天山乌鲁木齐河源1号冰川不同海拔可培养空气源酵母菌系统发育及多样性研究
2021-05-24周雅雯白莹莹阚泽宇倪永清
周雅雯, 白莹莹, 阚泽宇, 王 珍, 张 艳, 倪永清
(1.石河子大学生命科学学院,新疆 石河子 832000; 2.石河子大学食品学院,新疆 石河子 832000)
0 引言
地球表面近80%的生境被永久或定期暴露于低于5 ℃的寒冷生境中[1],包括深海、极地、冰盖、寒冷沙漠和高山冰川。其中冰川覆盖了地球陆地表面约10%[2],分布极为广泛,因其长期低温、寡营养、冻融循环、强辐射、低水活度等环境特点形成了独有的生境特征并成为制约其中微生物生存的重要限制因素[3]。全球范围下包括极地和非极地的山岳冰川在内的寒冷生境是低温微生物的重要来源,最适生长温度小于等于15 ℃、最高生长温度小于20 ℃,能够在0 ℃甚至更低温度生长的称为嗜冷微生物(psychrophiles);能够在0 ℃生长,但最适生长温度在20~30 ℃左右,最高生长温度在30~37 ℃的称为耐冷微生物(psychrotolerants)[4]。嗜冷菌和耐冷菌以高山冰川作为定植的栖息地,成为该生境中生物多样性重要组成部分,并在营养循环和有机物质矿化方面发挥着重要的作用[5]。然而,目前对于高山冰川的微生物研究主要在细菌多样性及细菌群落组成影响因素等方面,而对于冰川生态系统中酵母菌的多样性及分布格局报道较少[6]。
目前发现,冰川是以微生物为主导的低温生态系统,栖息着丰富多样的微生物类群,是低温极端微生物的良好栖息地[7]。冰川生境也是陆地生态系统中对于温度变化最为敏感的生态系统,常作为气候变化的敏感晴雨表[8]。全球变暖加速了高海拔的冰川退缩[9],因此生存于其中的低温微生物可被视为生物指示器来预测全球变暖趋势及规律。酵母菌作为低温真核生物的重要代表,发挥着极为重要的生态功能,尤其对寡营养生境冰川难降解有机物质的矿化作用和物质循环极为重要[10]。在积雪、冰芯、融水、冰尘、沉积物等不同生境中均有分布,是研究冰川生态系统微生物空间分布的良好对象。
研究表明,冰川表面的微生物主要来自于空气气溶胶,以及来自于陆地的尘埃,火山灰烬,以及粘附在雪粒、降雨、矿物质或有机物颗粒上的微生物,通过大气输送以及大气生物学过程沉降在冰川表面,此后随着冰川消融各种外源微生物进入冰川生态系统,发挥不同的生态功能[11]。因此,冰川表面空气中的微生物,影响着冰川各生境微生物群落和特殊种群的空间分布格局,为了解冰川微生物生物地理学分布提供重要的信息。但研究也发现,冰川表面生境空间异质性明显,包括积雪、融水、冰尘、冰层,对来自外源的微生物具有明显的生态选择作用[12]。
天山山脉横亘于亚洲中部,作为中亚地区最大的山脉,是全球水热循环的重要组成部分,深刻影响着亚洲以至全球的气候变化[13]。前期我们对天山乌鲁木齐河源1 号冰川(简称乌源1 号冰川)生境的真菌群落[14],冰川融水、积雪、冰心、沉积物的酵母菌的多样性及分布特征已做了相关报道[15]。本研究旨在揭示冰川空气源可培养酵母菌的多样性,并比较了乌源1号冰川表面不同海拔的位点之间酵母菌类群的异同,以了解冰川空气源酵母菌与其他生境之间的联系,为阐明冰川表面、底部低温酵母菌种群的原位栖息地及可能的生境扩散理论提供理论参考。同时揭示在冰川局域范围内,小幅度海拔梯度变化对空气酵母菌多样性的影响很小。研究将为了解冰川微生物来源,促进天山冰川微生物资源的开发和利用奠定基础提供数据支撑。
1 材料和方法
1.1 实验材料
1.1.1 样品采集
