环境条件对温和气单胞菌群体感应基因luxI/luxR表达的影响
2021-05-15孙晓佳李婷婷赫彬彬梅永超励建荣
孙晓佳,李婷婷,赫彬彬,梅永超,谢 晶,励建荣*
(1 渤海大学食品科学与工程学院 生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心辽宁省食品安全重点实验室 辽宁省高等学校生鲜食品产业技术研究院 辽宁锦州121013 2 大连民族大学生命科学学院 辽宁大连116600 3 上海海洋大学食品学院 上海201306)
群体感应(Quorum sensing,QS)是细菌细胞间通过分泌信号分子(Autoinducer,AI)进行相互交流的一种通讯机制,当信号分子浓度达到特定阈值时,细胞会启动特定基因的表达,以适应环境的变化,进而调控细菌的生物行为,包括生物发光、生物被膜形成及毒力因子表达等[1-3]。其中,革兰氏阴性菌的QS 调控系统为luxI/R 型,所分泌的信号分子为N-酰基高丝氨酸内酯(N-acyl-homoserine lactones,AHLs)[4]。微生物的生长代谢是引起水产品腐败的主要原因,其QS 系统在水产品的腐败过程中起着至关重要的作用。研究表明,微生物许多特性的表达受温度、pH 值、营养基质等外界环境因素的影响[5]。外界环境的改变、营养基质的差异都可能影响微生物的QS 系统及其致腐能力。食源性假单胞菌具有较强的分泌信号分子的能力,然而,不同温度和碳源条件下分泌的信号分子种类和产量明显不同[6]。孔西曼等[7]发现蜂房哈夫尼菌在不同碳源或氮源培养条件下分泌AHLs 的能力具有显著差异,分别以果糖、硫酸铵为碳源和氮源时AHLs 的分泌量最高。
温和气单胞菌(Aeromonas sobria),又称维隆气单胞菌温和生物型(A.veronii biovar sobria),属于弧菌科、气单胞菌属,广泛存在于植物、动物以及海洋环境中[8]。作为鱼类等水产品中常见的特定腐败菌之一,其毒力因子的产生和生物被膜的形成引起严重的食品安全问题,给水产养殖业带来巨大的经济损失[9-11]。本实验室工作人员从腐败大菱鲆中分离得到1 株温和气单胞菌,发现不同培养条件显著影响其分泌AHLs 的能力[12-13]。此外,有研究表明不同温度条件下蜂房哈夫尼亚菌中QS 调控基因luxI、luxR 的表达量明显不同,低温条件(10 ℃)下其表达量最高,而高于最适温度(28 ℃)时表达量降低[14]。温和气单胞菌在水产品贮藏过程中可能受到各种环境胁迫,如温度、渗透压、pH 值和营养基质的改变。然而,目前关于不同环境条件对温和气单胞菌群体感应系统表达的影响研究较少。
本文通过实时荧光定量PCR 技术检测不同环境条件下(温度、pH 值、NaCl 浓度、碳源、氮源)大菱鲆源温和气单胞菌luxI/luxR 基因(NCBI Reference Sequence:WP_101529659.1;WP_00 5335753.1)的表达情况,旨在探究环境因素对群体感应调控机制的影响,为抑制温和气单胞菌的致腐能力提供理论依据。
1 材料与方法
1.1 试验菌株及培养条件
温和气单胞菌从腐败大菱鲆中分离并鉴定,由本实验室保藏。
