结球甘蓝α-法呢烯合成酶基因的鉴定与功能分析
2021-05-18张彤汪一萍葛洋NTIRIEric周文武
张彤,汪一萍,葛洋,NTIRI Eric,周文武,*
(1.福建农林大学应用生态研究所,闽台作物有害生物生态防控国家重点实验室,福州350002;2.浙江大学昆虫科学研究所,农业农村部作物病虫分子生物学重点实验室,杭州310058)
植物依赖次生代谢物来应对环境胁迫,并进化出复杂的调控系统来调节这些化合物的合成。植物的次生代谢系统能产生众多的挥发性化合物,其中的植物挥发性有机物(volatile organic compounds,VOCs)在植物对植食者的防御中扮演着重要的角色,也是昆虫识别植物的化学指纹[1]。据现有的研究统计,植物体内的挥发物达3万多种,且它们的分子质量通常较小,在100~200 Da之间[2-3],这些挥发物根据其性质分为普遍存在的挥发物和特异性的挥发物[4]。特异性的挥发物是植物受植食性昆虫取食或侵害后,其体内被激发产生的化学物质[5-6],通常称之为虫害诱导的植物挥发物(herbivoreinduced plant volatiles,HIPVs)。这类挥发物作为信号因子,在“植物-植物”以及“农作物-害虫-天敌”之间的化学通信中行使重要的功能[7-8]。比如,棉花产生的挥发物硝基苯酚[(E)-4,8-dimethyl-1,3,7-nonatriene, DMNT]能抑制海灰翅夜蛾(Spodoptera littoralis)对棉花的取食,并影响成虫的交配行为,从而导致下一代种群个体数显著降低[9]。薰衣草产生的挥发物氧化芳樟醇会对草地贪夜蛾(Spodoptera frugiperda)雄蛾产生显著的驱避效果[10]。大花六道木(Abelia grandiflora)释放的β-蒎烯和苯乙醛可引诱 稻 褐 飞 虱(Nilaparvate lugens)、白 背 飞 虱(Sogatella furcifera)、灰飞虱(Laodelphax striatellus)等水稻害虫的寄生蜂——稻虱缨小蜂(Anagrus nilaparvatae)[11]。玉米植株受挥发物(Z)-3-乙酸己烯酯[(Z)-3-hexenyl acetate, HAC]影响可激活玉米植株内的虫害防御基因的表达,而HAC和吲哚2种挥发物的混用能增强茉莉酸信号的转导、防御基因的表达和次生防御代谢产物的产生,从而增强玉米的抗虫性[12]。
植物萜类合成酶(terpene synthase,TPS)基因是植物合成其萜类化合物的关键酶。根据TPS 的功能差异可以将其分为倍半萜合成酶、单萜合成酶和二萜合成酶等。萜类合成途径可分为甲羟戊酸(mevalonic acid,MVA)途径和丙酮酸(pyruvic acid,MEP)途径,该2种途径都可以合成萜类的前体物质异戊烯焦基磷酸(isopentene coke phosphoric acid,IPP)和二甲基丙烯基焦磷酸(dimethylpropylpyrophosphate,DMAPP)。在法呢基焦磷酸酶或异戊烯基转移酶的催化下,IPP和DMAPP分别生成法呢烯基焦磷酸(farnesenyl pyrophosphate,FPP)或牻牛儿基焦磷酸(geranyl pyrophosphate,GPP)和牻牛儿基牻牛儿基焦磷酸(geranylgeranyl pyrophosphate, GGPP)。FPP、GPP和GGPP在萜类合成酶的作用下生成异戊二烯单体分子以及由这个单体经复合反应产生的有机物,比如半萜(C5)、单萜(C10)、倍半萜(C15)以及二萜(C20)等。
