不同光照下梅尼小环藻对昼夜温差变化的生理响应
2021-05-19谭清乾王沛芳马晶洁
谭清乾 ,王沛芳,王 洵,马晶洁,胡 斌
(1.河海大学环境学院,江苏 南京 210098;2.河海大学浅水湖泊综合治理与资源开发教育部重点实验室,江苏 南京 210098)
联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change, IPCC)第五次气候变化评估报告显示,全球变暖已是不争的事实,报告同时还指出北半球大部分地区都出现不同程度的不对称增温,导致昼夜温差不断缩小[1-2]。这对各类生态系统产生了诸多不利影响[3-4],许多陆地、淡水和海洋物种的分布范围、季节性活动及物种间的相互作用已经发生改变。有效减少全球气候变化带来的负面影响,一方面要积极采取应对措施,另一方面也需要了解生态系统响应气候变化的机制。
研究发现气候变暖会引发藻类水华的暴发[5],从而影响水源供应、危害人类健康、降低生物多样性并严重破坏生态平衡[6]。如何预测、控制水华的暴发,对保护我国水资源具有重要意义。目前多数研究都用平均温度的升高来衡量气候变暖对藻类的影响[7-10],昼夜温差的变化容易被研究者忽视。藻类许多生理活动存在明显的昼夜节律[11-12],有研究指出夜温升高能促进藻类生物量与油脂含量的增长[13-14],这表明昼夜温差减小可能会促进藻类繁殖,破坏水生态系统的稳定性。大量研究证实温度与光照对藻类的影响存在耦合机制[15-17],Han等[18]发现低光环境中分离的藻类能抵抗夜温变化所带来的影响。因此在不同光照条件下,藻类对昼夜温差变化的响应可能存在差异。目前关于昼夜温差对藻类生长影响机制的探索不够,一方面研究多集中在藻类生物量、脂肪含量的变化,对光合作用、营养盐利用等生理特征的研究较少;另一方面,在考虑昼夜温差的影响时,忽略了其他环境因子的协同作用,从而降低研究的精准性。

1 材料与方法
1.1 试验材料
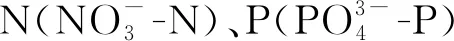
1.2 试验设计
正式试验在步入式恒温恒湿实验室(MHPW-020-TSW)内开展。为模拟不同环境的光照条件,参考我国部分地区太阳辐射监测数值,将正式试验光照强度设置为高光照(1 500 W/m2,记为G)和低光照(1 000 W/m2,记为P)[21-22],光暗周期均为 14 h∶10 h,结合硅藻水华多发地汉江4月间的天气数据[23-24]与中国昼夜温差的平均下降速度,并参考陈星星等[25]对昼夜温差试验的设置方法,在高、低光照强度下将昼夜温度设置3组:25 ℃/15 ℃、25 ℃/20 ℃、25 ℃/25 ℃,分别记为G1、G2、G3、P1、P2、P3,每组试验均设置3组平行。
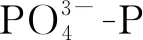
1.3 指标测定
1.3.1藻细胞密度与叶绿素a的测定
利用紫外可见分光光度计(TU1901)测定其在680 nm波长处的光密度(OD680)值,得到藻细胞密度[27-28]。叶绿素a质量浓度由浮游植物荧光仪(Phyto-PAM-Ⅱ)测定[29]。
1.3.2叶绿素荧光参数的测定
各处理组分别取2 mL藻液作为样品,利用Phyto-PAM-Ⅱ测定各叶绿素荧光参数[30-32]。
4.1.1 以护理项目为评价对象 护理项目是质量评价的基本单元,传统的护理质量评价主要将护理项目作为评价对象,如特护、1级护理质量、护理技术操作合格率、健康教育的实施效果等。
1.3.3水样指标测定
1.3.4硅吸收动力学参数测定
培养结束后收集藻液,经无硅培养基清洗2遍后,进行饥饿处理,饥饿过程持续2 d[33],每天监测藻细胞密度、光合活性,当其下降时,于4 000 rpm离心5 min,丢弃上清液,浓缩收集,在无光的环境中培养1 d,之后将藻液接入不同硅浓度(Na2SiO3)(0.5 mg/L、1 mg/L、2 mg/L、4 mg/L、8 mg/L、16 mg/L、20 mg/L、32 mg/L)[33]的8个250 mL锥形瓶中,测定初始藻细胞密度,并放至对应的光照条件下进行培养。接种完成立即取样5 mL经0.22 μm膜过滤后测定初始溶解态硅(DSi)浓度,经过3 h培养后[33]取样测定各培养基中DSi的浓度。DSi浓度的测定采用1,2,4氨基萘酚磺酸还原-硅钼蓝法[34]。最大吸收速率和半饱和浓度根据米氏方程公式拟合得到,吸收速率通过每个培养基中减少的Si质量浓度计算[35]:
(1)
式中:V为吸收速率,ng/(个·h);Vm为最大吸收速率,ng/(个·h);Km为半饱和浓度,表示当V=1/2Vm时底物Si的质量浓度,ng/L;ρ为培养液中Si的质量浓度,ng/L。
1.4 数据处理