1.2 主要试剂与仪器
琼脂糖、DNA Maker、Ezup 柱式细菌基 因组总DNA 提取试剂盒、Taq PCR Master Mix(2×,red dye),上海生工生物工程有限公司;LB 肉汤,青岛海博生物技术有限公司。
Legend Micro21R 台式微量离心机,美国Thermo 公司;MLS-3030CH 立式压力灭菌锅,广州三洋电机有限公司;MS105UD 电子分析天平,瑞士梅特勒-托利仪器有限公司;PCR 仪,德国Eppendorf 公司;Ge1Doc XR+全自动凝胶成像系统、CFX ConnectTM荧光定量PCR 检测系统,美国Bio-Rad 公司。
1.3 方法
1.3.1 菌株活化 将温和气单胞菌过夜活化后,按1∶100 的体积比接种于LB 液体培养基中,28℃,160 r/min 培养12~16 h。
1.3.2 引物设计及合成 以温和气单胞菌16S rRNA 为内参基因,luxI/luxR 为目的基因,使用Primer 5 软件设计引物。将所得引物序列由生工生物工程(上海)股份有限公司合成,引物信息见表1和表2。

表1 温和气单胞菌luxI/luxR 基因引物序列Table 1 The primers of luxI/luxR gene of Aeromonas sobria
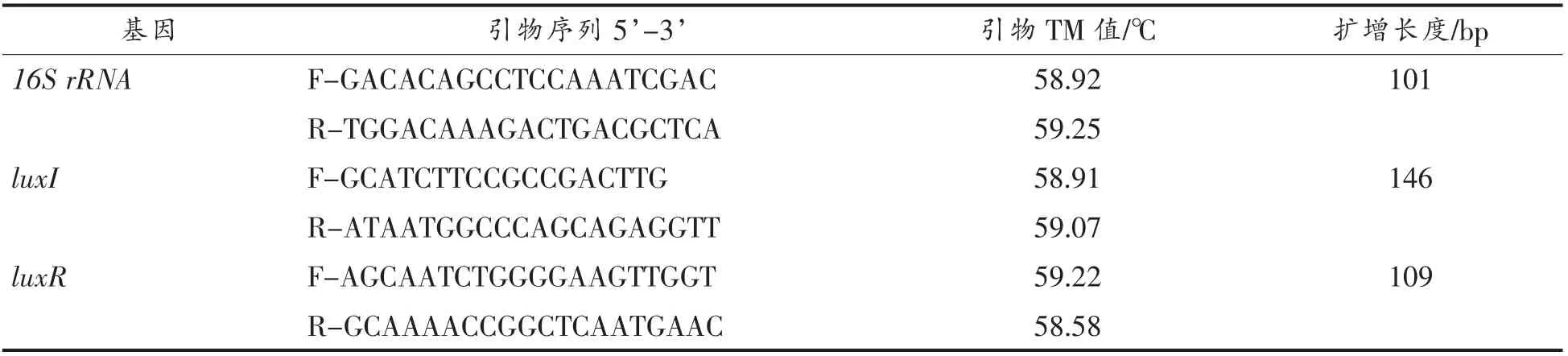
表2 温和气单胞菌luxI/luxR 基因RT-qPCR 引物序列Table 2 The primers of luxI/luxR gene of Aeromonas sobria by RT-qPCR
1.3.3 DNA 提取 使用Ezup 柱式细菌基因组DNA 抽提试剂盒提取细菌总DNA,具体步骤如下:
1)将过夜培养的菌液分装至1.5 mL 离心管,8 000 r/min 室温(25 ℃)离心1 min,弃上清液。
2)加入180 μL 消化缓冲液,再加入20 μL蛋白酶K,振荡混匀,56 ℃水浴1 h 至细胞完全裂解。
3)加入200 μL BD 缓冲液,充分混匀,再加入等量的无水乙醇,颠倒数次混匀。
4)将吸附柱放入收集管中,将上步所得溶液加入吸附柱中,静置2 min,12 000 r/min 室温离心1 min,倒掉收集管中的废液。
5)将吸附柱放回收集管,加入500 μL 含有异丙醇的PW 溶液,10 000 r/min 室温离心30 s,倒掉滤液。
6)将吸附柱放回收集管,加入500 μL 含有无水乙醇的清洗溶液,10 000 r/min 室温离心30 s,倒掉滤液。
7)将吸附柱重新放回收集管中,12 000 r/min室温离心2 min,去除残留的清洗溶液。
8)取出吸附柱,转移到新的离心管中,加入100 μL CE 缓冲液,静置3 min,12 000 r/min 室温离心2 min。提取的DNA 于-20 ℃下保存备用。
1.3.4 引物适用性检测 按上述方法提取温和气单胞菌DNA,以50 μL PCR 体系进行扩增(DNA模板1 μL;上游引物2 μL;下游引物2 μL;Taq PCR Master Mix 25 μL;无菌ddH2O 20 μL)。反应参数为94 ℃预变性4 min,94 ℃变性30 s,58℃退火30 s,72 ℃延伸95 s,共30 个循环,最后72℃延伸10 min。
