寒地早粳稻牡育稻42系谱构建及遗传基础解析
2021-05-19李洪亮孙玉友魏才强程杜娟曲金玲刘春光徐德海时新瑞
李洪亮, 孙玉友, 魏才强, 解 忠, 程杜娟, 姜 龙, 曲金玲,宋 泽, 刘春光, 徐德海, 王 丽, 时新瑞
(黑龙江省农业科学院牡丹江分院, 黑龙江 牡丹江 157041)

表1 牡育稻42的主要特征特性
黑龙江省水稻生产是我国粮仓储备的重要保障之一,在保证产量的同时提高稻米品质是当前育种工作的主要目标。黑龙江省具有独特的地理生态环境,以盛产优质粳稻著称,加强优质、抗逆品种选育,提高优质稻米市场竞争力,对保障我国粮食安全意义重大[1]。黑龙江省农业科学院牡丹江分院水稻所早在1958年就从事水稻育种研究,截至目前,育成了一批“牡丹江”系列优质、高产水稻品种,为黑龙江省东南部稻区水稻产业发展发挥了重要作用。牡育稻42 是黑龙江省农业科学院牡丹江分院以牡00-1066为母本,垦稻12为父本杂交选育而成的早粳高产型品种,于2019年5月通过黑龙江省农作物品种审定委员会审定(黑审稻20190015),适宜在黑龙江省≥10 ℃活动积温2 450 ℃区域种植,该品种的突出特点是早熟、矮秆、丰产、抗倒伏性极强。牡育稻 42 的育成创制了矮秆抗倒并兼具丰产特性的寒地粳稻新种质,是进一步开展高产、抗倒伏育种可供选择应用的优良亲本材料。阐明优良品种的遗传性状传递过程,对未来明确寒地水稻育种方向、选择优良亲本等具有重要意义。
1 材料与方法
1.1 材 料
牡育稻42主茎12片叶,长粒,抗病及耐冷性强,平均产量8 339.0~8 859.3 kg·hm-2,适宜区域为黑龙江省第二积温带下限,需≥10 ℃活动积温2 450 ℃左右[2]。本研究通过查阅《中国水稻品种及其系谱》[3]、《寒地粳稻育种》[4]构建了牡育稻42遗传系谱图。
1.2 方 法
针对牡育稻42亲本进行追溯并构建系谱图,并对牡育稻42的细胞核和细胞质遗传物质组成进行解析。细胞核遗传物质通常来自父母本双亲,则规定父本、母本细胞核遗传物质的贡献率各自计为50%,而通过系统选育、诱变处理和单倍体等技术手段育成的品种,其亲本细胞核遗传物质的贡献率计为100%;细胞质的遗传物质是通过母本独立遗传的,因此,规定细胞质遗传物质的贡献率全部来自母本计为100%。依照以上计算规则和方法,对牡育稻42细胞核和细胞质遗传物质组成进行逐级追溯和深入解析。在构建系谱图时,上方位置为母本材料、下方位置为父本材料。
