长湖浮游动物群落结构特征及其季节变化
2021-05-24龚森森吴嘉伟罗静波谭凤霞杨德国何勇凤
龚森森,吴嘉伟,柴 毅,罗静波,谭凤霞,杨德国,何勇凤
( 1.长江大学 动物科学学院,湖北 荆州 434025; 2.长江大学, 湿地生态与农业利用教育部工程研究中心,湖北 荆州 434025; 3.中国水产科学研究院 长江水产研究所, 农业农村部淡水生物多样性保护重点实验室,湖北 武汉 430223 )
浮游动物是水域生态系统中的初级消费者,也是高营养级水生动物幼体的主要开口饵料以及动物性滤食性鱼类的主要食物,能有效调控浮游植物数量,降低水体初级生产力,在水域生态系统的结构功能和生物生产力的研究中具有重要意义[1-3]。浮游动物通常被认为是水质评价的重要指示生物,其种类组成、优势种、丰度及生物量等群落结构特征可以反映出水环境变化状况,预测其变化趋势[4]。目前,浮游动物已被广泛应用于湖泊、水库等内陆水域次级生产力的评估以及水环境的监测和评价,对水域生态系统的可持续发展利用具有重要意义[5-6]。
长湖是湖北省第三大天然淡水湖泊,是长江中游地区重要湿地生态系统,2010年被批准建立国家级鲌类种质资源自然保护区,具有蓄洪、灌溉、养殖和航运等综合功能[7-8]。近年来,引江入汉、蒙华铁路和318国道长湖特大桥等涉水工程的陆续建设对该水域产生不同程度的影响。多种评价方法综合分析表明,自2007年以来,长湖水质状况一直处于中度—重度富营养化状态[9-10]。目前围绕长湖的研究主要涉及浮游植物、水质监测、高等水生植物以及鱼类资源调查等方面[11-13],其中关于浮游动物的研究仅见于2012年[14]。笔者阐明浮游动物季节演替规律并探讨与环境因子的关系,为长湖水生生物资源的可持续发展与利用提供重要参考,并为下一步生态系统的保护、恢复与修复提供科学参考依据。
1 研究方法
1.1 区域概况及采样点的设置
1.1.1 区域概况
长湖位于长江和汉江之间,地跨荆州、荆门和潜江,属于北亚热带湿润季风气候,年降雨量约1200 mm,平均水深2.1 m[15]。长湖自西向东可分为庙湖区、海子湖区、马洪台湖区和圆心湖区4个区域(图1)。“引江入汉”工程使庙湖区被2个堤坝(图1实线隔断)拦截成静水区域,故未在此区域设置采样点。海子湖区狭长,分布有多个进出水口,马洪台区和圆心湖区均为开阔水域,其中圆心湖区有较多的湖汊和湖湾[16]。