随着植物基因组测序技术的发展,植物TPS 家族规模不断扩大。根据现有的研究统计,植物中各类萜烯合成酶基因总数从30~100 个不等[13],特别是葡萄、苹果、毛果杨和大豆等植物的基因组测序促成了对它们萜类合成酶家族基因的解析和功能分析[14]。基于萜类合成酶的重要功能,一些研究者在农作物中围绕TPS基因开展了深入研究。这些工作从沉默萜类合成的关键基因出发,通过抑制特定萜类在植株内的合成和释放,解析萜类挥发物功能。如通过沉默水稻石竹烯合成酶基因OsTPS3的表达,水稻释放的石竹烯吸引稻飞虱及其天敌稻虱缨小蜂的机制得到了揭示[15-16]。此外,在黄豆中过表达GmAFS基因后,转基因黄豆的须根中释放的大量α-法呢烯增强了黄豆对根结线虫的抗性[17]。目前,在拟南芥、烟草和番茄等植物中围绕萜类合成酶也已有众多研究[13],但是,有关结球甘蓝(Brassica oleracea)的萜类合成酶尚未有深度的研究和报道。结球甘蓝的全基因组测序为其萜类合成酶基因的鉴定和功能研究提供了可能[18],也为深入解析一些重要萜类挥发物在其体内的合成机制及功能提供了条件。
结球甘蓝是全球范围内广泛栽培的一种十字花科芸薹属蔬菜,具有重要的经济价值。自20世纪90 年代以来,全球芸薹属作物种植面积急速扩大,并以集约化耕作为主,而该属蔬菜在生产中面临病虫害爆发的威胁[17]。小菜蛾(Plutella xylostella)是取食十字花科植物的重要寡食性昆虫[19],前期研究发现,α-法呢烯在甘蓝受小菜蛾幼虫取食后释放,对小菜蛾成虫具有一定的驱避作用;触角电位(electroantennogram,EAG)研究也证实了α-法呢烯能被小菜蛾成虫识别[20],说明该化合物在小菜蛾和结球甘蓝的互作中可能扮演着重要的角色。为解析α-法呢烯在结球甘蓝体内合成的机制,本研究克隆了其合成途径的关键酶α-法呢烯合成酶基因(BoTPSa),阐明了虫害和非生物胁迫因子对其表达的影响,并对α-法呢烯合成酶进行了原核表达和生化功能测定,证实其在萜类合成途径中的作用,并研究了该蛋白在植物体内的亚细胞定位情况。本研究首次系统解析了结球甘蓝的α-法呢烯合成酶的功能,为后续探索该蛋白及其产物的生态功能提供了基础,也为其他十字花科作物的萜类的合成和功能研究提供了依据。
1 材料与方法
1.1 试验材料
选用河北邢壮牌结球甘蓝种子‘京丰一号’(河北省邢台华丰种子有限公司),在光照培养箱中培养,处理周期为14 h光照/10 h黑暗,温度23 ℃,待其生长30 d后,选取长势相近的健康植株进行试验。
小菜蛾在福建农林大学应用生态研究所繁育,幼虫采用白菜幼苗室内喂养,小菜蛾成虫以5%蜂蜜水饲养。小菜蛾世代培育温度设置为24~26 ℃,相对湿度设置为45%~70%,光照周期为17 h光照/7 h黑暗。
1.2 BoTPSa 基因的克隆和重组质粒构建
利用“α-farnesene synthase”“Brassica oleracea”等关键词对结球甘蓝转录组测序结果进行搜索,对搜索到的序列在NCBI数据库(http://www.ncbi.nlm.nih.gov/genome/browse/)中的BLASTX 里再次确认序列的准确性。设计2条可以扩增完整开放阅读框的特异性引物来扩增目的基因全长序列,引物信息如下:上游引物序列为5´-GTTCATCACAATTTTGA GCTCTTT-3´;下游引物序列为5´-TGAGGAGGGA AATAAACCAATA-3´。引物由杭州尚亚生物有限公司合成。以结球甘蓝为材料,RNA提取步骤参照TRIzol Reagent RNA 提取试剂盒说明书(美国Invitrogen 公司)进行,然后使用cDNA 反转录试剂盒(北京全式金生物技术有限公司)合成cDNA第1链。使用Phusion 高保真DNA 聚合酶试剂盒(美国赛默飞世尔科技有限公司)扩增获得目的基因,将该基因命名为BoTPSa。
