、SlILL3、SlILL5、SlILL6和SlILL7,发现沉默目标基因不会影响其他SlILL的表达。同时发现离层区生长素能够在花药离层中发挥作用,是因为受到其浓度的影响,且SlILR1、SlILL5和SlILL6存在协同作用,而外源使用生长素后植株沉默表型明显减弱[43]。
3.2 VIGS在茄果类蔬菜生物胁迫应答机制中的研究应用
番茄卷叶病毒(tomato leaf curl virus,TolCV)侵染番茄后诱导了乙烯反向调控途径信号基因LeCTR1的表达。利用VIGS技术构建番茄LeCTR1沉默植株,发现沉默植株对TolCV病毒的抗性增加,卷叶症状减轻,活性氧(reactive oxygen species,ROS)积累显著减少,主要抗病相关基因(NPR1、PR1、PR5、AOS2、EIN2、EIN3和ERF5)表达增加,说明沉默LeCTR1可以增强番茄对卷叶病的抗性[44]。DEKs参与植物细胞的增殖、分化、衰老和死亡。利用VIGS技术沉默番茄DEK后,番茄对葡萄孢菌(Botrytiscinerea)和丁香假单胞菌(Pseudomonassyringaepv.TomatoDC3000,Pst DC3000)的抗病性降低,沉默番茄植株中ROS积累增加,而且下调了抗病基因的表达,从而证明了DEKs参与番茄抗病的基因功能[45]。细菌性枯萎病是由青枯病菌(Ralstoniasolanacearum)引起的,而其在茄子上的致病机理一直不清楚。研究者利用VIGS技术来探究亚精胺合成酶(spermidine synthase,SPDS)在茄子青枯病中的作用机理,发现接种青枯病菌后茄子SPDS被诱导表达,亚精胺(Spd)含量增加,尤其是在耐性系植株中更加明显,推断 Spd在茄子抗青枯病中有着重要的作用。之后又通过酵母单杂和荧光素酶报告基因试验来验证这一推论,发现R2R3-MYB转录因子SmMYB44确实可以和SPDS启动子直接结合而诱导基因表达,进一步通过超表达SmMYB44茄子植株证明了抗青枯病的SmMYB44-SmSPDS-Spd机制[46]。在辣椒上也利用 VIGS技术证明了青枯病的防御机制,研究发现 MYB转录因子CaPHL8参与辣椒青枯病的防御,通过氨基酸序列分析表明PHL8为 MYB转录因子,接种病菌植株的CaPHL8上调表达,而且GUS活性检测也证明了这一结论。通过VIGS技术证明了CaPHL8在辣椒抗青枯病中的作用,而且这个基因在高温高湿条件下也不会诱导表达[47]。在番茄青枯病的研究中,利用VIGS技术沉默MTC关联基因SAMS2、SAHH1和MS1以及GABA生物合成相关基因GAD2和SSADH1,发现高浓度和中浓度侵染的SAHH1、MS1和GAD2沉默番茄对青枯病的抗性减弱,从而验证了MTC和GABA生物合成在番茄青枯病中的致病作用[48]。在探索粉孢菌(Oidiumneolycopersici)引起番茄白粉病的研究中,利用VIGS技术构建了ShORR-1(solanum habrochaites oidium resistance required-1)沉默株系,证明ShORR-1在番茄抗白粉病中的基因功能[49]。辣椒CaWRKY45参与抗病的功能也利用VIGS技术得以验证[50]。