1.1.2 采样点设置
依据《水域生态系统观测规范》[17],共设置14个采样点:海子湖区4个(H1~H4),马洪台区5个(M1~M5),圆心湖区5个(Y1~Y5),采样点坐标见图1。2015年4月(春)、7月(夏)、11月(秋)及2016年1月(冬)各采样1次。
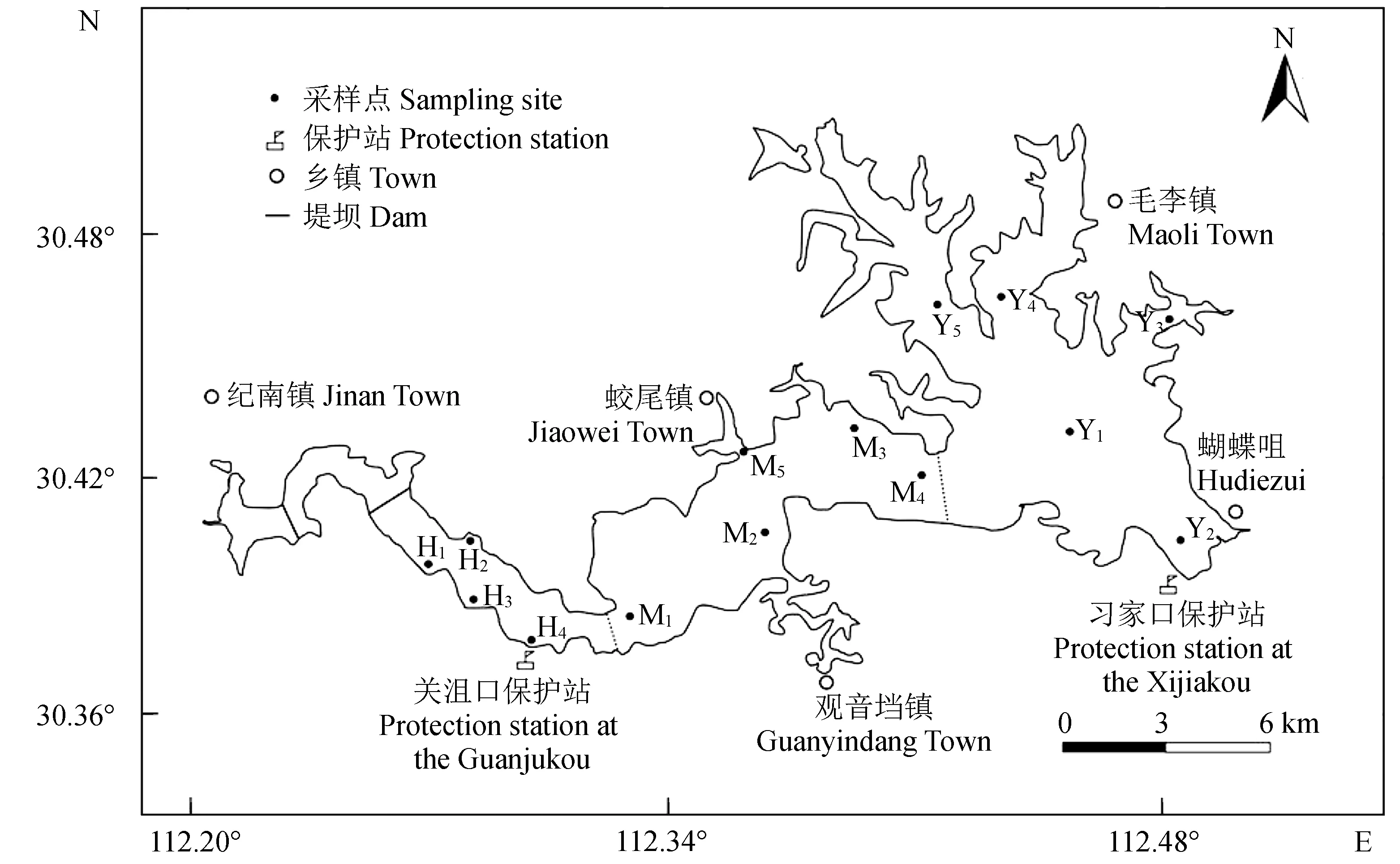
图1 长湖采样点示意Fig.1 Sketch map of sampling sites in Changhu Lake
1.2 样品的采集、处理及分析
1.2.1 样品采集与处理
参考《水域生态系统观测规范》和《水生生物学》[18]中样品采集方法采集浮游生物样本。参照《中国淡水轮虫志》[19]、《中国常见淡水枝角类检索》[20]、《中国动物志》[21]、《淡水微型生物图谱》[22]鉴定种类;定量计数采用多片计数取平均值的方法,生物量为固定样本运用体积法[18]得出的湿质量(mg/L)。
1.2.2 水质理化因子
按照国家水质标准方法GB 3838—2002,利用美国HQ40D(Hach Loveland)多功能水质分析仪测定并记录各采样点溶解氧含量、pH和水温,用黑白盘测定透明度,水样带回实验室测定总磷、总氮、氨氮、硝态氮、亚硝态氮、磷酸盐和总悬浮物含量等水质参数。
1.3 数据处理
1.3.1 优势度
用优势度指数(Y)确定优势种,Y>0.02即为优势种。
Y=(ni/N)×fi
(1)
式中,ni为浮游动物群落中第i种物种的丰度,N是群落中物种的总丰度,fi为第i种浮游动物在各站位的出现频率[23]。
1.3.2 物种更替率
物种更替率(R)按下式计算:
(2)
式中,a和b分别表示相邻两个季节的物种的种类数,c为两个季节共有物种的种类数[24]。
1.3.3 排序及分析
排序及分析运用R 3.5.3软件的Rstudio窗口完成。对浮游动物优势种与环境因子进行冗余分析,对各季节不同站位的生物量、丰度及物种数等生物指标进行非度量多维尺度分析,对优势种的各项生物指标进行聚类分析。参考《数量生态学—R语言的应用》[25]使用R语言软件。