于2017年7月在乌源1号冰川西支尾部及表面分3个位点进行采样:低海拔位点(LA,43°07′125″N、86°48′707″E,海拔3 750 m,气温7.23 ℃)、中海拔位点(MA,43°06′54.09″N、86°48′40.07″E,海拔3 857 m,气 温6.18 ℃)、高 海 拔 位 点(HA,43°06′26.63″N、86°48′22.99″E,海拔4 013 m,气温4.67 ℃)。使用JH-6120 型综合大气采样器,TSP切割器标定流量:100 L·min-1,气体入口流速:0.3 m·s-1。微孔滤膜采用玻璃纤维素滤膜,直径90 mm,孔径0.22 μm,每张滤膜都累计8 小时的采样时间,空气样品采集完成后立即用酒精灼烧灭菌的镊子将滤膜装入已灭菌的无酶采样管,置于-4 ℃的车载冰箱中于6 小时内运回实验室,于24 小时内取样处理分菌。
1.1.2 培养基
为尽可能全面揭示空气中可培养酵母菌的多样性,本实验运用稀释涂平板法来分离培养酵母菌。此外还使用了4 种培养基分离酵母菌,所有培养基成分均采用常规用量的1/5,以此来模拟冰川的寡营养状态。所用抗生素为氯霉素(100 mg·mL-1)。
RDBC 培养基:Peptone 5 g·L-1,Glucose 10 g·L-1,KH2PO41 g·L-1,MgSO47H2O 0.5 g·L-1,Dichloran 0.002 g·L-1,Rose bengal 0.025 g·L-1,Agar15 g·L-1;DG18培养基:Glycerol 220 g·L-1,Peptone 5 g·L-1,Glucose 10 g·L-1,KH2PO41 g·L-1,Mg-SO4.7H2O 0.5 g·L-1,Dichloran 0.002 g·L-1,Agar 16 g·L-1;MYP 培养基:Malt extract 7 g·L-1,Glucose 1 g·L-1,Yeast extract 5 g·L-1,Soya peptone 2.5 g·L-1,Agar 15 g·L-1;MYP5 培养基:Malt extract 7 g·L-1,Glucose 1 g·L-1,Yeast extract 5 g·L-1,Soya peptone 2.5 g·L-1,Agar 15 g·L-1,Nacl 50 g·L-1。
1.2 酵母菌分离纯化
将采样空气滤膜置入100 mL灭菌生理盐水中,进行均质化。待混合均匀后,吸取定量上述液体于孔径0.45 μm、直径47 mm 的无菌硝化纤维膜过滤[10]。待过滤完成后将滤膜分别贴于四种分离培养基表面,RDBC 培养基、DG18 培养基、MYP 培养基、MYP5 培养基。将贴有滤膜的培养基置于16 ℃下培养7 d 后挑取单菌落,连续转接划线培养3 次后,记录菌落大小、颜色、透明度、边缘特征、湿润度、菌落凸起等特征,根据表型差异进行初步分离筛选并纯化,所得纯培养物转接YEPG(Yeast extract 10g·L-1;Peptone 10g·L-1;Glucose 20 g·L-1)斜面培养基4 ℃保藏备用。液体纯培养物离心后,加入新鲜培养液重悬,补充15%的灭菌甘油冷冻保藏在-80 ℃冰箱。
1.3 DNA提取和PCR扩增
对所有分离到的酵母菌进行DNA 提取,DNA的提取参照文献[16]的方法进行。DNA 提取后加入100 μL 1×TE 溶解,-20 ℃保存。采用超微量分光光度计定量核酸浓度和纯度。PCR 指纹图谱参照文献[17]进行。对具有相同DNA 带型的酵母菌进行分组,并初步认为其属于同一个种,将不同带型的代表菌株进行测序。
1.5 ITS 基因间隔序列的扩增及菌株系统发育分析