2 结果与分析
2.1 牡育稻42系谱图
如图1所示,牡育稻42细胞质遗传物质由龙粳长粒香通过牡00-1066传递至牡育稻42,其贡献率为100%。
核遗传物质则由祖先亲本(原始亲本)龙粳长粒香、富士光、台中27、高雄53、北海84、石狩白毛、色江克、合选58、农林11、下北、虾夷、笹锦、奥羽239、曲系17和藤系117按不同比例共同提供。
牡育稻42的育成是以合江20、东农3134等关键桥梁种质的成功创制为基础,同时也离不开虾夷、富士光和藤系138等重要亲本材料的成功引入[5]。
建国前,黑龙江省水稻生产上的主要品种来源是通过引种和试种两种方式,其中以从日本引种为主,石狩白毛等在此阶段引入我国。建国后,育种工作分为评选地方良种、系统选种、杂交育种和综合技术育种4个阶段。前3个阶段引入了下北和虾夷等骨干亲本材料。20世纪70年代,杂交育种、单倍体育种、诱变育种等多种育种方法的出现标志着我国育种工作进入第4阶段,此阶段育成了代表性品种普选10号和合江20。20世纪80年代,合江22、牡丹江19、东农413和东农415等一批综合性状好的品种被相继育成,同时从日本引入了优良品种富士光和藤系138,使得此时期黑龙江省水稻生产进入了一个新的发展时期。20世纪90年代,陆续育成了龙粳10号、东农416、牡丹江22、绥粳4号、五优稻1号、垦稻8号等一大批优异品种。21世纪,优质高产品种东农428、五优稻4号、垦稻12、龙稻5号和龙粳59[6]等被成功选育。正是经过前几个不同育种阶段各个祖先亲本遗传基础的积累,才成功选育出寒地高产、抗逆早粳稻新品种牡育稻42。
2.2 亲本来源与遗传解析
牡育稻42母本分支较为简单,母本牡00-1066是由龙粳长粒香和富士光杂交育成,富士光遗传有日本知名优质米品种越光的基因,牡00-1066不但遗传了亲本品质优良的特性,同时具有株高相对较矮、抗倒伏性极强、丰产性突出的特点。
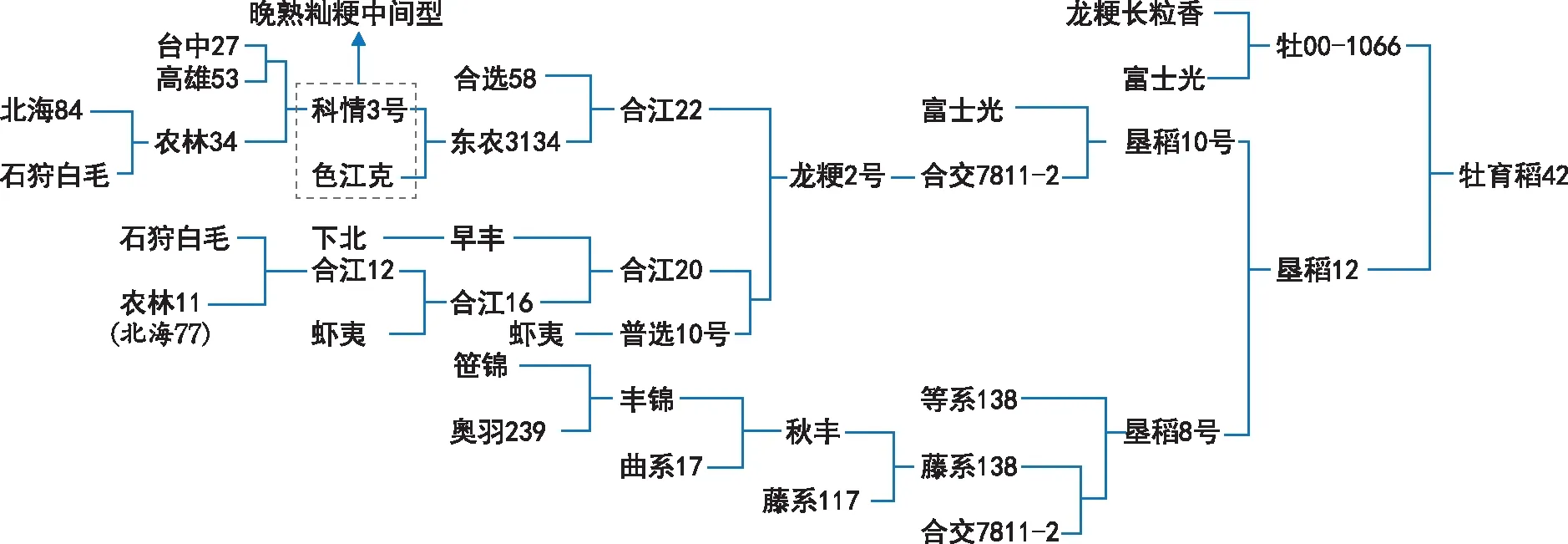
图1 寒地早粳稻牡育稻42系谱图

表2 牡育稻42亲本