2 结 果
2.1 浮游动物种类组成
2.1.1 种类组成
本次调查共鉴定出浮游动物3门34属61种(表1)。其中轮虫13属29种(占总数的47.54%),原生动物9属15种(占比24.59%),枝角类7属11种(占比18.03%),桡足类6属6种(占比9.84%)。夏季种类数最多,为43种,其次是春季,为37种,秋、冬季节分别为20种和19种。

表1 长湖浮游动物种类组成Tab.1 The species composition of zooplankton in Changhu Lake
2.1.2 丰度和生物量
各类群浮游动物在各区域的丰度变化见图2,其变化特征为夏(6781 个/L)>秋(2912 个/L)>春(1944 个/L)>冬(1359 个/L),季节性差异显著(单因素方差分析,F=17.910,P<0.001)。其中轮虫占绝对优势,最高达89.12%(夏季)。从各区域丰度分布来看,海子湖区浮游动物丰度在各季节均最高。
各季节平均生物量整体表现为夏(17.78 mg/L)>秋(15.45 mg/L)>春(5.07 mg/L)>冬(2.34 mg/L),季节性差异显著(单因素方差分析,F=8.443,P<0.001)(图3)。其中轮虫生物量在春、夏、秋3个季节当中均为最高,最高达13.99 mg/L(秋季)。从各区域分布来看,海子湖区平均生物量在春、夏、冬3个季节中均为最高。
2.2 优势种
利用优势度指数共筛选出12种浮游动物优势种(表2),包括8种轮虫和4种原生动物,其中萼花臂尾轮虫和针簇多枝轮虫为4季共有优势种,二者在各采样点的出现频率均为100%;由优势度指数可知,针簇多枝轮虫在4个季节的优势度均为最高,其中冬季达0.657。
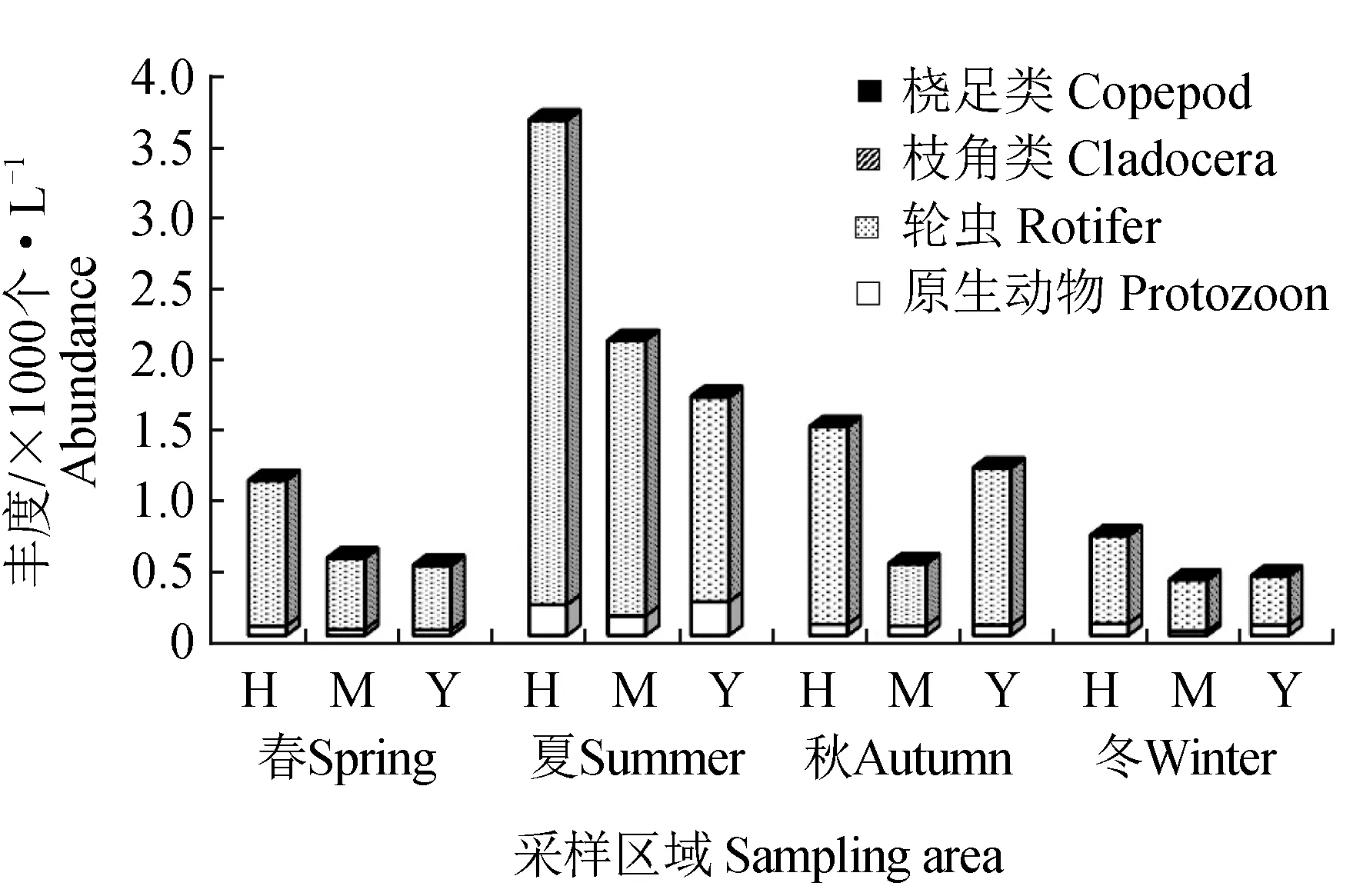
图2 各采样区域浮游动物丰度的季节变化Fig.2 Seasonal changes in zooplankton abundance in sampling sitesH.海子湖区域; M.马洪台区域; Y.圆心湖区域; 下同. H.Haizi Lake area; M.Mahongtai area; Y.Yuanxin Lake area;et sequentia.