将扩增后MSP-PCR 条带对比去重后,挑取同一基因型中的代表菌株以ITS1f(5’-CTTGGTCATTTAGAGGAAGTAA-3’)和ITS4(5’-TCCTCCGCTTATTGATATGC-3’)为 引 物 进 行PCR 扩增。PCR 反应体系为25 μL(2×PCR Master Mix 12.5 μL;DNA模板2 μL;3 μmol·L-1的ITS1f:0.5 μL;0.2μmol·L-1的ITS4:0.5 μL;ddH2O 9.5 μL)。扩增程序:预变性94 ℃3 min;变性95 ℃30 s,退火55 ℃30 s,延伸72 ℃30 s;40 cycles,终延伸72 ℃6 min。将3 μL PCR 产物点样于2%浓度的凝胶电泳检测后将剩余的PCR 产物作为样品送至生物工程(上海)股份有限公司进行测序。
将所测的基因序列在NCBI 核酸序列数据库进行同源序列比对(BLAST),找出与所测序列相似度最高(97%~100%)的BLAST 库的亲缘标准菌株,CLUSTAL X 2.0 软件序列联配。进化距离采用邻接法neighbor-joining method 计算,在MEGA 7.0 软件[18]中用p-distances和Kimura-2parameter双参数法构建系统发育树,并用bootstrap 检验进化树分支聚类的置信度,重复1 000次[5]。
1.6 酵母菌生长温度测定
利用测序结果和NCBI 数据库比对筛选出代表菌株,将其活化后按2%的接种量接入5 mL YEPG液体培养基中,分别置于4、10、18、25、30、37 ℃共6个温度梯度下培养,培养96 h 后,使用紫外分光光度计于420 nm测OD值。
2 结果与分析
2.1 空气源酵母菌分离纯化与系统发育分析
利用4 种琼脂培养基在3 个空气采样位点分离到414 株酵母菌,从各培养基分离的酵母菌数量分别为:RB(219 株);DG18(43 株);MYP(117 株);MYP5(35株)。酵母菌的丰富度程度与海拔并无明显线性关系,其中135 株来自于低海拔位点(LA),176 株来自于中海拔位点(MA),103 株来自于高海拔位点(HA)。
本研究分离的所有酵母菌,皆为担子菌门,其中白色与乳白色的菌落分离物被视为无色素酵母菌共有258株,另有156株色素菌株。乌源1号冰川空气中分离纯化后的菌株依据菌落颜色,形态分类,菌落颜色丰富(图1),有纯白色、灰白色、粉白色、奶油色、乳黄色、浅黄色、粉红色、橘色,浅棕色等。酵母菌菌落直径约1.5~3.5 mm,小部分菌落直径小于1 mm,酵母菌的细胞形态通常有球形、近球形、卵圆形、椭圆形、圆柱形、棒形、藕节形、柠檬形等,一般为2~4 μm。

图1 乌源1号冰川空气源部分酵母菌代表菌株菌落图片(A:bca-86,Rhodotorula araucariae;B:bca-321,Naganishia adeliensis;C:bca-248,Filobasidium stepposum;D:bca-32,Filobasidium floriforme;E:bca-502,Dioszegia hungarica;F:bca-175,Rhodotorula glutinis)Fig.1 Morphological characteristics of colony of representative strains of some yeast species in the Urumqi Glaciers No.1,Tianshan Mountains(A:bca-86,Rhodotorula araucariae;B:bca-321,Naganishia adeliensis;C:bca-248,Filobasidium stepposum;D:bca-32,Filobasidium floriforme;E:bca-502,Dioszegia hungarica;F:bca-175,Rhodotorula glutinis)