1.3.5 RNA 提取及质量检测
1)将过夜培养的菌液分装至1.5 mL 离心管,10 000 r/min 离心2 min,弃上清液,加入1 mL Trizol 混匀,室温放置5 min,以充分裂解。
2)加入1/5 体积量的氯仿,摇晃20 s,静置3 min,12 000 r/min 离心15 min(4 ℃),将上层无色液体转移至新的离心管中。
3)加入500 μL 异丙醇,静置10 min,12 000×g 离心10 min(4 ℃),弃上清液。
4)加入1 mL 75%预冷乙醇(DEPC 水配制),颠倒混匀,7 500 r/min 离心5 min(4 ℃),弃上清液,静置5~10 min,即得到纯化的RNA。
1.3.6 cDNA 合成 使用RevertAid First Strand cDNA Synthesis Kit 试剂盒(美国THERMO SCIENTIFIC 公司)将提取的总RNA 转录成cDNA。
第1 步反转录体系为:RNA 模板 1 ng;oligo(dT)18 Primers 2 μL;RNase-free Water 9 μL。反转录条件:65 ℃5 min;4 ℃2 min;4 ℃∞。
第2 步反转录体系为:第1 步RNA 变性溶液12 μL;5X 反应缓冲液4 μL;RiboLock RNA 酶抑制剂1 μL;10 mmol/L dNTP 混合液2 μL;RevertAid M-MuLV 1 μL。反转录条件:42 ℃60 min;70 ℃5 min;4 ℃2 min;4 ℃∞。
1.3.7 荧光定量PCR 将获得的cDNA 溶液进行稀释,以20 μL 体系进行荧光定量PCR 扩增(cDNA 5 μL;上游引物0.5 μL;下游引物0.5 μL;2×SYBR Green PCR Master Mix 10 μL;无 菌ddH2O 4 μL),建立熔解曲线,所得数据跟据方程2-△△Ct计算分析基因表达量的差异。反应参数为95 ℃3 min,95 ℃10 s,55 ℃20 s,72 ℃20 s,于75℃停留5 s,收集荧光信号,共40 个循环并在65~95 ℃范围内建立熔解曲线。
表达量计算公式:
△Ct(试验组)=Ct(目标,试验组)-Ct(内参,试验组)
△Ct(计算)=Ct(目标,计算)-Ct(内参,计算)
△△Ct=△Ct(试验组)-△Ct(计算)
基因表达量=2-△△Ct
1.4 不同环境因素对温和气单胞菌luxI/luxR 基因表达情况的影响
1.4.1 不同温度对温和气单胞菌luxI/luxR 基因表达情况的影响 将温和气单胞菌过夜活化后,按1∶1 000 体积比接种于LB 液体培养基中,分别于不同温度(4,15,28,37 ℃)下,160 r/min 振荡培养24 h。以LB 液体培养基为空白对照组,按照1.3节方法测定luxI 及luxR 基因表达量。