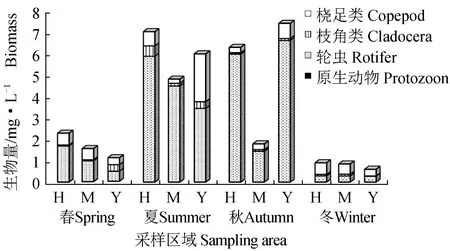
图3 各采样区域浮游动物生物量的季节变化Fig.3 Seasonal changes in zooplankton biomass in sampling sites in various seasons
2.3 物种更替率
统计相邻2个季节的总物种和共有物种,利用物种更替率公式计算出各类群物种更替率(表3)。总体来看,平均更替率呈现桡足类(80.01%)>轮虫(75.18%)>原生动物(74.66%)>枝角类(46.88%)。其中原生动物秋—冬季更替率最高(90.00%),枝角类冬—春季更替率最低(25.00%)。
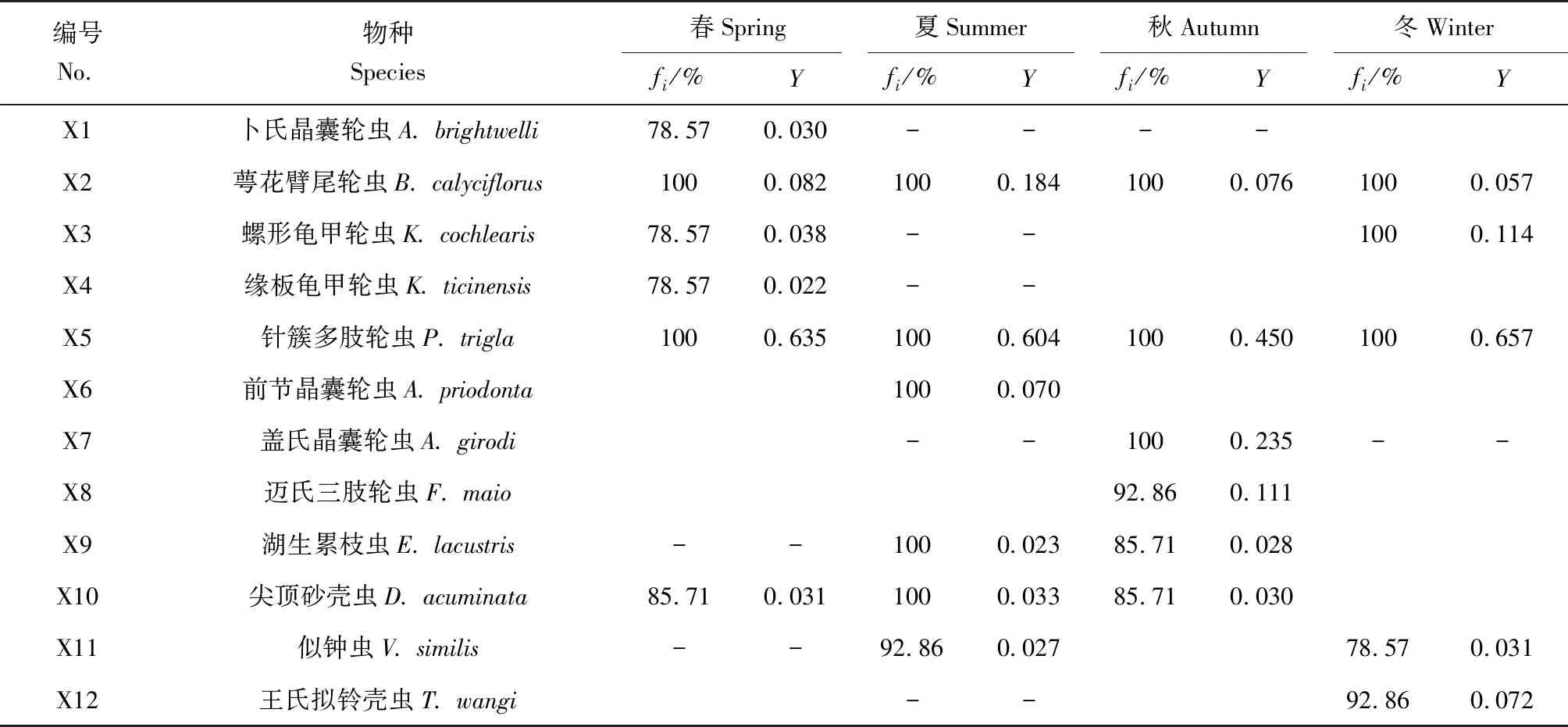
表2 长湖浮游动物优势种Tab.1 Dominant species of zooplankton in Changhu Lake
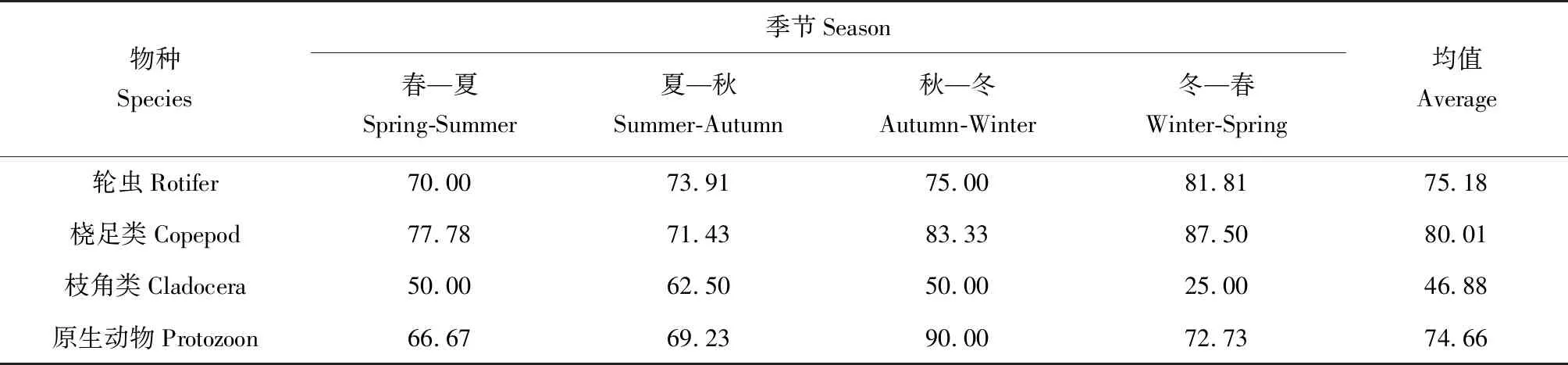
表3 长湖各类群物种更替率季节性变化 %Tab.2 The seasonal variation in species succession rate of various zooplanktons in Changhu Lake
2.4 相似性分析
以4个季节采样点为分类单元,以各样点在各季节的浮游动物丰度及生物量为依据进行非度量多维尺度分析,排序分析的平均胁强系数(Stress)为0.092∈(0.05,0.10),表明其结果可信[26]。各季节不同采样点的浮游动物丰度及生物量聚类分布差异明显,较好地表现出其季节性差异;其中各分类单元在冬季很好地聚为一类,与其他3个季节有明显的差异,而春季分类单元的分布较零散,且与夏、秋季个别分类单元表现出一定的相似性(图4)。