根据菌落形态和生长特性对所有分离得到酵母菌株进行初步分组,并对形态和生长特征相类似的菌株划分为一组。扩增已分离纯化的低温酵母菌菌株的ITS 片段并测序,通过使用NCBI(National Center of Biotechnology Information)中的BLAST 比对工具在GenBank数据库中进行同源性比对。结果显示,从乌源1号冰川的表面空气中共分离414株酵母菌(表1),MSP-PCR 指纹分型划分为53 个基因型,从每个基因型中选出2或3株具有代表性的菌株进行ITS-rRNA 基因序列分析,共测序139 株菌株。测序结果显示分别隶属于9属28种,同时选取28株代表菌株的ITS-rRNA序列构建系统发育树(图2)。
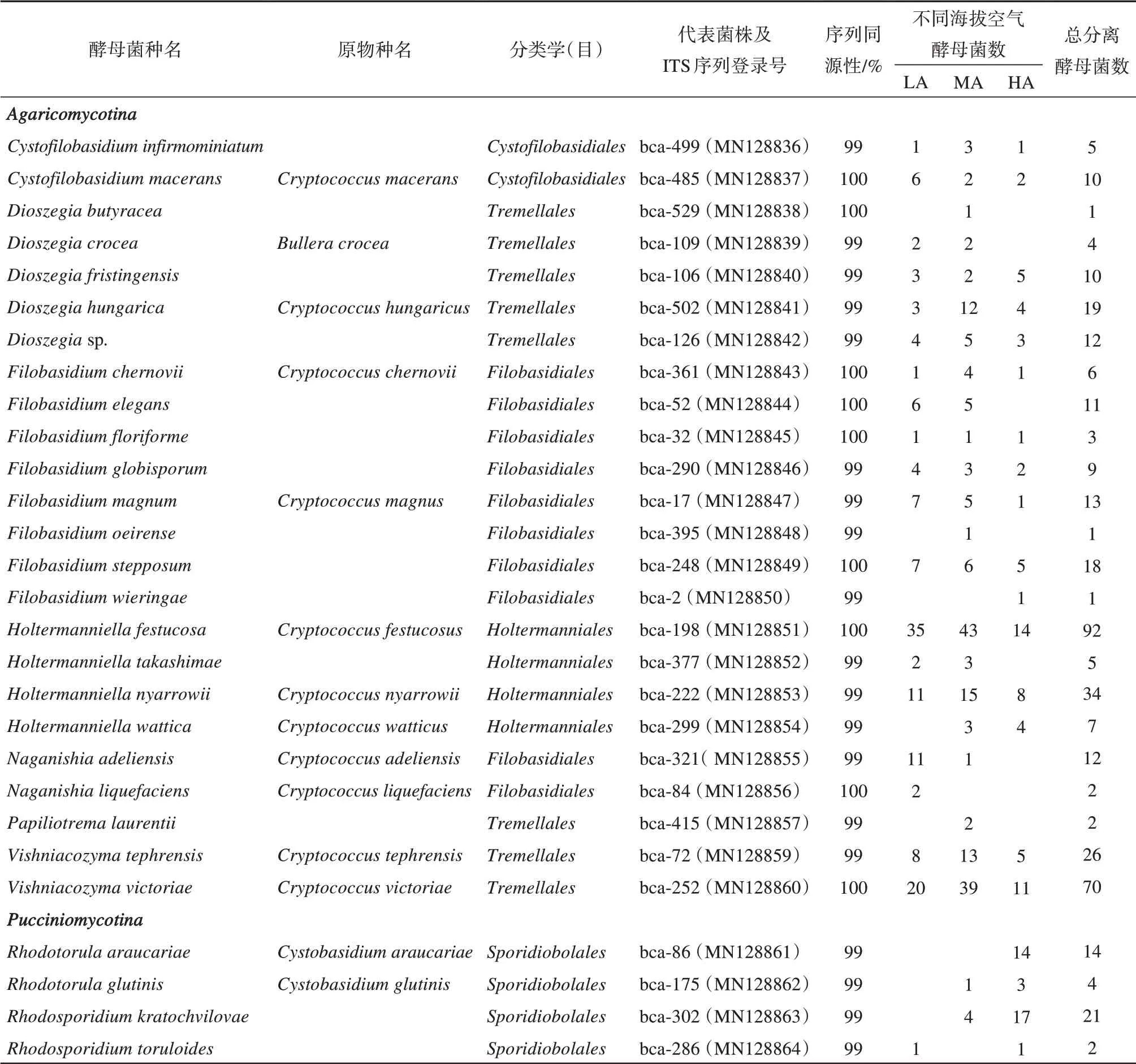
表1 乌源1号冰川空气源酵母菌种的分布及数量Table 1 Distribution of yeast species of airborne in the Urumqi Glaciers No.1,Tianshan Mountains

图2 基于ITS rRNA基因序列的乌源1号冰川空气源酵母菌系统发育树Fig.2 Neighbour-joining tree showing the phylogenetic relationships among yeast strains,taken from the Urumqi Glacier No.1,based on ITS rRNA gene partial sequences and related sequences downloaded from GenBank(The numbers at the nodes indicating the bootstrap values based on neighbor-joining analyses of 1 000 resampled datasets;bar 0.02 at the bottom is the sequence divergence)
对空气源酵母代表菌株进行ITS 序列分析(图2),有伞菌亚门24 株,柄锈菌亚门4 株。其中bca-2与Filobasidium wieringae;bca-72 与Vishniacozyma tephrensis;bca-529 与Dioszegia butyrace;bca-502 与D.hungarica;同源性高达99%亲缘关系最近。bca-198、bca-222、bca-377、bca-299 分属于同一个进化支,与Holtermanniella同源性最高,相似度为100%;bca-286、bca-175、bca-86、bca-302 分属于同一个进化支,与Rhodotorula同源性最高,相似度为100%;bca-106、bca-529、bca-109、bca-502 聚为一类,与Dioszegia亲缘关系最近;菌株bca-415 与Papiliotrema laurentiiCBS:5297 相似度为100%。分离自HA 位点的菌株bca-2 与模式菌株CBS11580(GenBank 登录号KY103449)Filobasidium wieringae有99%的相似性。分离自MA 位点的菌株bca-529 与模式菌株CBS10122(KY103348)Dioszegia butyracea有99%的相似性。而来自MA 位点的菌株bca-84与模式菌株CBS968(AF444345)Naganishia liquefaciens有97%的相似性。种水平的系统发育分析如图2所示,显示了我们所分离菌株与GenBank 库中登记的最接近的CBS 酵母菌菌种库的模式菌株的亲缘关系。