聚合酶链式反应(polymerase chain reaction,PCR)产物用DNA 凝胶回收试剂盒(美国Vitagene公司)纯化回收,利用重组Taq酶(日本TaKaRa公司)在目的片段中加上黏性末端,产物连入pMD18-T载体后转化入DH5α感受态细胞中,经蓝白斑筛选阳性克隆,由上海泽衡生物有限公司测序后保菌于-80 ℃冰箱中,备用。
1.3 萜类合成酶的生物信息学分析
采用SMART 蛋白结构预测软件(http://smart.embl-heidelberg.de/)对BoTPSa结构域进行预测,接着利用SWISS-MODEL(https://swissmodel.expasy.org/)对BoTPSa 的三级结构进行预测及分析。使用在线软件Clustal Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/)对BoTPSa核酸序列及其他植物的TPS基因进行多序列联配,之后使用最大似然法在MEGA 7.0中输入联配的核酸序列,构建系统进化树。
1.4 结球甘蓝BoTPSa 基因的表达规律研究
分别设置虫害处理组及对照组,逆境处理组及对照组,各组设置5个生物学重复。虫害处理组:提前饥饿处理小菜蛾幼虫,再让其取食甘蓝叶片1 h后移走。之后在停止虫害4、8和20 h后分别对受虫害的结球甘蓝叶、茎和根进行采样,测定各部位的BoTPSa基因表达水平。逆境处理组:1)高盐处理组,用20 mL 200 µmol/L NaCl 溶液浇灌甘蓝,每12 h 一次,浇灌2 次;2)光合作用抑制处理组,将甲基紫精溶液涂抹于叶片近叶柄处,持续24 h;3)茉莉酸甲酯诱导处理组,取1 mL融于羊毛脂(7.5µg/μL)的茉莉酸甲酯涂抹于近叶柄处的后半片叶上,诱导24 h;4)高温处理组,将植株放在38 ℃环境下处理24 h;5)低温处理组,将植株放在4 ℃环境下处理24 h;6)蚜虫胁迫处理组,将5 头蚜虫置于甘蓝叶上,持续24 h后采样。以上样品经液氮研磨后提取RNA,将RNA反转录为cDNA后,于-20 ℃条件下保存。GAPDH和EF-1为内参基因。利用生物软件Geneious(美国Invitrogen 公司)设计2 条定量聚合酶链式反应(quantitative polymerase chain reaction,qPCR)引物,上游引物序列为5´-TGCTCG GGAACAAATATGCG-3´,下游引物序列为5´-TGT TGCCTTAGGAGTCGGAA-3´。引物由杭州尚亚生物有限公司合成。
荧光定量PCR 使用SYBR Premix ExTaqⅡ反应体系在CFX 96荧光定量PCR仪上进行。反应程序如下:95 ℃预变性30 s;95 ℃变性5 s,60 ℃延伸30 s,40 个循环。通过熔解曲线来判断引物的特异性,基因的表达差异用2-△△CT方法进行计算。
1.5 结球甘蓝BoTPSa 原核表达载体的构建