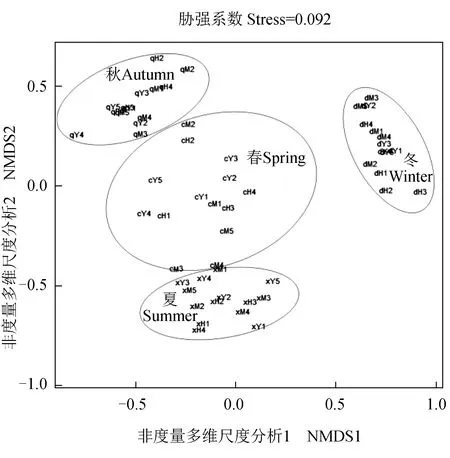
图4 长湖各样点非度量多维尺度分析Fig.4 Non-metric multi-dimensional scale analysis of various sampling site in Changhu Lake
以所有优势种为分类单元进行层次聚类分析。先计算Bray-curtis相异系数(取值0~1,越接近于1表明样本间浮游动物群落组成差异越大),再计算同表型距离及Gower距离对聚类方法进行筛选。结果显示,算术平均的非权重成对组聚类法最优;利用轮廓宽度图对最佳聚类簇数进行筛选,最终将优势种聚类树划分为4个簇,划分簇的Height值约为0.81(图5)。聚类结果表明,各分类簇中卜氏晶囊轮虫和缘板龟甲轮虫的时空分布特征最为相似,萼花臂尾轮虫和针簇多肢轮虫次之。
2.5 相关性分析
选取各季节水环境因子与对应季节优势种进行相关性排序分析。先将各季节优势种的原始统计数进行去趋势对应分析,结果显示,4个季节排序前4个轴中最大值依次是1.07、0.64、1.43和0.85,均小于3.0,选做冗余分析。再将各季节水质因子与优势种统计数经lg (X+1)转换(pH除外)后进行冗余分析,获得4个季节环境因子与优势种的相关双序图(图6)。图中箭头表示环境因子,“Δ”表示优势种,箭头连线的长短以及物种到原点连线与环境因子夹角的余弦值表示物种分布与环境因子相关性的大小。
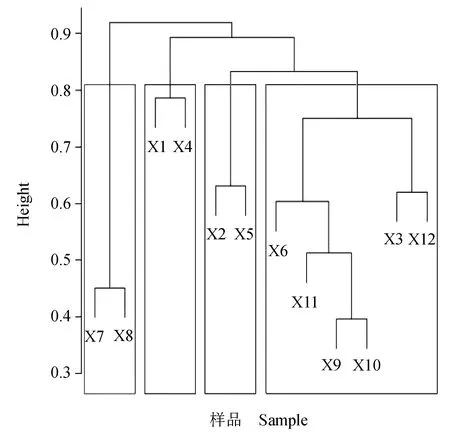
图5 长湖浮游动物优势种聚类分析Fig.5 Cluster analysis of dominant species of zooplankton in Changhu Lake X1~X12.优势种代码(表2). X1—X12.dominant species code(Tab.2).