2.2 不同海拔空气源酵母菌群落分布特征
乌源1 号冰川表面空气中9 个属酵母菌种群结构显示,Filobasidium种类最多分离到8 个种,Dioszegia分离到5个种,Holtermanniella分离到4个种,Cystofilobasidium、Naganishia、Vishniacozyma、Rhodotorula、Rhodosporidium均分离到2 个种,Papiliotrema种类最少,仅分离到一个种。本研究依据每种酵母菌占总分离酵母菌数量的比例确定其对应的相对丰度。酵母菌属水平结果显示(图3),Holtermanniella相对丰度最高为33.33%,为最优势属,其 次 为Vishniacozyma(23.19%)和Filobasidium(14.98%)。除了Rhodotorula和Naganishia以外,其他属酵母在3 个海拔位点均有分布。随海拔升高,Holtermanniella,Vishniacozyma与Dioszegia相对丰度出现先升高再降低趋势;而Filobasidium,Naganishia与Cystofilobasidium相对丰度呈持续降低趋势;Rhodotorula相对丰度呈持续升高趋势。

图3 不同海拨冰川空气酵母菌属水平的群落组成Fig.3 Community composition of airborne yeasts at genus level in different altitude of the Urumqi Glacier No.1
酵母菌种水平结果显示(图4),Holtermanniella festucosa的相对丰度最高(22.22%),其次为Vishniacozyma victoriae(16.91%),而Holtermanniella nyarrowii、Vishniacozyma tephrensis和Rhodosporidium kratochvilovae相对丰度也比较高,均超过了5%。Dioszegia crocea、Rhodosporidium kratochvilovae等菌株丰度均低于1%。综合分析相对丰度与发生率(每个酵母菌种在每个地理位点的出现频率),乌源1号冰川表面空气酵母菌优势种为Holtermanniella festucosa,其占LA 样品总菌株数的25.93%,占MA 样品总菌数的24.43%,占HA 样品总菌数的13.59%。
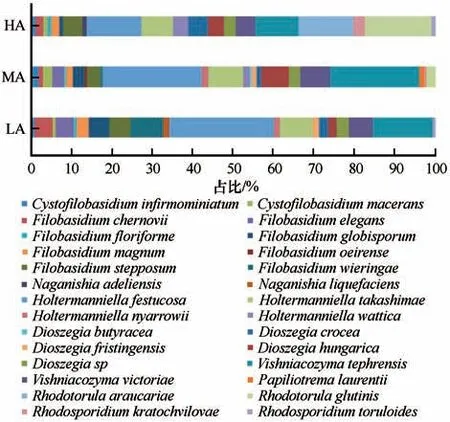
图4 不同海拨冰川空气酵母菌种水平的群落结构组成Fig.4 Community composition of airborne yeasts at species level in different altitude of the Urumqi Glacier No.1
2.3 空气源酵母菌最适生长温度
实验结果表明,30 株测试的代表菌株中,22 株菌最适生长温度为22~25 ℃,在30~37 ℃缓慢生长;6 株菌生长温度范围在4~22 ℃,其中最适生长温度在16~18 ℃。根据一般定义[4],前者属于耐冷菌属,后者属于嗜冷菌,这说明乌源1 号冰川空气酵母菌大部分属于耐冷菌(表2)。
2.4 酵母菌群落组成及多样性分析
总体而言,乌源1 号冰川表面3 个不同位点的空气样品中,酵母菌丰度分布存在小幅度的差异。从3 个海拔分离出Holtermanniella属酵母菌,占总分离物的33.33%,其次是Vishniacozyma属酵母菌,在分离物总数中占比23.19%。根据最新的分类系统,本研究在不同海拔位点空气分离菌株共有7 个属,除Naganishia出现在2 个海拔和Papiliotrema只在1 个海拔以外,其余各属在3 个海拔均有分布(图5)。