1.4.2 不同pH 值对温和气单胞菌luxI/luxR 基因表达情况的影响 将温和气单胞菌过夜活化后,按1∶1 000 体积比接种于不同pH 值(5.0,6.0,7.0,8.0,9.0)的LB 液体培养基中,28 ℃,160 r/min振荡培养24 h,同上述方法测定luxI 及luxR 基因表达量。
1.4.3 不同碳源对温和气单胞菌luxI/luxR 基因表达情况的影响 参考Zimmer 等[15]方法配制AB培养基,3 g/L K2HPO4,1 g/L NH4Cl,0.3 g/L MgSO4·7H2O,0.15 g/L KCl,1 g/L NaH2PO4,0.01 g/L CaCl2,0.0025 g/L FeSO4·7H2O,酪蛋白水解氨基酸5 g/L,碳源(5 g/L):葡萄糖、木糖、蔗糖、果糖、乳糖和麦芽糖。将温和气单胞菌过夜活化后,按1∶1 000 体积比接种于含有不同碳源的AB 培养基中,28 ℃,160 r/min 振荡培养24 h,同上述方法测定luxI 及luxR 基因表达量。
1.4.4 不同氮源对温和气单胞菌luxI/luxR 基因表达情况的影响 将温和气单胞菌过夜活化后,按1∶1 000 体积比接种于含2%氮源(酵母浸粉、胰蛋白胨、牛肉膏、氯化铵、脲素)的LB 液体培养基中,28 ℃,160 r/min 振荡培养24 h,然后同上述方法测定luxI 及luxR 基因表达量。
1.4.5 不同NaCl 质量分数对温和气单胞菌luxI/luxR 基因表达情况的影响 将温和气单胞菌过夜活化后,按1∶1 000 体积比接种于不同NaCl 质量分数(0.5%,1%,2%,3%,4%,5%)的LB 液体培养基中,28 ℃,160 r/min 振荡培养24 h,同上述方法测定luxI 及luxR 基因表达量。
1.5 数据分析
用SPSS 19.0 软件进行数据分析处理,Origin 8.5 进行绘图处理。
2 结果与分析
2.1 温和气单胞菌群体感应luxI/luxR 基因的验证
利用设计的引物进行PCR 扩增,通过琼脂糖凝胶电泳验证扩增产物。将扩增成功的PCR 产物送至上海生工生物工程股份有限公司测序,然后将所得序列与NCBI 上GeneBank 数据库中的己有序列进行BLAST 比对分析。图1为luxI/luxR 基因琼脂糖凝胶电泳图。从图中可以观察到在预期大小处有明亮且单一的条带,无引物二聚体出现。将测序结果经BLAST 比对后,发现相似度高达99%以上,表明扩增产物与目的序列的一致性。
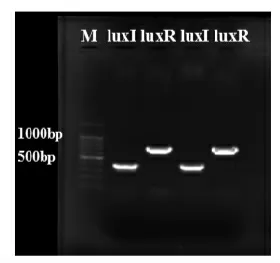
图1 温和气单胞菌luxI/luxR 基因特异性引物PCR 扩增电泳图Fig.1 Agarose electrophoresis of Aeromonas sobria luxI/luxR genes amplification on products for ordinary PCR
2.2 荧光定量PCR 引物检测
2.2.1 引物特异性检测 以cDNA 作为模板进行普通PCR 扩增,通过琼脂糖凝胶电泳验证扩增产物,结果如图2所示,所设计的引物在样品中均扩增出预期大小的片段,条带单一且明亮,无引物二聚体,说明引物特异性良好,可用于后续试验。