从牡育稻42父本垦稻12分支上进行分析(图1),牡育稻42的育成离不开合江号种质材料的成功创制,黑龙江省农业科学院水稻研究所通过将石狩白毛[7]与农林11进行配组杂交,利用双亲的耐寒和抗倒伏特性,使后代材料合江12具备了耐寒和抗倒的特点;又以具有多蘖特性的虾夷材料[8]为亲本与合江12进行有性杂交,选育出了具有中等分蘖力的合江16;进而以合江16和早丰(下北)作为双亲材料,成功创制出超亲材料合江20,合江20是当时育成的集分蘖力、抗病性和抗倒性均较强的代表性品种之一[8-10];东北农业大学引入了科情3号(遗传了台湾品种台中27和高雄53的籼稻血缘)与巴基斯坦品种色江克进行配组杂交,育成了遗传基础相对较宽、抗病性极强的中间材料东农3134,东农3134在1969—1971年间日本品种及当地品种严重感染稻瘟病的状况下表现出极强的抗病性,且稻瘟病抗性多年间表现稳定,但因其耐寒性差、对光温反应十分敏感等缺点而难以大面积推广。黑龙江省农业科学院水稻研究所利用合选58与东农3134进行杂交,将合选58的耐冷基因与东农3134进行整合,育成了综合抗性强、耐冷、耐深水品种合江22。继而利用合江20、合江22和普选10号(没有遗传到虾夷多蘖的特性)创制出重要中间材料合交7811-2,合交7811-2与适应性好、配合力高、遗传基础广的藤系138[11-12]和富士光[13]进行基因重组,分别育成了活秆成熟、外观米质优的垦稻8号[14]和垦稻10号[15],为进一步育成早熟、抗病、优质、丰产、适应性强的垦稻12[16](缺点抗倒伏性较差)奠定了直接材料基础。牡育稻42是通过垦稻12与抗倒性极强,丰产性突出的牡00-1066进行遗传性状重组而选育出的产量性状突出、综合特性优异、区域适应性广的早粳高产抗逆型新品种。
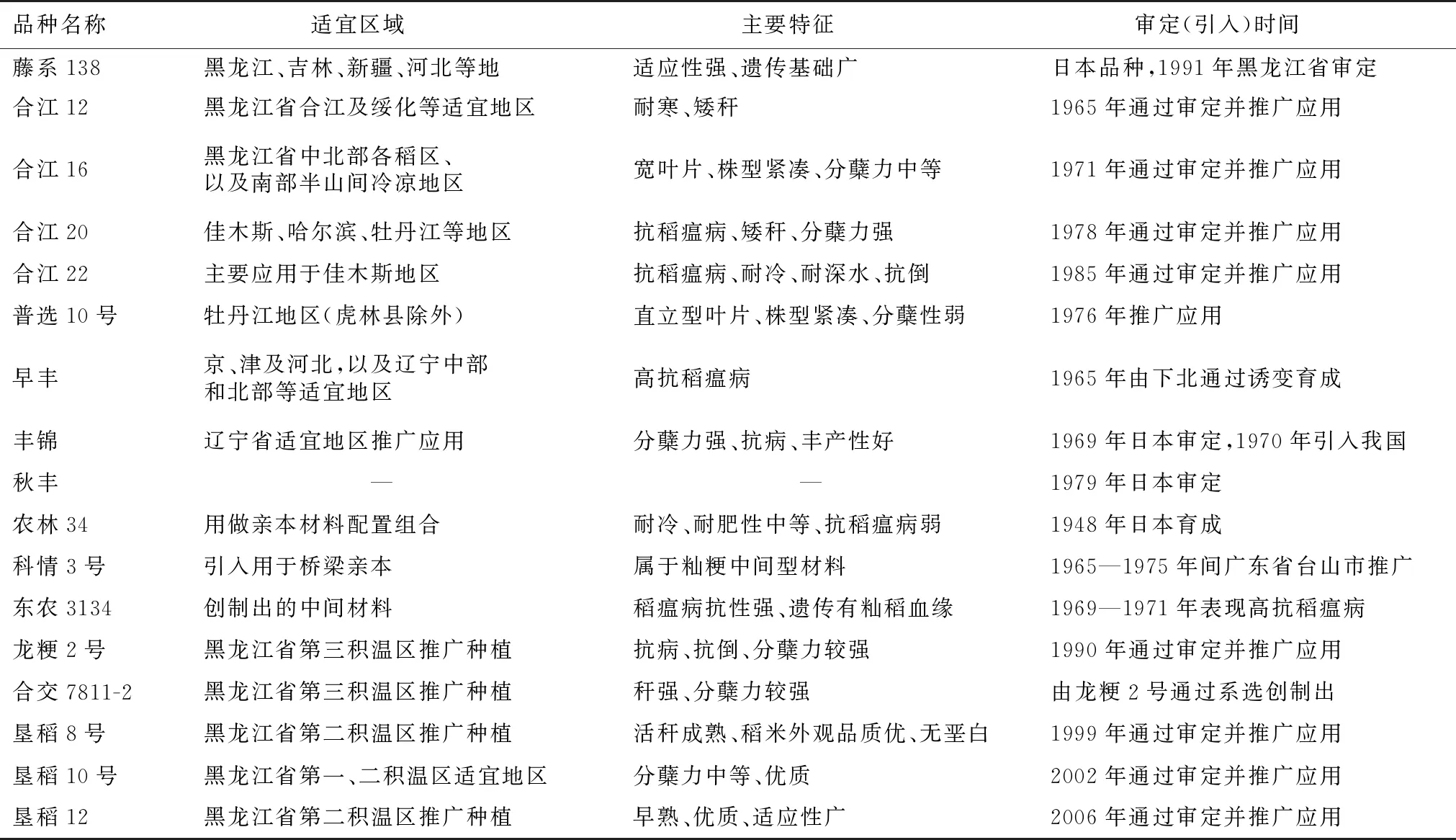
表2(续)(Continued Table 2)
2.3 亲本核遗传贡献率
由表3可知,以原始骨干亲本进行核遗传贡献率分析。龙粳长粒香只被应用1次,核遗传贡献率为25%;而富士光、台中27、高雄53、北海84、色江克、合选58、农林11、下北、笹锦、奥羽239、曲系17和藤系117分别被应用2次,核遗传贡献率分别为37.5%、0.585 937 5%、0.585 937 5%、0.585 937 5%、2.343 75%、4.687 5%、0.585 937 5%、2.343 75%、2.343 75%、2.343 75%、4.687 5%、9.375%;石狩白毛和虾夷分别被应用4次,核遗传贡献率分别为1.171 875%和5.859 375%。通过计算表明,台中27、高雄53、北海84和农林11的核遗传贡献率相对较小。