图5 乌源1号冰川空气源酵母菌种水平生境组合强度Fig.5 Horizontal habitat assemblage intensity of glacial airborne yeast species in the the Urumqi Glacier No.1(HA,LA,and MA representing three sample collection sites,respectively).Note:the bars representing the cumulative relative abundance of each indicator yeast species in all the samples;the size of each circle defining the association strength(indicator value)of a yeast species with the different habitats,such as 0~10:not characteristic;10~20:weakly characteristic;20~30:characteristic;30~40:strongly characteristic
ACE 指 数Chao 指 数 可 反 映 物 种 丰 富 度[19],Chao 指数代表物种丰富度,对稀有物种变化敏感。由表3可知,随着海拔小幅度增加,各样点酵母菌群落丰富度指数(ACE 指数)与(Chao 指数)均显示为MA>HA>LA。在海拔梯度上,ACE 指数和Chao 指数呈先上升后下降的趋势,在MA位点达到做大值,分别为28.26 和27.67,说明空气源酵母菌在MA 时物种最为丰富,但统计结果显示差异不显著。在海拔上升的过程中,Chao 指数的增加幅度较大,说明稀有菌株出现得更多。Shannon 指数代表了样本中物种的均匀度,随海拔梯度上升,各样点酵母菌群落多样性指数(Shannon 指数)呈略下降趋势,但差异仍然不显著。

表2 乌源1号冰川空气源酵母菌形态特征及生长特性Table 2 Growth characteristics and morphological characteristics of airborne yeasts in the Urumqi Glaciers No.1,Tianshan Mountains
3 讨论
无论在极地还是高山冰川的生态系统中,低温酵母菌都代表了冰川微生物群落的一个重要组成部分[20]。目前的报道显示,在两极地区,多年冻土、高山冰川、深海以及寒冷沙漠等低温环境中,可培养酵母菌主要为担子菌门和子囊菌门两大类[21],但研究显示,担子菌门酵母菌比子囊菌门酵母菌有更好的低温生态适应性[22]。由于不同地区生态环境的差异,不同生态系统中低温酵母菌类群差异显著。本研究结果表明,在门水平上的空气源酵母菌中的优势菌门全部为担子菌门。
3.1 冰川生境酵母菌种群与空气酵母菌种群的关系

表3 乌源1号冰川不同位点空气酵母菌群落丰富度和多样性指数Table 3 Abundance and diversity of airborne yeasts communities of different sites in the Urumqi Glaciers No.1,Tianshan Mountains