BoTPSa的密码子适应指数(codon adaptation index, CAI)<0.8,评估表明CAI 需要进一步优化,所以将BoTPSa基因序列与原核表达效率更高的pET-32a 载体相连,并使用密码子优化设计网站(www.genscript.com/tools/gensmart-codon-optimization)对密码子进行优化。输入未优化的BoTPSa序列信息,并选择宿主偏好为“Escherichia coli”。待优化结果反馈后(优化后CAI=0.97),使用Geneious软件(美国Invitrogen 公司)分析pET-32a 载体序列和BoTPSa基因序列特点,设计插入位点的碱基排布:1)选择在BoTPSa序列前加入肠激酶(enterokinase,EK)酶切位点碱基序列GACGACGACGACAAG,并在EK酶切位点前加入KpnⅠ酶切位点GGTACC;2)在BoTPSa序列后加入终止密码子TAA 以及XhoⅠ酶切位点CTCGAG。选择自带硫氧还蛋白A标签蛋白的pET-32a 作为结球甘蓝的BoTPSa 原核表达载体。以KpnⅠ和XhoⅠ为双酶切位点,利用同源重组原理,连接pET-32a和优化后的BoTPSa基因,具体试验步骤参照一步克隆试剂盒说明书(日本TaKaRa 公司)进行。将重组pET-32a/BoTPSa转入Trans5α感受态细胞后涂板,于37 ℃条件下过夜培养后挑取单菌落进行PCR 鉴定。菌液PCR 产物经测序验证正确后,重新摇菌提取重组质粒进行双酶切并验证条带大小。提取构建成功的基因表达质粒,再次转化入Rossetta(DE3)表达感受态细胞中,使用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)检测表达情况。
1.6 结球甘蓝BoTPSa 重组蛋白的诱导表达条件优化及其复性
BoTPSa 重组蛋白的诱导表达条件如下:采用0.7 mmol/L 异丙基硫代半乳糖苷(isopropyl-β-Dthiogalactoside, IPTG)进行诱导,分别放入16 ℃和37 ℃的摇床振荡培养箱中,以220 r/min 振荡培养16 h。确定蛋白表达的最适温度后,其他条件不变,分别加入不同浓度(0.1、0.4、0.7、1.0 mmol/L)的IPTG 进行诱导,然后使用SDS-PAGE 检测表达情况。
收集经IPTG诱导后的大量菌液,按照质量体积比1∶10,采用匀浆机充分重悬,之后采用高压匀浆机进行破菌,压力700~750 Pa,温度2~8 ℃,重复2次。破菌液于2~8 ℃条件下,以1.2×104r/min 离心30 min,收集沉淀[破菌液原料及终浓度:50 mmol/L Tris、10 mmol/L 乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)]。包涵体沉淀和洗涤液①按照质量体积比1∶20 在冰上搅拌洗涤2 h(洗涤液①原料及终浓度:20 mmol/L Tris、10 mmol/L EDTA、0.1 mol/L NaCl、1%TritonX-100)。包涵体沉淀和洗涤液①在2~8 ℃条件下,以1.2×104r/min 离心10 min,收集沉淀。包涵体沉淀和洗涤液②按照质量体积比1∶20 在冰上搅拌洗涤2 h(洗涤液②原料及终浓度:20 mmol/L Tris、10 mmol/L EDTA、0.1 mol/L NaCl、2 mol/L 尿素)。包涵体沉淀和洗涤液②在2~8 ℃条件下,以1.2×104r/min 离心10 min,收集沉淀。洗涤后沉淀和裂解液按照质量体积比1∶20 重悬(裂解液原料及终浓度:50 mmol/L Tris-HCl、7 mmol/L GuHCl、1 mmol/L EDTA、0.5%β-巯基乙醇)。