各类群浮游动物优势种与环境因子表现出不同程度的相关性。夏季原生动物中湖生累枝虫、尖顶砂壳虫和似钟虫均与总氮含量呈较明显的正相关性。春季萼花臂尾轮虫与硝态氮含量和透明度呈正相关关系,针簇多肢轮虫与磷酸盐含量也表现出正相关,夏季萼花臂尾轮虫与总氮以及针簇多肢轮虫与总磷均有较明显的正相关,秋季盖氏晶囊轮虫与pH和水温、萼花臂尾轮虫和针簇多肢轮虫与总磷含量也呈正相关。冬季萼花臂尾轮虫、螺形龟甲轮虫、针簇多肢轮虫与溶解氧含量呈显著正相关。
3 讨 论
3.1 浮游动物群落结构特征
作为水域生态系统食物链中的初级消费者,浮游动物同时受到生产者(主要是浮游植物)和次级消费者[主要是水生动物幼体和鳙鱼(Aristichthysnobilis)等动物性滤食性鱼类]的影响,其中浮游植物的种类组成、丰度和生物量等特征可直接影响浮游动物的组成和分布等群落特征。在本研究中,浮游动物的种类组成为轮虫占比最高,总种类数为夏季最多。与同期浮游植物调查结果[27]比较发现,长湖属于典型的以绿藻门(占比50.8%)为主的水域类型,且种类数量也是夏季最多,这表明长湖浮游植物和浮游动物具有相似的季节分布和变化规律。还有研究表明,轮虫分布广,环境适应性强,尤其在富营养化浅水湖泊中,原生动物和轮虫通常是浮游动物的主要组成部分[28]。本次调查中原生动物和轮虫占比超过70%,占据了浮游动物的主要组成部分,尤其是轮虫的丰度和生物量均占绝对优势,符合轮虫在富营养化水域环境中的分布规律。
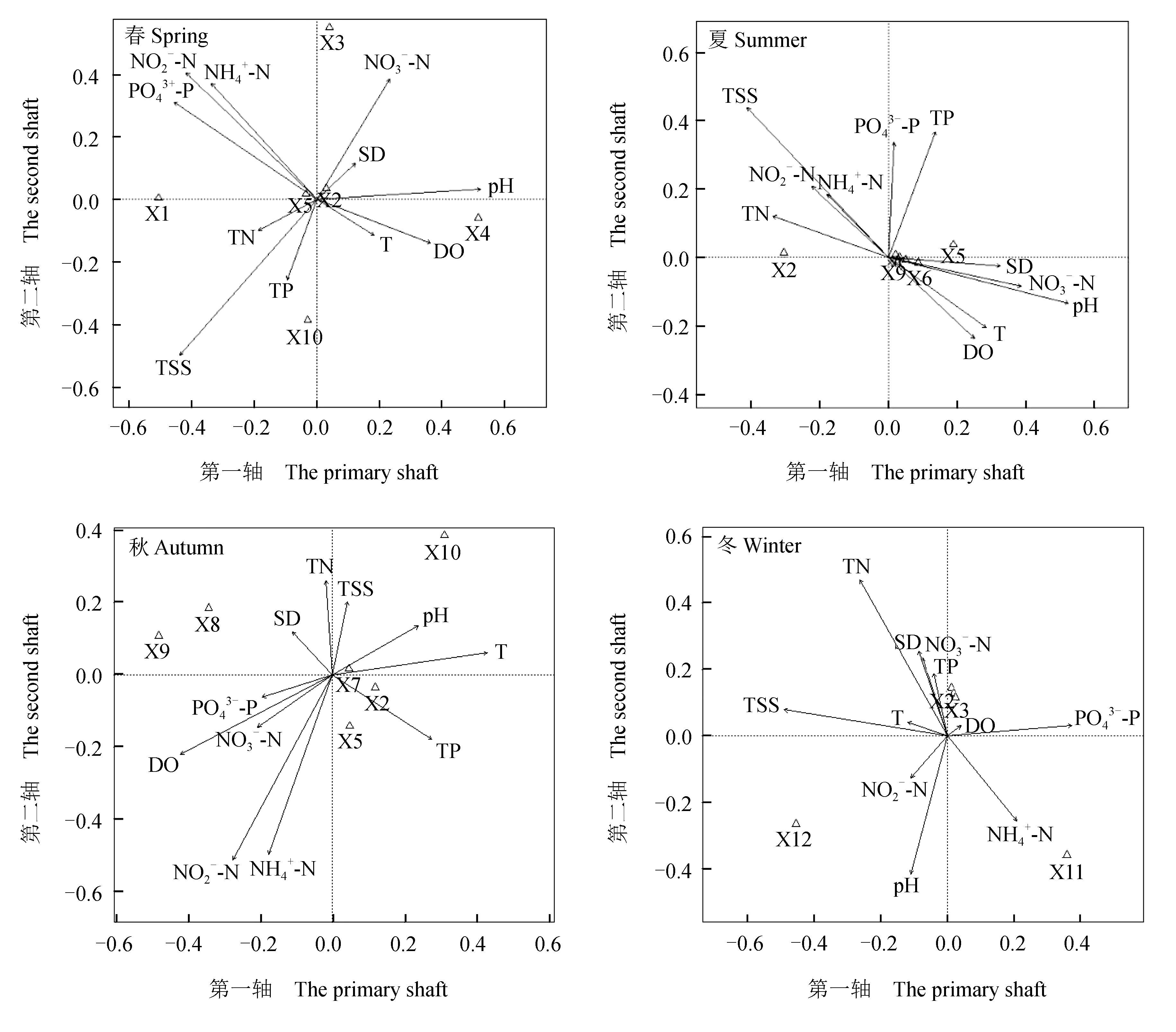
图6 各季节长湖内浮游动物优势种与对应环境因子的冗余排序Fig.6 RDA sequencing diagram between dominant species of zooplankton and corresponding environmental factors in Changhu Lake in each season DO.溶解氧; T.水温; TP.总磷; TN.总氮; NH4+-N.氨氮; NO3--N.硝态氮; NO2--N.亚硝态氮; PO43--P.磷酸盐; SD.透明度; TSS.总悬浮物; X1~X12为优势种代码(表2). DO.dissolved oxygen level; T.water temperature; TP.total phosphorus level; TN.total nitrogen level; NH4+-N.ammonia nitrogen level; NO3--N.nitrate nitrogen level; NO2--N.nitrite nitrogen level; PO43--P.phosphate level; SD.Secchi disk transparency; TSS.total suspended material level; X1—X12.dominant species code(Tab.2).