表4 乌源1号冰川空气酵母菌与其他4种生境中酵母菌的比较Table 4 Distribution of yeast species of airborne(TA)and other habitats of the Urumqi Glacier No.1,Tianshan Mountains,including supraglacial ice(BB),cryoconite sediment(BC),supraglacial meltwater(BW)and supraglacial snow(SN)
将本研究结果与课题组前期对乌源1号冰川中冰尘、表面融水、积雪和冰芯共五种生境的酵母菌组成进行比较研究(表4),冰川五种生境中担子菌酵母均占明显优势,这是多数冰川生态系统的共同特征[23-24]。很多研究报道显示,原属于隐球菌属(Cryptococcus)的酵母菌Vishniacozyma属,其中的Vishniacozyma victoriae与V.tephrensis在5 种 冰 川生境均有分布(表4)。然而,文献报道显示,在地球其他生境这两个种也有分布,譬如植被及果蔬表面。我们的研究发现,乌源1号冰川空气中Vishnia-cozyma victoriae与V.tephrensis为优势菌,很可能是通过空气传播将植物体表的菌株携带到了冰川生境。但目前对不同生境种群之间的进化关系没有进行深入报道,不同菌株之间ITS 基因序列显示了很高的同源性,因此有可能属于世界广布种,也有可能属于同种的不同生态型。从表4 可知,乌源1号冰川不同生境广泛分布红酵母属(Rhodotorula)菌株,具有很高的分离频率,表明在低温生境具有很强的适应性[25]。其原因可能是Rhodotorula属酵母菌可产生类胡萝卜素,作为光保护剂能够有效避免紫外线对细胞的损伤。此外,类胡萝卜素是一类膜保护的抗氧化色素,可抵抗UV 并增强细胞膜的流动性,能够有效清除自由基[26]。此外,一般认为该属还能够产生胞外多糖,使它们在冰川生境中具有更强的生态胁迫耐受性[27]。实际上,在空气、叶面和腐烂的植物碎片中均能分离到它们,其中Rhodotorula araucariae在腐败的植物体表很容易分离到[28]。因此,以Rhodotorula属酵母菌为模式菌,研究在极端环境条件下,紫外线辐射在对酵母菌菌群结构具有重要的生态学意义。同样,Rhodotorula属在乌源1 号冰川空气中也存在,也可能是通过空气传播将植物体表的菌株携带到了冰川生境。但目前对红酵母属的系统发育和分类做了大的修改,也有可能是隶属于不同种的酵母在不同生境的分布需要做深入研究[26]。
鉴于以上情况,人们很容易认为冰川生境的酵母菌与空气酵母菌密切相关。但本研究一个例外的发现是空气源的优势酵母属Holtermanniella,它们在冰川中很少被分离到[13],这说明冰川对外源微生物具有选择作用,即Holtermanniella属作为外源种群不适应冰川生境[13]。表4显示,Holtermanniella属中H.festucosa、H.takashimae、H.nyarrowii、H.lawattica在冰川的冰尘,融水,雪水,冰芯中均未被分离到,但在空气中属于优势种群。表明冰川生态系统对外源微生物具有显著的生境过滤作用,只有适应性很强的种群才能进入冰川生态位生殖繁衍,进而通过微进化发生生态生理渐变[29]。因此,我们预期在冰川生境景观尺度下,极端环境因素(包括紫外线、温度、寡营养环境等)对微生物的群落构建(群落结构和物种分布格局)和空间分布格局产生显著的影响[30]。以酵母菌作为范式,研究冰川生境的群落构建过程中,由非生物因素与生物因素的相互作用,及其对微生物群落动态格局贡献度很有必要。
3.2 空气酵母菌群落结构特点