2 结果与分析
2.1 生物量变化
2.1.1细胞密度

(a) 高光照条件
2.1.2叶绿素a质量浓度
图2为昼夜温差和光照强度对梅尼小环藻叶绿素a质量浓度的影响。由图2可见,培养结束后,G1、G2、G3组的叶绿素a质量浓度显著高于对应的P1、P2、P3组(P≈0.000<0.05),G处理中增长最多的是G2组,从培养前的155.97 μg/L增加到924.15 μg/L,增长率为492%,P处理中P2组增长最多,从153.7 μg/L增加至705.3 μg/L,增长率为358%,低于G2组,这与藻细胞密度的变化趋势一致。在G处理下,各组叶绿素a质量浓度在第7天出现显著差异(P<0.05),培养结束后表现为G2>G3>G1,组间差异进一步扩大(P<0.01)(图2(a))。图2(b)显示在P处理下,各组叶绿素a质量浓度在第9天出现显著差异(P<0.05),培养结束后表现为P2>P3>P1,与G组相似,组间差异显著(P<0.05)。在G、P处理下,恒温组的叶绿素a质量浓度均高于10 ℃温差组,低于5 ℃温差组,这与藻细胞密度的变化趋势不同。

(a) 高光照条件
2.2 光合活性变化
2.2.1最大量子产生量
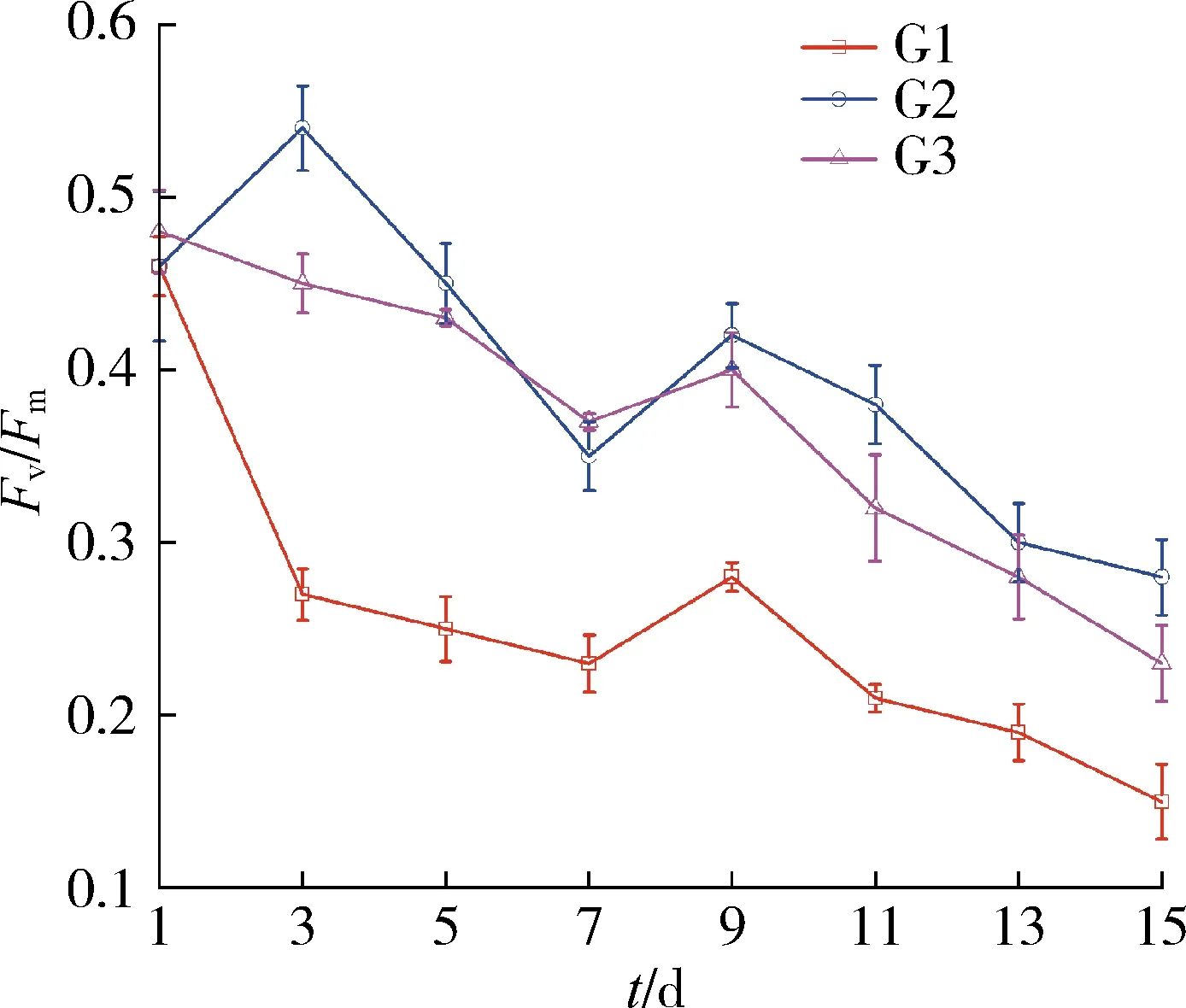