表3 牡育稻42祖先亲本核遗传贡献率
3 讨 论
3.1 准确制定育种目标与精心选择亲本材料
追溯牡育稻42祖先亲本系谱过程中,发现其聚合了大量不同生态区的基因性状,其中包括石狩白毛的耐寒性状、富士光的优质性状、虾夷的多蘖和抗病性状。同时,籼粳亚种杂种优势得以充分利用,这主要体现在对含有籼稻基因的台湾品种台中27和高雄53以及巴基斯坦品种色江克的应用上。正是通过在自然及人工高压选择下,不断累加亲本的优良性状、剔除无益性状,使牡育稻42最终具有集高产、优质、抗病、抗倒伏、适应性广为一身的特征特性。
将已育成品种的系谱进行深入分析,对进一步开展育种工作具有重要意义,它能阐明品种各性状的遗传传递过程,进而总结出优良亲本选配方法。首先,在搜集的众多品种资源中选择具有育种目标性状的材料,同时要考虑所选材料是否兼具较多的其他优良性状。目标性状要具体,更要明确目标性状的构成性状。如在抗病育种中要明确其所抗病害的种类、抗谱如何等,继而制定出亲本配置组合方案。所配组的两个亲本间可拥有更多的共同优点,但不能有共同的缺点,即双亲要做到性状互补。用经济性状优良、遗传差异大的亲本配组,选出理想类型的几率相对越大。
3.2 打破遗传基础狭窄限制及拓展育种思路和育种方法
寒地水稻育种从最初的引种和试种发展至今,其育种技术水平大幅提升,但就当前寒地水稻育种现状来看,现有的育种材料遗传基础狭窄,新的可利用基因较难被发掘和利用。因此,突破目前现有遗传基础狭窄的限制,创制出突破性的中间桥梁材料十分必要。这就要求资源引入地域要更加远缘,不能再仅以亚洲区域引种为主,应在世界范围内的水稻遗传多样性中心进行种质引进和改良创制。
在育种思路和育种方法上也应有所创新,过去通常建议育种者选择当地主栽品种作为亲本之一,利用其他外引材料对其进行性状改良,好处是后代材料可能在当地会表现出更好的适应性,在时间上也相对容易快速的选育出新品种。但从育种工作的长远角度来看,创制出一批真正突破性的中间桥梁种质是打破当前寒地水稻遗传基础狭窄的必要选择,即使短期育种成果不显著,但对后续寒地水稻育种的长远发展具有重大意义。同时,随着时代发展,当前水稻育种工作已进入了合作交流的时代,既包括育种材料的合作,也包括育种手段的合作,而当前可应用到育种实践中的技术手段还比较单一,新的生物技术与常规育种手段结合还有待深入。