植物根系分泌物和根结线虫均会引起土壤性状的改变,根结线虫也会侵染植物根系。在最新的一项关于植物、土壤和根结线虫的报道中,研究者利用VIGS技术沉默番茄根系 ABC转运蛋白基因ABC-C6和ABC-G33,其中ABC-C6沉默植株抑制结瘤线虫(Meloidogynespp.)和黄金线虫(Globoderaspp.)产卵,ABC-G33沉默株系只抑制黄金线虫产卵。ABC-C6沉默植株对土壤农杆菌(Agrobacteriumtumefaciens)和枯草芽孢杆菌(Bacillussubtilis)没有抑制作用,但ABC-G33沉默植株对枯草芽孢杆菌具有抑制作用,发现ABC-C6在生物防治根结线虫上的重要作用,从而证明了 ABC转运蛋白基因在土壤根系分泌物和线虫之间的关系,为保护植物免受线虫侵染提供了依据[51]。在茄子上的研究发现,核苷酸结合位点富含亮氨酸的重复序列nucleotide-binding site leucine-rich repeat(NBS-LRR)抗逆基因SacMi,在接种线虫的植株中该基因表达量上调,沉默SacMi植株表现出对线虫的敏感性[52]。
3.3 VIGS在茄果类蔬菜非生物胁迫应答机制中的研究应用
CaWRKY27能够正调控辣椒植株对细菌性青枯病的抗性,同时负调控植株的耐热性。研究者利用VIGS技术沉默CaWRKY27,发现沉默的辣椒植株对盐胁迫和渗透胁迫抗性显著增强,还发现CaWRKY27不仅在辣椒逆境胁迫中是转录激活因子,而且还能编码抗逆境阻遏蛋白[53]。谷胱甘肽转移酶基因(GSTU43)参与了5-氨基乙酰丙酸(5-aminolevulinic acid,ALA)诱导的抗氧化胁迫,并调节低温胁迫下叶绿素的合成。利用VIGS技术沉默GSTU43后,发现沉默植株中谷胱甘肽S-转移酶(glutathione S-transferase,GST)、超氧化物歧化酶、过氧化氢酶、抗坏血酸氧化酶和谷胱甘肽还原酶活性降低,膜脂过氧化物含量增加,而外源ALA处理后这些指标均恢复,说明ALA可引起由GSTU43编码的GST增加,从而增强番茄对低温引起的氧化胁迫抗性[54]。研究人员在番茄中利用CRISPR/Cas9系统构建 CRISPR-BZR1突变体,证实BZR1还通过调节细胞膜受体蛋白激酶FERONIA基因(FER)参与番茄的耐热性[61],但整个番茄植株材料均在bzr1突变体的基础上才能完成。