除浮游植物外,食物链中的次级消费者(主要是鱼类)对浮游动物牧食压力也会影响其群落结构特征。王亚龙等[8]在2014年对长湖鱼类进行资源调查发现,长湖鱼类资源已出现衰退,以湖泊定居型为主,优势鱼类以小型鱼类[鲫鱼(Carassiusauratus)、(Hemiculterleucisculus)、麦穗鱼(Pseudorasboraparva)、棒花鱼(Abbottinarivularis)、小黄黝鱼(Micropercopsswinhonis)、达氏鲌(Culterdabryi)等]或大型鱼类[鲢鱼(Hypophthalmichthysmolitrix)、鳙鱼、鲇鱼(Silurusasotus)、鲤鱼(Cyprinuscarpio)等]的幼体为主,而幼鱼和滤食性鱼类对浮游动物的摄食需求较大,也会影响其群落组成、丰度和生物量等,且具有较明显的季节性变化。
3.2 浮游动物优势种
本次调查优势种共8种轮虫和4种原生动物。其中,萼花臂尾轮虫和针簇多肢轮虫在4个季节的出现频率均为100%;夏季出现频率为100%的种类还有前节晶囊轮虫、湖生累枝虫和尖顶砂壳虫;秋季出现频率为100%还有盖氏晶囊轮虫;冬季还有螺形龟甲轮虫。这些种类均为富营养化水体常见种,一般属广温性种类,分布广,适应性强。从优势度指数来看,针簇多肢轮虫在4个季节的优势度指数均为最高,不同季节优势度指数较高的种类还有:夏季萼花臂尾轮虫(0.184),秋季盖氏晶囊轮虫(0.235)、迈氏三肢轮虫(0.111),冬季螺形龟甲轮虫(0.114)。这些种类均属于典型浮游种类,水平分布和垂直分布均较广。据报道,营浮游生活的典型轮虫的形态往往有明显的季节变异现象[29]。本次调查鉴定中也发现,螺形龟甲轮虫夏季类型的棘突明显较冬季类型长且明显,这也是轮虫适应环境的一种变异。
物种聚类和排序的结果反映了物种分布和数量,非度量多维尺度分析,被认为是研究生态学较好的方法之一。本次对优势种的聚类分析结果表明,卜氏晶囊轮虫和缘板龟甲轮虫聚为一簇且Height值较高,萼花臂尾轮虫和针簇多肢轮虫次之,表明了同簇内物种占据相似的生态位或对生境有相同或相似的需求。本次各季节不同采样点(以对应季节该样点浮游动物为依据)的非度量多维尺度分析二维点阵排序图胁强系数小于0.1,较好地体现了不同季节各采样点物种的关系。不同季节各采样点均能较好地聚为一类,这充分体现了浮游动物分布的季节差异性。夏季主要以耐高温、高营养盐的种类居多,而冬季则以耐低温种类为主,春、秋季则由于温度、光照度等因素的相似性而出现部分重叠,也表现出物种对生境有相同或相似的需求,而相同季节各采样点(丰度和生物量)之间的分化不明显。
3.3 与环境因子的相关性分析
除受到食物供给、捕食、竞争等生物因素影响外,浮游动物的群落结构特征也与水环境因子等非生物因素紧密相关。冗余分析排序可以分析物种与环境因子的关系,反映出物种对环境的需求程度[30]。从本次浮游动物优势种与环境因子的冗余分析可以看出,总氮、总磷、硝态氮、磷酸盐、总悬浮物以及pH等是影响浮游动物群落变化的重要环境因子。如夏季前节晶囊轮虫、湖生累枝虫、尖顶砂壳虫和似钟虫均与总氮含量表现出较明显的正相关性;春季萼花臂尾轮虫与硝态氮含量以及秋季与总磷含量均呈明显的正相关关系。