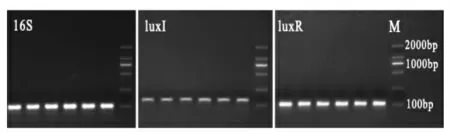
图2 温和气单胞菌luxI/luxR 基因荧光定量引物PCR 扩增电泳图Fig.2 Agarose electrophoresis of Aeromonas sobria genes amplification on products for qPCR
2.2.2 荧光定量PCR 扩增曲线及引物特异性分析 将反转录得到的cDNA 作为模板,使用所设计的引物进行荧光定量PCR 扩增,建立的熔解曲线如图3所示。在荧光定量PCR 试验中,SYBR Green 荧光染料只与双链DNA 结合发出荧光,随着特异性PCR 产物的指数扩增,其荧光信号强度同步增加,而该染料也可能与非特异性扩增产物结合,从而干扰试验结果,因此需要通过对熔解曲线进行分析以确定引物特异性。由图可知,各熔解曲线均只出现了单一的信号峰,没有产生非特异性条带及引物二聚体,说明各引物具有良好的特异性,可进行后续试验。
2.3 不同环境因素对温和气单胞菌luxI/luxR 基因表达情况的影响

图3 温和气单胞菌基因荧光定量PCR 熔解曲线Fig.3 The melting curves of real-time PCR for gnens of Aeromonas sobria
2.3.1 不同温度对温和气单胞菌luxI/luxR 基因表达情况的影响 温度作为调节微生物生长代谢与活力的重要因素,可直接影响细胞的生理功能,温度过低会抑制微生物的生长,过高则会导致死亡[16]。图4所示为不同温度条件下温和气单胞菌luxI/luxR 基因的表达情况。由图可知,温度为37℃时luxI/luxR 的表达量最低,仅为28 ℃条件下(对照组)的49.32%和36.34%。而15 ℃低温条件下luxI/luxR 基因表达量最高,分别为对照组的214.17%和196.72%。此外,在4 ℃冷藏条件下,luxI/luxR 基因的表达量也能达到较高水平。因此,适当的低温条件有利于luxI/luxR 基因的表达,而当温度升高时,2 种基因的表达能力下降。这可能是由于温和气单胞菌是1 种嗜冷菌,低温胁迫刺激了其QS 系统,进而通过调控QS 相关基因的表达来抵抗不利环境。研究表明,副溶血弧菌中III型分泌系统相关毒力基因(vcrD1,vopS,vopD1)在16 ℃时表达量最高,37 ℃时基因表达量有所下降,这与本试验结果类似[17]。
2.3.2 不同pH 值对温和气单胞菌luxI/luxR 基因表达情况的影响 pH 值对维持微生物细胞正常的生长代谢具有重要作用,其影响着细胞膜所带的电荷、酶活性及膜运输蛋白的功能,不同微生物对环境中pH 值的耐受性有所差异[18-19]。如图5所示,相较于其它pH 值条件而言,pH=8 条件下luxI/luxR 基因表达量最高。在pH=5 条件下,luxI/luxR 基因表达量分别为pH=7 条件下(对照组)的39.83%和56.80%,且当pH=9 时luxI/luxR 基因表达量仅为对照组的36.98%和47.24%。结果表明,中性条件对温和气单胞菌luxI/luxR 基因的表达情况影响较小,而酸碱性较强条件下luxI/luxR 基因的表达受到抑制。研究发现,嗜水气单胞菌中溶血素(ahH)、气溶素(aerA)、外膜蛋白(omP)和粘附素(ahA)这4 种毒力基因在中性及偏酸性条件下均可表达,而在碱性条件下只有ahH 基因能够表达[20]。Medina-Martínez 等[21]也报道了气单胞菌属在碱性条件下AHL 分泌量较少且不稳定,均与本研究结果相似。