各组梅尼小环藻的最大量子产量(Fv/Fm)变化如图3所示。培养结束后,P处理下的梅尼小环藻Fv/Fm均显著高于G处理(P<0.05),各组Fv/Fm总体上呈现波动下降趋势。从图3(a)可见,在G处理下,不同温差组梅尼小环藻Fv/Fm下降幅度不同,在培养的第3天,G1组Fv/Fm下降幅度远高于其他两组,从0.46下降到0.27,随后趋于稳定;G2组与G3组的变化幅度相似,整体呈现缓慢下降趋势;培养结束后,G1、G2、G3分别下降了65.2%、36.3%、45.9%,组间差异显著(P<0.05)。图3(b)显示,P处理下,各温差组的变化表现出相似的规律,呈现波动下降的趋势,均能维持较高的Fv/Fm;其中P1、P2、P3分别下降了22.4%、19.6%、21.7%,组间差异不明显(P>0.05);从总体上看,P处理下各组间Fv/Fm总体变化趋势相同,各组培养结束后的最小值无显著差别。
2.2.2快速光响应曲线
图4为梅尼小环藻快速光响应曲线的变化。图4(a)(b)显示,P处理下梅尼小环藻的快速光响应曲线均高于G处理,这是不同光照条件所带来的差异。在G处理下,各组梅尼小环藻的光利用率(α)中,G1组显著低于G2、G3组(P<0.05),而G2、G3组变化趋势大致相同。在P处理下,各组变化趋势保持一致,均呈现先波动升高后缓慢下降,组间差异不明显(P>0.05)。图4(c)(d)(e)(f)为不同组中梅尼小环藻最大电子传递速率(Emax)和光饱和点(Ik)的变化曲线。G、P处理下,各温差组Emax和Ik的变化趋势相似,呈现先缓慢下降后保持基本稳定状态,组间差异不显著(P>0.05),在培养的前5天呈现下降趋势,第7天时进入稳定状态。
(a) 高光照条件
2.3 磷利用情况