F-box蛋白、CaDIF1(辣椒干旱诱导F-box protein1基因)可以由ABA、干旱胁迫、H2O2和NaCl胁迫诱导表达,CaDIF1沉默株系展现出干旱敏感性,而超表达株系展现出 ABA敏感性和干旱耐性。利用酵母双杂交试验确定了CaDIS1(辣椒 DIF1-Interacting SKP1)在细胞质和细胞核中与CaDIF1存在互作现象。CaDIF1和CaDIS1沉默株系表现出对ABA不敏感以及干旱耐性降低,且两种植株叶片气孔变大和蒸腾速率增加,进一步说明CaDIF1和CaDIS1可互作,并且参与依赖 ABA信号转导的干旱胁迫抗性过程[55]。研究者也利用VIGS技术揭示了MYB80参与调控辣椒低温胁迫的机制,发现低温处理下MYB80沉默植株抗冷性减弱[56]。
4 VIGS在瓜类、叶菜类和豆类蔬菜作物中的应用
表2列举了利用VIGS技术沉默瓜类、叶菜类和豆类蔬菜相关功能基因及沉默后的表型。瓜类蔬菜有黄瓜、西葫芦、丝瓜、香瓜和西瓜等,VIGS试验一般使用ALSV病毒作为载体侵染葫芦科作物[69]。克隆黄瓜叶片PDS和SU(300 bp),分别构建ALSV-PDS和 ALSV-SU两个载体侵染葫芦科植物的子叶,沉默植株分别表现出光漂白和黄叶表型,说明导入ALSV-PDS和ALSV-SU载体的植物内源PDS和SU被沉默,表明ALSV介导的VIGS体系成功构建[28]。研究者还利用 TRV介导的 VIGS技术构建黄瓜glycerol-3-phosphate 2-O-acyltransferase 6(GPAT6)基因沉默体系,探究GPAT6在黄瓜自毒胁迫中的功能,沉默植株表现出对自毒物质肉桂酸的抗性增强[62]。在甜瓜(Cucumismelovar.makuwaMakino)CmLOX10功能验证中,沉默植株CmLOX10表达量下调,pTRV-CmPDS沉默植株叶片出现白化现象,绿色荧光蛋白(GFP)检测的结果进一步确认了沉默现象,证明 TRV病毒诱导的基因沉默体系在甜瓜上构建成功[63]。
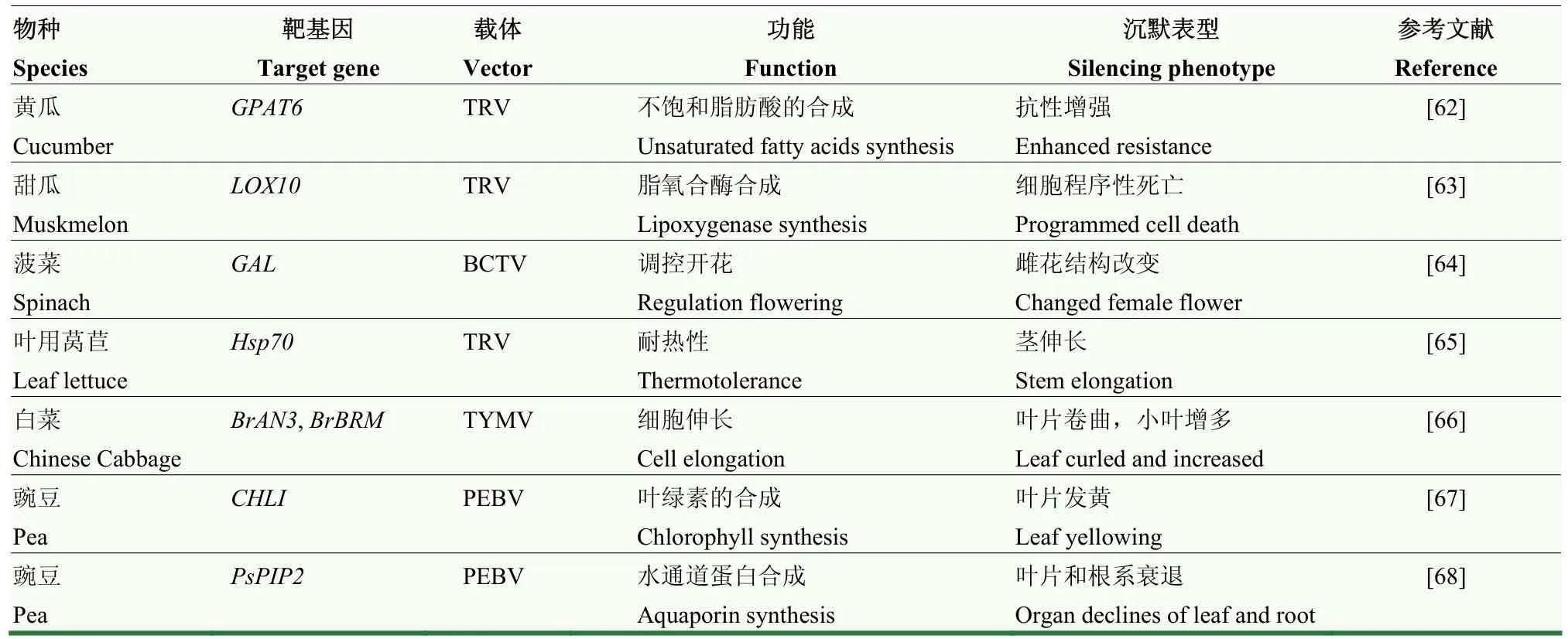
表2 利用VIGS研究的瓜类、叶菜类和豆类蔬菜基因及其表型Table 2 List of genes silenced by VIGS and the phenotype observed in melon, leaf and legume vegetable
芜菁黄化花叶病毒(turnip yellow mosaic virus,TYMV)具有正义 RNA链,可以侵染许多十字花科植物。例如以白菜(B.rapassp.pekinensiscv.Bre)BcPDS和BcNRT1为靶基因,从BcPDS和BcNRT1中选取合适的40 bp序列,反向互补处理后用T4连接酶进行连接,分别构建pTY-BcPDS和pTY-BcNRT1载体。白菜植株被 pTY-BcPDS侵染可导致白菜中该基因的表达显著降低,沉默效果比较明显;pTY-BcNRT1侵染白菜植株,显著抑制了BcNRT1在转录水平的表达[29]。最近的一项关于白菜包心紧实度受转录辅激活因子 ANGUSTIFOLIA3(AN3)的研究中,也利用TYMV-VIGS技术,发现基因沉默的白菜在莲座期和团棵期AN3都下调表达,证明基因沉默刺激了白菜叶片包心的形成[66]。