姚佳玫等[31]对富春江的调查发现,在一定范围内淡水水体中轮虫种类组成和数量、密度与pH呈正相关,一般表现为:pH升高轮虫数量和密度也会增加。此外总磷还是评价水体富营养化程度的重要指标——尤其浅水湖泊中,富营养化程度随着总磷水平的升高而加剧,浮游植物的密度和生物量也随之升高,这也影响轮虫群落结构。一些原生动物和轮虫对水体的营养条件有偏好性,如鞭毛虫类和臂尾轮虫明显偏好富营养化水体,有柄的纤毛虫类如小口钟虫(V.microstoma)和褶累枝虫(E.plicatilis)等通常作为污水处理的指标生物。如本次调查中的优势种萼花臂尾轮虫、湖生累枝虫、似钟虫和螺形龟甲轮虫均是富营养化湖泊和池塘的常见种,而尖顶砂壳虫和王氏拟铃壳虫则为寡污带常见浮游原生动物,因此以原生动物和轮虫为环境指示种和水环境状况评价指标。
大量研究结果表明,水温也是对水生生物生长发育及群落特征影响较大的环境因子[32-35]。本研究中,长湖浮游动物丰度、生物量及物种丰富度从春季到夏季皆随温度的升高而上升,从夏季到秋季再到冬季则相反,冗余分析结果与该直观变量的变化结果相一致。笔者还发现,轮虫类群的丰度受水温、溶解氧及pH的影响较大,营养盐也对其有一定的影响。原生动物丰度除了受到水温、溶解氧、pH的限制以外,还受悬浮物、总氮及总磷等指标的影响。长湖的地形地貌及其他生物和非生物因素如水流、水位、降雨量、水草种类与分布等也会一定程度影响浮游动物。
3.4 长湖浮游动物演替特征
从本次调查中季节更替率来看,除枝角类冬—春季节物种更替率较低(25.00%)外,其余各种浮游动物的季节更替率均超过50%,其中超过80%的有秋—冬季原生动物(90.00%)、冬—春季轮虫(81.81%)、秋—冬季桡足类(83.33%)和冬—春季桡足类(87.50%),表明浮游动物各类群季节变化明显,这与浮游植物季节性变化规律密切相关。有研究发现[36],夏季浮游植物以喜高温的蓝藻和绿藻门种类为主,尤其以具有假空泡的蓝藻门占优势,早春、晚秋以及冬季均以耐低温的金藻和硅藻种类为主。因此长湖浮游动物的种群更替率也受同期浮游植物的种类组成、丰度和生物量等因素影响。
本次调查是继郭坤等[14]之后第二次对长湖浮游动物的系统研究,并增加了对原生动物的种类鉴定。这两次调查结果比较发现,长湖浮游动物种类多样性降低,丰度升高(轮虫种类数42种减少为29种,枝角类16种减少为7种,夏季平均丰度4412 个/L升高为6781 个/L等)。这表明长湖水域生态系统的稳定性继续减弱,水体富营养化程度进一步加深。另一方面,优势种仍以浮游轮虫为主,且大部分属富营养化水体的指示种。优势种增加了4种原生动物,其中湖生累枝虫和似钟虫也偏好多污带或中污带水体,这些都表明长湖水域仍处于富营养化状态,这也与本研究中用浮游动物丰度、生物量以及水环境因子对水质的评价结果相一致;此外,该结果还与魏志兵等[37]对同时期长湖浮游植物的研究所得结论相符。本研究除了增加了原生动物种类外,还对优势种和各季节采样点进行了聚类分析,较好地体现了浮游动物优势种以及各采样点在不同季节的关系,较2012年的调查又有了更深入的分析。