(a) 高光照条件下α的变化

图5 不同处理下质量浓度随时间变化
2.4 硅吸收动力学特征
利用Michaelis-Menten动力学方程的非线性拟合,得到不同组梅尼小环藻对硅的最大吸收速率Vm和米氏常数Km(表1)。试验结果表明在G、P处理下,各温差组的Km和Vm均表现出显著差异(P<0.05)。不同光照条件下,均是10 ℃温差组的Km值大于5 ℃温差组,表明在一定范围内,梅尼小环藻的Km值随着温差减小而减小。在G处理下,5 ℃温差组的Km值小于恒温组,而在P处理下,5 ℃温差组的Km值大于恒温组,表明了在不同光照下昼夜温差对其Km值的影响不同。不同光照条件下,梅尼小环藻Km的最小值不同。G处理中G2组的Km最小,为 1.755 ng/L,而P处理中P3组Km最小,为4.673 ng/L,是G2组的2.6倍,说明梅尼小环藻Km值随着光照强度的升高而减小,对硅的吸收能力更强。

表1 梅尼小环藻硅吸收动力学参数
3 讨 论
3.1 梅尼小环藻生物量累积对昼夜温差的响应
在淡水生态系统中,昼夜温差会影响藻细胞的合成代谢、营养盐吸收等生理过程[36],从而影响藻类的生长繁殖。有研究表明藻类生物量在夜温越高的条件下损失越大[14],由于暗呼吸速率增加,消耗的物质越大,从而不利于生物量积累。然而本研究结果表明,昼夜温差从10 ℃减小至5 ℃时,促进了梅尼小环藻细胞密度及叶绿素a的增长,而从5 ℃温差变为恒温培养时,其生长受到了一定程度的限制。这是因为在夜温较低的条件下,夜间增温能够加强藻类的各项生命活动,有利于细胞的增殖[37]。而当夜间温度升高至一定界限时,继续增温导致呼吸作用所消耗的物质大大增加,从而消耗了大量的碳水化合物,不利于细胞的生长,这与杨未男[13]发现适宜温差能显著促进小球藻生长的结论一致。目前,气候变化正在朝着昼夜温差不断减小的趋势变化,中国昼夜温差的平均缩小速度为每10年 -0.25 ℃[38],在短期内这种变化趋势对梅尼小环藻的生长繁殖有利,这可能为硅藻水华的暴发提供了有利条件。在高光照下,梅尼小环藻的生长速度均高于低光照培养,并且昼夜温差减小对其生长繁殖的促进作用更加显著,这表明在高光环境中,一定范围内昼夜温差的减小可能更容易导致其大量繁殖。梅尼小环藻是水库等深水水体主要的有害浮游植物[39],是常见的硅藻的水华优势种,对低温具有较强的耐受性,能够在高海拔水库环境中正常生长[40]。在这种环境中,光照强度大,昼夜温差减小对其生长的促进作用更加显著,这可能使梅尼小环藻异常增殖,进而导致水库水质变差、出水水质异味等问题[41],给我国水资源的保护带来巨大的危害。
3.2 梅尼小环藻光合活性对昼夜温差的响应
昼夜温差可以通过影响藻细胞中酶的活性,间接对光能的获取、利用产生影响[15]。本研究发现,昼夜温差对低光照下梅尼小环藻的Fv/Fm无显著影响,各组叶绿素荧光参数都呈现出相同的变化趋势,在整个培养期间都保持较高水平。在高光照条件下,10 ℃温差组的Fv/Fm显著低于其他两组,并且叶绿素荧光参数中α显著降低。费磊[42]研究发现夜间增温会增强第2天藻类的光合作用,这与本研究结果相似,可能是因为夜温升高提高了RubisCO酶活性,进而提高了藻细胞的光利用率[43]。这表明在高光环境中,昼夜温差的减小将促进梅尼小环藻的光合活性,有利于其合成三磷酸腺苷(ATP)用于生长代谢。这也在一定程度上说明了为何在高光环境中,昼夜温差减小对梅尼小环藻生长促进更加明显。因此利用简单的光控制措施如物理遮盖、种植水生植物等,在小型富营养水体中控制硅藻水华可能是有效的方法。
3.3 梅尼小环藻磷利用对昼夜温差的响应
3.4 梅尼小环藻硅吸收动力学特征对昼夜温差的响应
营养盐吸收动力学参数Km是目前最常用表达藻类对营养盐亲和力的参数,Km的降低表明藻类竞争营养盐的能力增强[47]。试验结果显示,高光照下梅尼小环藻对硅的亲和力均高于低光组,吸收效率更高。一些研究同样发现,光胁迫会导致硅藻对硅的吸收增加[48]。这可能是由于藻类对营养盐的吸收需要消耗ATP[33],在高光环境中,梅尼小环藻能合成更多的ATP,因此对硅的吸收能力更强。有研究指出夜晚温度升高,会影响浮游植物对营养盐的吸收[36]。本研究发现,随着昼夜温差从10 ℃减小至5 ℃,梅尼小环藻对硅的亲和力显著提高,这一变化在2个光处理组中均有出现,表明在一定范围内,昼夜温差的减小有利于梅尼小环藻对硅的吸收。硅在硅藻的生长、繁殖过程中起到了重要的作用,硅的吸收与耗竭是硅藻水华发生与消亡的重要原因[49],有研究发现在硅丰富、氮磷营养盐缺乏的水体中,当气候环境适宜时,硅藻仍然能够正常繁殖,甚至形成水华[23]。随着全球昼夜温差减小,梅尼小环藻对硅亲和力的增加,可能满足其快速生长对硅的高需求,从而导致在贫营养水体中大量增殖,危害水生态系统的稳定性。当昼夜温差从5 ℃减小至0 ℃时,高光组中梅尼小环藻对硅的亲和力下降,低光组则相反,可能是在不利光照条件下藻类的一种生理调节[36]。这表明在不同光环境中,梅尼小环藻硅吸收对昼夜温差的响应存在一定的差异。
4 结 论
a. 在5~10 ℃范围内,昼夜温差的减小有利于梅尼小环藻生物量的积累,高光照下的促进作用更加明显。在全球昼夜温差缓慢减小的背景下,这种变化将有利于梅尼小环藻的生长繁殖,为水华的发生提供有利的温度条件。
b. 不同光照下梅尼小环藻光合活性对昼夜温差的响应不同。昼夜温差对低光照下梅尼小环藻的光合活性无显著影响;在高光照下,5 ℃~10 ℃范围内的温差减小,会增加其光利用率,提高光合活性。在水华的防止过程中,可采用简单的光控制措施限制硅藻的生长。