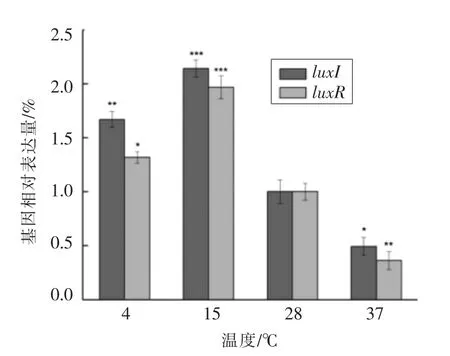
图4 不同温度对温和气单胞菌luxI/luxR 基因相对表达量的影响Fig.4 Effect of different temperature on luxI/luxR gene expression in Aeromonas sobria
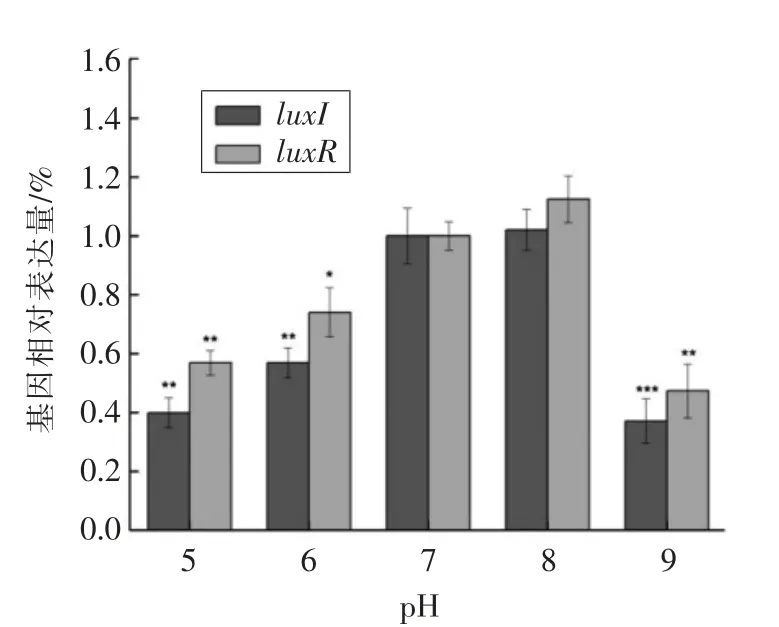
图5 不同pH 值对温和气单胞菌luxI/luxR 基因相对表达量的影响Fig.5 Effect of different pH value on luxI/luxR gene expression in Aeromonas sobria
2.3.3 不同NaCl 质量分数对温和气单胞菌luxI/luxR 基因表达情况的影响 渗透压对于微生物维持正常的生理功能具有重要作用,盐浓度过高会使环境中的渗透压增加,易造成细胞失水及细胞质浓缩,对生长与代谢造成胁迫[22]。因此,本文探究不同NaCl 质量分数对温和气单胞菌luxI/luxR基因表达量的影响。如图6所示,NaCl 质量分数为0.5%时,luxI/luxR 基因表达量最高。随着NaCl质量分数逐渐增大,luxI/luxR 基因表达量逐渐下降。当NaCl 质量分数为5%时,luxI/luxR 基因表达量最低,仅为NaCl 质量分数1%时(对照组)的27.04%和12.04%。由此可知,盐浓度能够影响温和气单胞菌luxI 及luxR 基因的表达情况,且低盐浓度条件有利于2 种基因的表达,而高盐浓度会抑制2 种基因的表达。研究发现,单增李斯特菌中毒力基因inlB 在NaCl 质量分数范围为1.5%~2.0%时可高效表达,而NaCl 质量分数为3.5%以上毒力基因inlB 的表达受到抑制,这与本研究结果相似[23]。
2.3.4 不同碳源对温和气单胞菌luxI/luxR 基因表达情况的影响 碳源是微生物生命活动中不可缺少的物质,为细胞的生长代谢提供所需的能量[24]。因此,本文研究不同碳源对温和气单胞菌群体感应luxI/luxR 基因表达情况的影响。如图7所示,葡萄糖为碳源时,luxI/luxR 基因表达量最高,分别为260.90%和145.34%。而当木糖为碳源时,luxI/luxR 基因表达量最低,仅为22.34%和19.89%。其中,luxI 基因对不同碳源的利用能力依次为:葡萄糖>蔗糖>麦芽糖>乳糖>果糖>木糖;luxR 基因对不同碳源的利用能力依次为:葡萄糖>乳糖>蔗糖>麦芽糖>果糖>木糖。因此,不同碳源能够影响温和气单胞菌luxI/luxR 基因的表达,且对luxI 基因表达量的影响较为明显。本实验室前期探究了不同碳源对温和气单胞菌分泌AHLs 的影响,结果发现以葡萄糖、蔗糖和麦芽糖为碳源时,分泌AHLs 能力最强,与本研究结果呈一致性[13]。