低温下利用磁力搅拌至全部溶解后,于2~8 ℃条件下,以1.35×104r/min 离心30 min,收集上清液。缓慢搅动,将变性蛋白液加入复性液中(复性液原料及终浓度:50 mmol/L Tris、0.1 mmol/L NaCl、1 mmol/L EDTA、6 mmol/L半胱氨酸、0.5 mol/L精氨酸、0.6 mmol/L 胱氨酸),于4 ℃冰箱中复性72 h。利用超滤膜对复性蛋白液进行超滤浓缩,并以0.22 μm 孔径滤膜除菌,过滤除菌后的复性蛋白液置于4 ℃冰箱中保存,备用。
1.7 酶活反应产物的气相色谱-质谱分析
超声破碎得到待检测蛋白溶液后,向其加入50 μL 检测缓冲液和一定量的MgCl2、Na2WO4、NaF、FPP,配置成终溶液为100 μL的待测液,且使得混合液 中 含 有10 μmol/L FPP(或GPP 或GGPP 或ddH2O)、10 mmol/L MgCl2、0.2 mmol/L Na2WO4、0.1 mmol/L NaF、0.05 mmol/L MnCl2、50 mmol/L KCl。将装有反应液的玻璃瓶放入预热至30 ℃的水浴锅中,采用顶空固相微萃取法(head space-solid phase microextraction, HS-SPME)(萃取纤维禁止接触样品液面)进行吸附。1 h后,打开气相色谱-质谱(gas chromatography-mass spectrometer, GC-MS)联用仪对各个样品与底物反应后产生的萜类化合物进行检测和分析。GC-MS 色谱条件:采用岛津RTX-1MS非极性石英毛细管柱(30 m×0.25 nm×0.25 μm)。预先设置进样口温度为250 ℃,确定柱箱温度已采用自动升温模式程序:反应开始时,柱箱温度设置为60 ℃,维持3 min;之后,以8 ℃/min的速度升温,将温度升至125 ℃;最后以5 ℃/min的速度升温,将温度升至240 ℃,此过程保留5 min。用氦气当载气,气压为80 kPa,流速为1 mL/min。进样方式:进样不分流,进样方式一栏选择手动顶空进样。质谱条件:电子轰击(EI)式离子源,冲击电压70 V,离子源温度200 ℃,接触表面温度250 ℃,扫描范围30~500 amu。待检测结束后,定性分析每个质谱峰在化合物库中检索出的相应化合物。每个样品重复检测3次。
1.8 原生质体转化及亚细胞定位观察
以pMD18-T/BoTPSa质粒为模板,设计亚细胞定位引物对其进行扩增。亚细胞定位引物上游序列为5´-GCTCTAGAATGAAAACACAGTCACAG CC-3´;下游序列为5´-GGGGTACCTGGAACAGG GTAGACGAGCA-3´。PCR 产物经1%琼脂糖凝胶电泳检测正确后,酶切回收基因片段。使用T4连接酶将目的基因片段与亚细胞定位载体Psup1300 相连,反应体系为:Psup1300载体1µL,目的基因片段7µL,T4连接酶1µL,T4连接酶缓冲液2µL,ddH2O 9µL。再将转入目的基因的亚细胞定位载体转入大肠埃希菌感受态细胞DH5α中。挑取测序无误的阳性克隆进行菌液扩繁并提取目的质粒。获取拟南芥原生质体悬浮液后,共转化目的质粒与标志物基因,在弱光下培养8~10 h后,在激光共聚焦显微镜下观察BoTPSa基因的亚细胞定位情况。
2 结果与分析
2.1 BoTPSa 基因的克隆及其蛋白质三级结构预测分析