在菠菜雌花发育机制的研究中,通过甜菜曲顶病毒(beet curly top virus,BCTV)诱导的VIGS体系,对DELLA家族转录因子SpGAI在GA参与雌花形成中的基因功能得到了验证。沉默SpGAI后雌花表现出雄花器官的表型特征,中度表型是发育一个雄蕊代替雌蕊,但仍产生两个萼片;重度表型是发育4个萼片、1个雌蕊和1个雄蕊,同时花有4个萼片,类似于藜科植物中的完全花。沉默SpGAI后雄花发育成野生型雄花,但对外观表型没有影响[64]。MA 等[70]以甘蓝BoPDS为靶基因,利用CRISPR/Cas9系统实现甘蓝基因组的精准编辑,主要是依赖单链向导 RNA表达基因来完成突变体的构建,与VIGS技术相比,构建程序复杂,且目标基因在核苷酸预期位置容易缺失。在研究叶用莴苣热胁迫下Hsp70表达量和形态变化时,研究者构建pTRV-LsHsp702711沉默体系,发现未进行胁迫处理的沉默植株LsHsp70-2711表达量下降,茎明显伸长,热胁迫和干旱处理后的沉默植株LsHsp70-2711表达量显著低于对照植株,且高温胁迫对sHsp70-2711的影响大于干旱胁迫[65]。
CONSTANTIN等[71]利用豌豆早枯病毒(pea early browning virus,PEBV)研发了一套有效的VIGS体系,使得在豌豆中利用反向遗传学方法研究基因功能成为可能。在研究ROS、Ca2+参与豌豆叶绿素合成的研究中,利用VIGS技术构建了叶绿素合成关键基因CHLI沉默体系,采用组织化学和荧光染色试验发现沉默豌豆植株中ROS和胞间游离Ca2+增多,且豌豆发黄叶片中产生超氧阴离子和过氧化氢[67]。研究者还利用VIGS技术沉默了豌豆质膜水通道蛋白基因PsPIP2;1,发现沉默植株的叶片和根系中PsPIP2;1下调表达,从而证明豌豆叶片和根系水分运输中PsPIP2;1对水通道蛋白具有调节作用[68]。ZHANG等[72]利用菜豆荚斑驳病毒(bean pod mottle virus,BPMV)在大豆上成功建立了VIGS体系,且该病毒载体能够应用于大豆、菜豆等豆类蔬菜上,改造后拓展了BPMV病毒的宿主范围。
5 VIGS技术的不足之处和研究展望
尽管VIGS技术自被开发以来,已在许多蔬菜作物的基因功能鉴定研究中发挥了重要作用,同时建立了多种有效的VIGS载体,但是还存在一定的局限性。首先,沉默茄果类蔬菜后植株的沉默表型不均一、表型难以辨识,如在沉默番茄、辣椒等果实颜色相关基因时,表现出沉默植株表型不均一;再比如,利用ALSV病毒侵染的藜麦叶片摩擦接种到辣椒上并不能成功侵染,沉默植株表型不均一,但使用ALSV病毒浓缩液则侵染成功,且沉默效率达到90%。其次,VIGS技术存在靶基因沉默不完全、功能敲除不彻底、脱靶沉默和干扰症状等现象,如同时沉默番茄的两个基因SlEBF和SlEBF2后发现植株表现出败育和生长缓慢等一系列表型,影响了2个基因功能的研究;再者,这种瞬时沉默技术的稳定性和持续时间较差,且无法稳定遗传。