图6 不同氯化钠质量分数对温和气单胞菌luxI/luxR基因相对表达量的影响Fig.6 Effect of different sodium chloride mass fractions on luxI/luxR gene expression in Aeromonas sobria
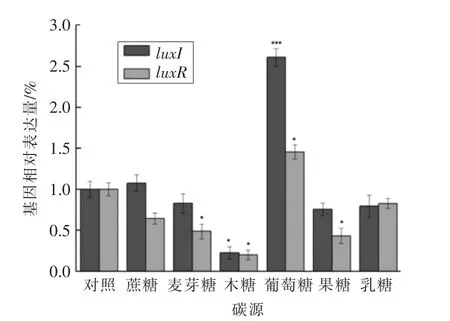
图7 不同碳源对温和气单胞菌luxI/luxR 基因相对表达量的影响Fig.7 Effect of different carbon sources on luxI/luxR gene expression in Aeromonas sobria
2.3.5 不同氮源对温和气单胞菌luxI/luxR 基因表达情况的影响 氮源作为微生物生长代谢的基础营养物质,可以为细胞生长提供能源,不同细菌对不同氮源的利用率有所差异[25]。图8所示为不同氮源对温和气单胞菌luxI/luxR 基因表达情况的影响。从图中可以看出,当胰蛋白胨为氮源时,luxI/luxR 基因相对表达量达到最高,分别为150.23%和208.89%。而当脲素为氮源时,luxI/luxR 基因相对表达量最低,分别为42.40%和22.51%。其中,luxI 基因对不同氮源的利用能力依次为:胰蛋白胨>氯化铵>牛肉膏>酵母浸粉>脲素;luxR 基因对不同氮源的利用能力依次为:胰蛋白胨>氯化铵>酵母浸粉>牛肉膏>脲素。由此可知,胰蛋白胨为温和气单胞菌培养的最佳氮源,可明显促进luxI/luxR 基因的表达能力。研究发现,嗜水气单胞菌以胰蛋白胨为氮源时,其毒力因子胞外蛋白酶的产量最高,这与本研究所得结果类似[26]。
3 结论

图8 不同氮源对温和气单胞菌luxI/luxR 基因相对表达量的影响Fig.8 Effect of different nitrogen sources on luxI/luxR gene expression in Aeromonas sobria
本研究以温和气单胞菌QS 调控基因luxI/luxR 为研究对象,探究了不同环境条件对该基因表达情况的影响。结果发现,温和气单胞菌luxI/R系统的表达情况受环境因素调控,当受到环境胁迫时,可通过QS 系统调节其生物行为。适当低温(15 ℃)及弱碱(pH=8)条件有利于luxI/luxR 基因的表达,而高温(37 ℃)及强酸碱性(pH=5 或9)使2 种基因的表达受到抑制。此外,随着NaCl 质量分数升高,2 种基因的表达量呈下降趋势。不同碳源条件下luxI/luxR 基因的表达情况明显不同,其中在以葡萄糖为碳源时,2 种基因的表达水平最高,且luxI 基因对碳源的敏感程度显著高于luxR基因。不同氮源条件下luxI 基因表达情况依次为胰蛋白胨>氯化铵>牛肉膏>酵母浸粉>脲素,而luxR 基因表达情况依次为胰蛋白胨>氯化铵>酵母浸粉>牛肉膏>脲素。本文主要研究了环境条件对温和气单胞菌QS 相关基因luxI/luxR 表达情况的影响,为进一步探究环境因素对温和气单胞菌QS 系统的调节作用提供理论基础。