经克隆得到的基因序列全长约1 800 bp,与预期相符。将产物连入pMD18-T 载体上,经蓝白斑筛选阳性克隆。对部分阳性克隆菌液进行测序,其结果与预期相符,于是将该基因命名为BoTPSa。利用SMART 和ExPASy-PROSITE 软 件 对BoTPSa 蛋白的结构域进行分析,结果显示,BoTPSa 蛋白含2个保守结构域:萜类合成酶N 末端结构域和C 端活性结构域(图1A)。所以,初步确认BoTPSa 为萜类合成酶家族成员。BoTPSa蛋白的三级结构如图1B所示,以甜橙(Citrus sinensis)的柠檬烯合成酶蛋白为模板,它们的序列同源性为43.98%。
2.2 BoTPSa 基因的进化分析

图1 BoTPSa蛋白的结构域和三级结构Fig.1 Structural domains and tertiary structure of BoTPSa protein
获取拟南芥、玉米以及常见的芸薹属作物等16种植物的TPS核酸序列并构建进化树。结果(图2)表明:BoTPSa基因与芸薹属作物的同源基因(如白菜萜类合成酶基因BrTPS、芥菜萜类合成酶基因BnTPS、油菜萜类合成酶基因BjuTPS和黑芥萜类合成酶基因BniTPS)成簇分布,亲缘关系较近,同时也说明芸薹属作物α-法呢烯合成酶基因高度保守;与拟南芥AtTPS03基因亲缘关系次之;与百合萜类合成酶基因LhTPS、黄瓜萜类合成酶基因CmTPS13、莴苣香叶烯合成酶基因LsTPS、苏拉威西蝴蝶兰萜类合成酶基因PeTPS和文心兰萜类合成酶基因OhTPS的亲缘关系较远。BoTPSa基因与杂交杓兰萜类合成酶基因CpTPS和木本棉萜类合成酶基因GaTPS6、玉米萜类合成酶基因ZmTPS1、蓖麻萜类合成酶基因RcSeTPS13和江南卷柏萜类合成酶基因SmTPS4位于系统进化树中的不同分支上,亲缘关系最远。
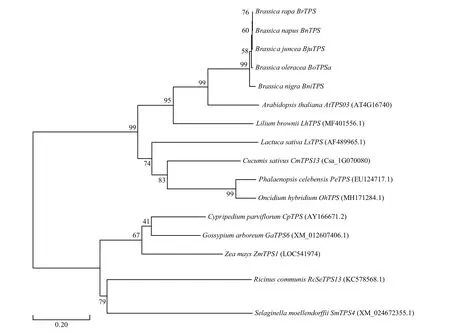
图2 16种植物中TPS基因的系统进化树Fig.2 Phylogenetic tree of TPS genes in 16 plants
2.3 结球甘蓝BoTPSa 基因的时空表达规律
qPCR 结果显示:在停止虫害处理4 h 后,结球甘蓝的各器官(叶、茎、根)中的BoTPSa基因表达量显著上调(P<0.05);虫害处理组中叶、茎、根的BoTPSa基因表达量分别是对照组的11.0 倍、8.1 倍和7.0 倍。在停止虫害处理8 和20 h 后,甘蓝BoTPSa基因表达量仍然显著上调,但诱导表达水平随着时间延长呈下降趋势(图3)。蚜虫的取食危害也能显著诱导BoTPSa基因表达量的上调;试验发现,结球甘蓝叶中的BoTPSa基因表达水平可受低温(4 ℃)和高温(38 ℃)影响而明显上调(P<0.05)(图4),但上调幅度不如小菜蛾虫害诱导。
2.4 结球甘蓝BoTPSa 蛋白的表达
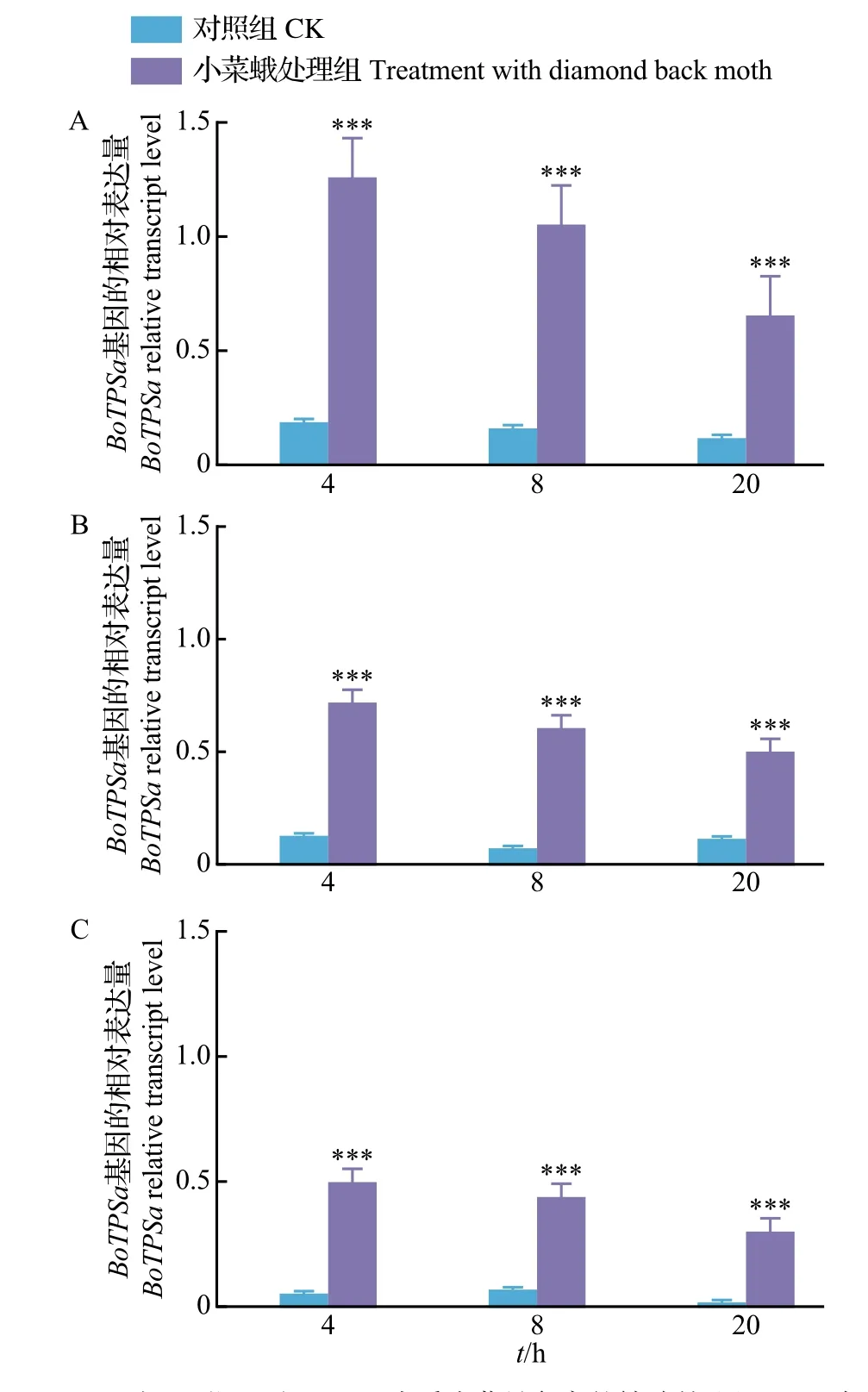
图3 小菜蛾诱导结球甘蓝叶、茎、根BoTPSa 基因的相对表达量Fig.3 Relative transcript levels of BoTPSa gene in leaves,stems and roots of B. oleracea induced by diamond back moth