另外,基因家族功能冗余情况也会因缺乏某一特定家族成员基因完整序列而导致干扰;此外,环境因素如温度、湿度和光照等可影响病毒在宿主植物中的扩散速度,其中温度对病毒粒子的扩散起主要作用。如 TRV病毒侵染番茄时,较好的沉默表型在22℃或者更低的温度下才能发生。应用VIGS技术的困扰因素还有农杆菌的保存期短、农杆菌活性以及扩繁效率低,因此,建立并优化农杆菌扩繁发酵技术与工艺也是今后努力的方向[73]。VIGS载体种类仍然有限,因此限制了VIGS技术的广泛应用[74]。
VIGS自被开发以来,已经开始应用于蔬菜作物病毒病的预防和控制、杂草的防控和蔬菜作物重要生长阶段的调控等领域,改变了传统的化学农药防治方法。例如,NEENA等[75]使用一种纳米材料 BioClay装载dsRNA并沉默同源RNA,以控制烟草病毒病的传播。目前,一些农业化工企业已经开始研发相关技术和产品[76]。未来会有相关产品取代传统的化学农药[77]。在优化VIGS技术方面,为避免脱靶现象,可利用不同种类的核酸数据库和在线软件精确设计靶基因序列,以提高靶基因沉默的准确度。此外,病毒载体可从RNA病毒、DNA病毒载体、卫星病毒等多种载体中选择适用于不同种类蔬菜作物的沉默载体。今后随着VIGS技术的不断发展,主要研究的方向可从以下几个方面进行研究:一是开发特异性与稳定性更高的病毒载体;二是优化病毒侵染后植株的培养条件,如光照、温度等方面进行系统研究,提高基因沉默的稳定性;三是果菜类蔬菜果实离体VIGS技术的研发,以便用于果实采后保鲜;四是提高VIGS诱导基因沉默的持续时间,从而实现在蔬菜长季节栽培过程中生长发育调控基因功能的研究;五是选择高效的基因片段进行沉默,以提高基因沉默的准确性。最后是病毒载体的安全性问题,病毒是作物的重要病原物,且大多数植物病毒都能以昆虫为媒介进行传播。所以,在构建VIGS病毒载体时应对病毒载体进行修饰,使其不引起严重症状或被介体传播,试验结束后对试验材料进行合理规范处理以免造成病毒传播。
RNAi能够特异性地降低或关闭靶基因表达,是基因功能研究的重要手段,但很难控制靶基因的沉默效率,且需要很长一段时间来建立遗传转化体系,大大增加了科研工作者的时间成本,降低了研究效率[78]。CRISPR/Cas9技术虽然具有一次编辑多个基因的功能优势,但存在脱靶效应,且对于多倍体植物而言,基因组改造的难度很大[79-80]。VIGS技术在选择目的基因靶位序列时,对插入片段的方向并无要求,且不易脱靶[81-82]。因此,随着高通量测序等新技术的不断研发和VIGS技术的优化,基于非编码的单链小分子RNA构建的VIGS技术,可以用于农作物新品种目标基因的高通量快速筛选,促进了蔬菜基因工程和蔬菜作物改良以及分子育种的发展,推进生产不携带外源基因的蔬菜品种育成的进程[83]。相信VIGS作为高效的研究基因功能的技术在蔬菜作物上的应用必将被逐步完善,在蔬菜功能基因组学研究的应用前景会更加广泛。