众所周知,冰川区域在空间地理上环境多变,尤其在垂直尺度上环境因子相差较大,形成了不同的小生境,蕴含了丰富的微生物类群。乌源1 号冰川表面空气微生物可能来源于内部和外部环境,其中来自周边生境的冰川低温酵母属于原位栖息的本地种(endemic species),由于大气环流随机迁移的属于陆源性外来种(exotic species)。研究发现Naganishia adeliensis在不仅在空气中分离到,在冰川多个生境均有发现,表明该属种类大多为本地菌[13]。Cystofilobasidium infirmominiatum、Filobasid-ium chernovii、Filobasidium elegans、Filobasidium wieringae主要在空气与冰川表面的积雪生境所分离到,很有可能是从积雪表层传递到空气。第二类是近距离外来菌(Local exotic species),这些菌株很可能主要来自于冰川前沿退缩地的植物花蜜和植物源附生酵母菌。如Rhodotorula属,在空气、叶面和腐烂的植物碎片中均能分离到它们,R.araucariae、R.glutinis、R.kratochvilovae、R.toruloides是空气与冰川表面冰所共有的菌种[27]。第三类是远距离外来菌(Distant exotic species),通过大气环流运动,通过降水(雨、雪),水凝华(云、雾、冰雹)和灰尘沉降等被携带[31-32],这些种类可能来自空间地理差异较大的生态系统,如沙漠[33]、森林[34]等。有报道发现冰川的冰芯中分离出了来自于温带和热带地区的微生物[35]。Holtermanniella属的菌株大多分离自植物体表,在本研究中从冰川生境没有分离到它们,很可能来自温带森林。此外Vishniacozyma victoriae是极地环境的优势种类,也是本研究中的优势菌株,但在非极地环境中普遍分离出该种亲缘关系相近的菌株,表明是全球的广布酵母菌种类[36]。
3.3 不同海拔梯度下冰川表面空气源酵母菌群落构建机制
本研究位点LA 选取在冰川前沿高山草甸处,以高山垫状植被为主,此处植物群系开始多元化且动物出没频繁,受动植物及人为干扰因素大。位点MA 选取在冰川前沿退缩处,此处均为裸露及风化的岩石,有动物活动。位点HA 选取在冰川冰舌中部,此处周围即冰川腹地,下方为冰雪及冰尘洞,受外界因素干扰较小。在冰川表面成功定植的微生物需要耐受大范围幅度剧烈的温度波动,以及抵御太阳辐射,并耐受寡营养等胁迫条件[37-38],在大幅度海拔梯度上呈现出了不同的生态位分布格局[39]。但基于本研究的结果,3 个梯度海拔的可培养酵母菌多样性存在差异,很可能是每个位点采集空气样本时,部分环境因素(如风速和颗粒物质量浓度)导致空气采集有一定程度的差异。对于空气源酵母菌来说,由于空气的空间异质性相对较小,在小梯度的海拔梯度下以连续的状态处于动态变化,可培养酵母菌差异并不显著。冰川生境受大气环流及周边人类和动物活动影响,内源、外源酵母菌分辨需要更精确的方法甄别[13],因此,对冰川表面和空气微生物关系需要深入研究。
4 结论
基于乌源1 号冰川表面3 个海拔高度的空气酵母菌群落结构分析及其与冰川表面生境酵母菌的比较,得到结论如下:
(1)分类特征:乌源1号冰川表面空气分离到的酵母菌全部隶属于担子菌门(Basidiomycota),包括9属28种,伞菌亚门(Agariomycotina)成员占绝对优势。Holtermanniella festucosa与Vishniacozyma victoriae为冰川空气广布的优势酵母菌种。酵母菌大多为来自周边生境的耐冷酵母菌。
(2)分布特征:在小幅度的海拔梯度上,空气酵母菌群落结构有所不同,但多样性指数没有显著差异。基于空气酵母菌和冰川酵母菌种群的比较,冰川生境酵母菌与空气酵母菌密切相关,但冰川生境对外源酵母菌具有明显的生境过滤与选择作用。