根据BoTPSa基因序列密码子适应性指数分析,该序列不适合在大肠埃希菌中高效表达(CAI=0.56),所以需要对其密码子进行优化。优化后BoTPSa基因序列的CAI=0.97,说明该序列在大肠埃希菌内能够高效表达目的蛋白。经IPTG 诱导后,未分离的重组目的蛋白在分子质量82 kDa处出现特异性条带,且在分离后的重组蛋白沉淀中发现了相同的条带。说明通过优化密码子以及连接融合标签蛋白的方法,成功使得BoTPSa 在大肠埃希菌中高表达,但是仍然无法实现可溶性表达。在16 ℃、220 r/min摇床振荡16 h和在37 ℃、220 r/min摇床振荡16 h 情况下,依旧为包涵体表达(图5A);且梯度IPTG 浓度诱导结果显示无显著差异(图5B),因此,本试验对包涵体进行复性以获得具有活性的蛋白样品Trx-BoTPSa。
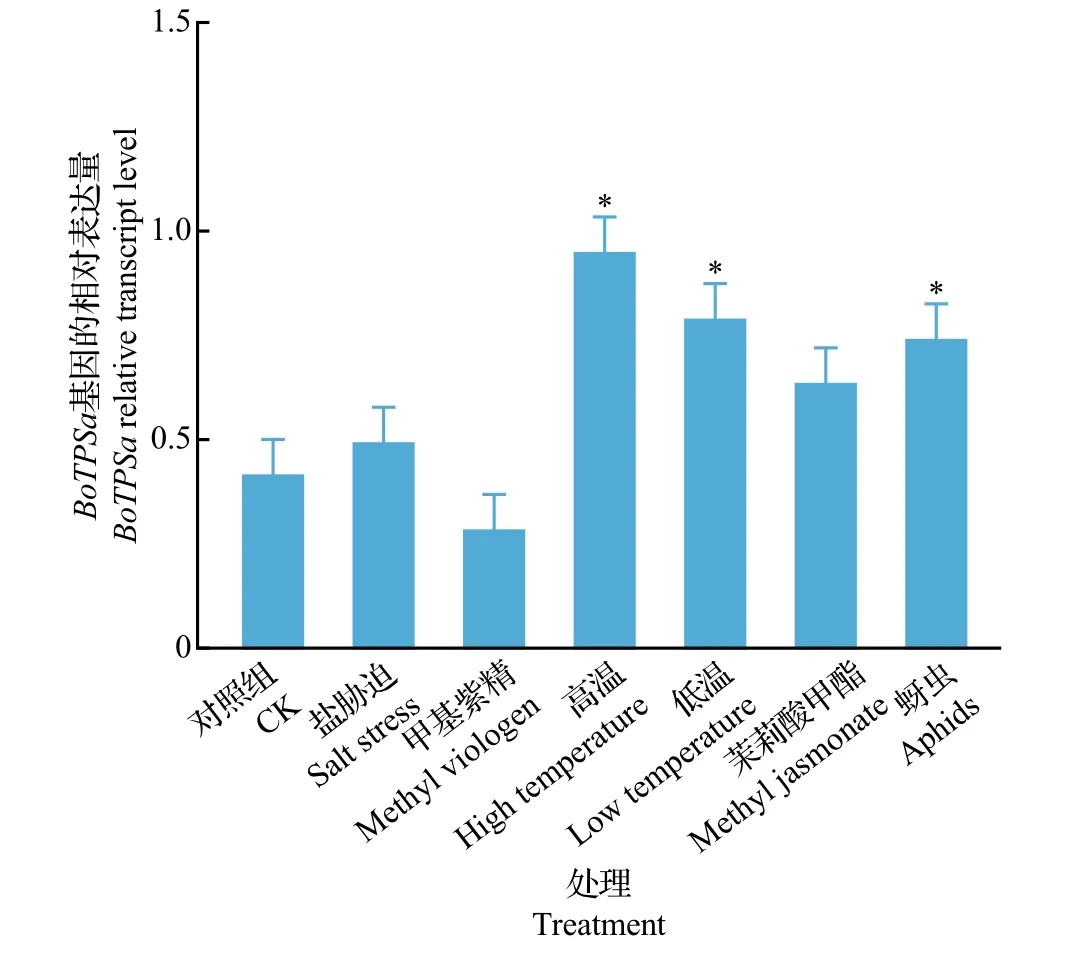
图4 外界因素诱导结球甘蓝叶中BoTPSa基因的相对表达量Fig.4 Relative transcript levels of BoTPSa gene in leaves of B.oleracea induced by external stresses
2.5 结球甘蓝BoTPSa 蛋白的包涵体复性
在37 ℃、0.1 mmol/L IPTG的诱导条件下,大量诱导培养重组Trx-BoTPSa 菌种,破菌后进行包涵体复性以获得具有活性的蛋白样品。对复性样品采用超滤浓缩法(20 kDa超滤膜)进行超滤后,获得特异性条带清晰、质量浓度约1×103mg/L 的Trx-BoTPSa样品(图5C)。
2.6 酶活反应产物气相色谱-质谱分析结果
使用顶空固相微萃取法吸附酶活反应产物,结合GC-MS技术来分析酶活反应液中产生的萜类的种类。用HS-SPME 纤维吸附各样品1 h 后,取出HS-SPME萃取头,在GC-MS仪器上手动进样分析酶活反应产物。获得的质谱图如图6所示。以FPP纯品为对照的样品中无产物峰(图6A),而以稀释1×104倍的法呢烯与烯烃混合液作为标准样品的产物峰图如6B 所示,在提取(93.0±0.7)m/z范围内的离子流后,发现在以FPP 为底物的样品中产生了多个产物峰(图6C)。对比检测到的化合物的保留时间和离子提取峰后发现,甘蓝的BoTPSa 能以FPP为底物生成草烯、β-法呢烯、α-法呢烯。
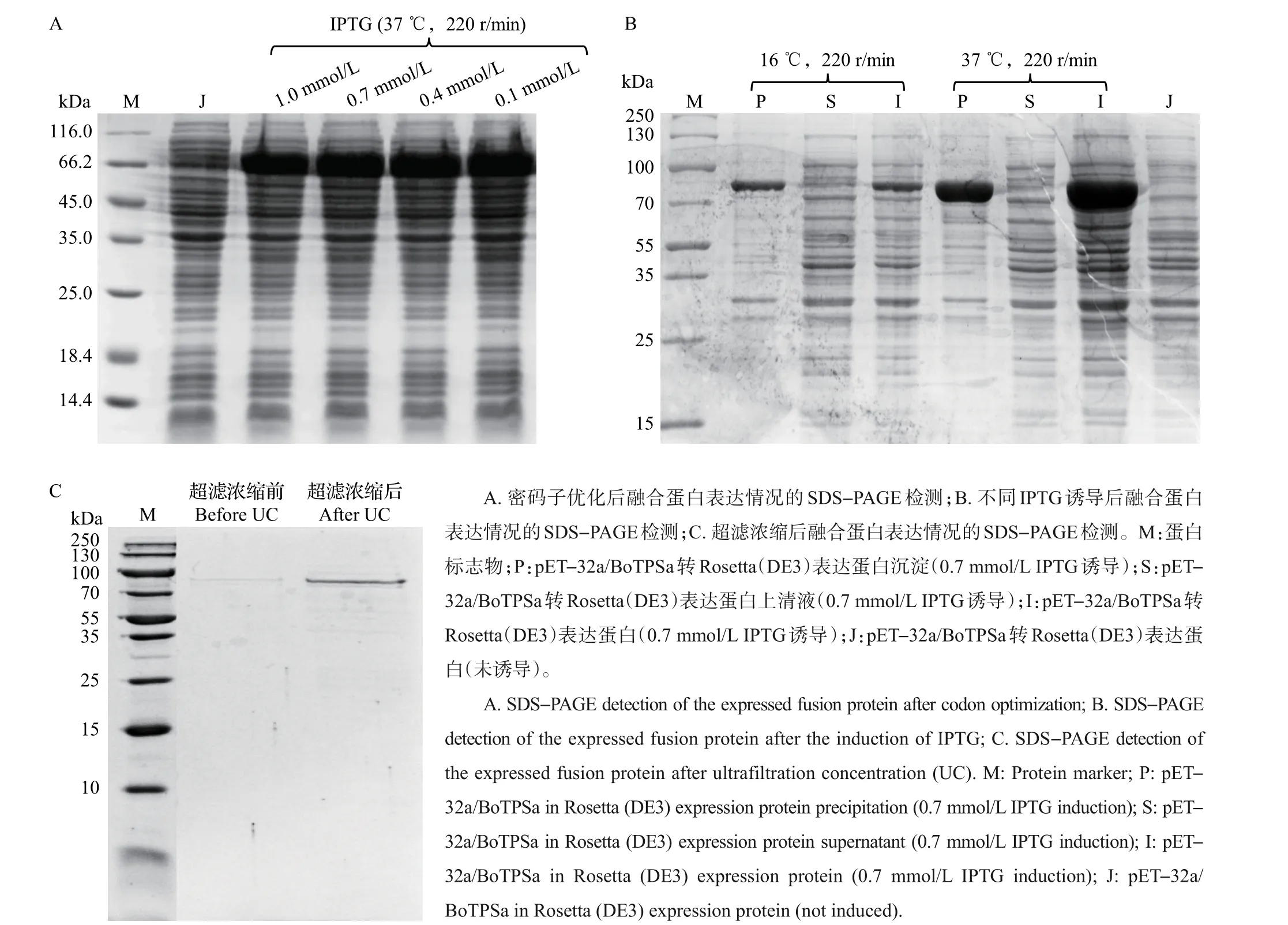
图5 融合蛋白表达的SDS-PAGE检测Fig.5 SDS-PAGE detection of the expressed fusion protein
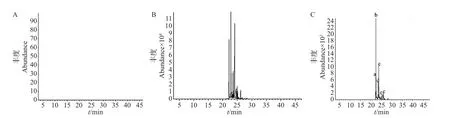
图6 重组BoTPSa酶催化产物的GC-MS分析Fig.6 GC-MS analysis of enzymatic reaction product of recombinant BoTPSa
2.7 BoTPSa 的亚细胞定位分析
通过带绿色荧光蛋白(green fluorescence protein,GFP)标签的目的基因与细胞核标志物基因共转拟南芥原生质体瞬时表达后,在激光共聚焦显微镜下观察到,BoTPSa 荧光与细胞核标志物荧光并不重合(图7A),证明BoTPSa基因不在细胞核中表达。再通过带GFP标签的目的基因转入拟南芥以后,在激光共聚焦显微镜下观测GFP 荧光与叶绿体自发荧光的位置发现,BoTPSa 荧光与标志物荧光并不重合(图7B),说明BoTPSa基因也不在叶绿体中表达。
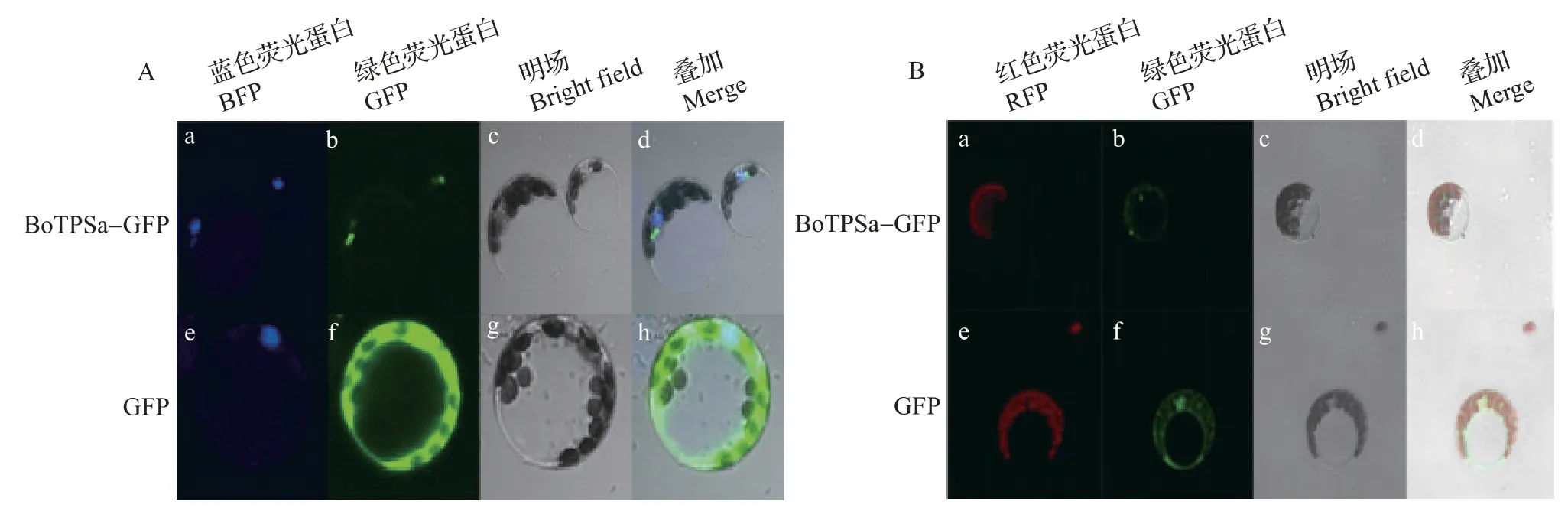
图7 BoTPSa在拟南芥原生质体中的亚细胞定位Fig.7 Subcellular localization of BoTPSa in protoplast of A.thaliana
3 讨论
萜类化合物是自然界中种类最多、结构最多样的一类化合物[21]。萜类合成酶作为植物萜类化合物合成过程中的关键酶,它决定着萜类化合物的结构和种类[22]。随着植物基因组测序的深入发展,学者们能获取植物中萜类合成酶的基因序列,并对其功能特性进行深入解析。目前,虽然萜类合成酶基因已被广泛鉴定,但其在芸薹属植物中的功能研究尚未深入。本研究在结球甘蓝中鉴定出了α-法呢烯合成酶基因BoTPSa,通过预测其结构域,结果发现该蛋白序列含有高度保守的N端结构域和C末端活性结构域,与前人对TPS结构的保守区域的研究结果相符[23]。同时,本研究分析了该基因与其他作物的萜类合成酶基因的亲缘关系,并对它的表达受虫害处理和胁迫处理的影响进行了解析。
随着众多的植物萜类合成酶的生物学功能被相继揭示,其应用也得到了研究者们的关注。然而,植物中萜类合成酶表达水平相对较低、提取步骤复杂、人工提取效率低等问题制约着萜类化合物的广泛应用。而高效率、易改造的原核表达系统为异源合成萜类化合物提供了有效的解决途径。如WONG等[24]研究发现,将外源萜类合成酶基因转入大肠埃希菌可大幅提升其对应的萜类化合物的合成效率。徐娅莹[25]发现,佛手的CmTPSl 重组蛋白能够以FPP为底物生成以双环牻牛儿烯为主的多种倍半萜化合物。本研究利用大肠埃希菌成功异源表达了结球甘蓝BoTPSa 蛋白,并检测发现该酶能以FPP 为底物,催化合成多个倍半萜烯类化合物。其中,β-法呢烯已被发现参与到植物与昆虫的互作中[26],而草烯在结球甘蓝抗虫反应中的作用尚不清楚。
基于体外酶活试验的结果,我们推测结球甘蓝的BoTPSa可能是一个多功能酶。植物的萜类合酶在植物体内可同时介导多个挥发物的合成,这些挥发物通过综合作用来影响植食者,如双条杉天牛(Semanotus bifasciatus)对单一的倍半萜化合物不具趋性,但这些倍半萜的混合物却能引诱大量的双条杉天牛[27],说明萜类化合物之间具有增效效应。所以,我们推测α-雪松烯、γ-依兰烯、没药烯、γ-榄香烯、罗汉柏烯和布藜烯的混合可以增强α-法呢烯调控的结球甘蓝-害虫互作,它们之间具体的联系仍需进一步的探究。当然,由于萜类合酶在植物细胞内的功能与其能获得的底物有关,BoTPSa 在体外和植物体内的酶活产物的类型不一定相同。
萜类合酶在植物体内主要位于细胞质和质体中,分别利用其中的甲羟戊酸途径和甲基赤藓醇4-磷酸途径的产物作为底物来合成不同类型的萜类[28]。因此,萜类合酶在细胞内的分布对于其产物类型有着重要的影响[29]。BoTPSa 亚细胞定位为BoTPSa 蛋白的分子功能解析提供了线索,并为其他萜类合成酶基因功能的解析提供了参考依据。为解析BoTPSa 蛋白在植物细胞内的分布,我们使用拟南芥原生质体悬浮液瞬时表达BoTPSa 蛋白,并对其亚细胞定位进行跟踪,结果发现该BoTPSa不在细胞核和叶绿体中表达,其表达的位置有待后续进一步的研究。
4 结语
本研究结果表明,结球甘蓝BoTPSa基因与十字花科植物拟南芥以及多种芸薹属植物的同源萜类合酶基因在核酸序列水平上高度相似,说明其功能在十字花科植物中可能十分保守。而BoTPSa基因表达受虫害所诱导,在应对非生物胁迫中,本研究发现低温和高温处理均能诱导BoTPSa基因的表达,说明其可能参与抗虫和抗温度胁迫的生态反应。原核表达BoTPSa 蛋白和生化研究发现,该蛋白能催化底物FPP 生成多种倍半萜类挥发物,说明其可能是一个多功能酶。对BoTPSa蛋白的亚细胞定位表明,BoTPSa 蛋白不在细胞核和叶绿体中,其亚细胞定位有待后续研究。本研究为深入解析甘蓝的萜类次生代谢化合物合成机制提供了理论依据。
