D型短指症与HOXD13基因的相关研究进展
2021-05-08刘通陈宇许强姬广林
刘通 陈宇 许强 姬广林

【关键词】 D型短指;球拍拇指;HOXD13基因
中图分类号:R726.2 文献标志码:A DOI:10.3969/j.issn.1003-1383.2021.03.014
短指症(brachydactyly,BD)是一类由于指骨或掌骨发育异常导致的指骨缩短畸形,临床上患病率为0.41%~4.0%[1]。其中以D型短指(brachydactyly type D,BDD)最为常见,因拇指甲板和甲床的宽度大于长度,形状短而宽,而余指体不受累,故又称为球拍拇指(racquet thumbs)[2]。常表现为末节指骨发育不良,伴指间关节活动轻度受限。近年来,随着发病率的升高,BDD越来越受到人們的重视,研究显示HOXD13基因突变参与了BDD的形成。HOX基因属于同源盒家族的一员,通常成簇分布、高度保守,D簇是肢体发育的关键,它参与发育过程中末节肢体的分化和形态发生[3~4]。但相关的致病机制尚不明确,因此BDD有关分子水平的研究在单基因病家系中有着重要的意义。现对BD分类、流行病学,BDD分子遗传学特点、与HOXD13基因的联系及治疗等研究进行综述,以期为人类指骨发育的分子调控机制研究及治疗提供参考。
1 BD分类及流行病学
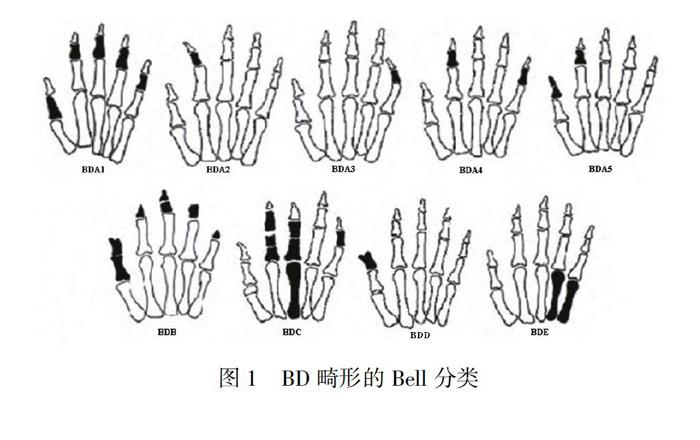
根据临床表型,可将BD分为单纯型BD以及包含BD的综合征,又因畸形发生部位和受累程度的不同,Bell将单纯型BD分为A、B、C、D和E等5种类型(图1)[1]。其中A型中指骨短;B型中指骨短且伴有指骨末端发育不全或缺如、拇指畸形、指关节粘连、并指等,为BD中最严重的一类;C型第Ⅱ、Ⅲ指中指骨及近端指骨异常,近端指骨分节过多;D型拇指的末端指骨短而宽;E型由于掌骨变短形成,部分患者可伴其他骨骼异常。其中A型又分为5个亚型,A1最为典型,表现为所有指骨的中指节缩短伴第1指的近端指节缩短,并出现中指节和远端指节融合的现象,又称Farabee型短指症[5]。其次,BD可出现在不同种族背景的人群中。其中D型和A型最为常见,其他类型相较少见[6]。但它们的发生率在不同族群之间有着极大的差异,D型的患病率为0.41%~4.0%,其中以日本人和以色列阿拉伯人患病率最高;A型的患病率为3.4%~21%,在美国印第安人和蒙古人中更为常见[7]。此外,我国人群中也有相应的BD家系的报道[8]。
2 BDD分子遗传学特点及相关的综合征
BD最先于1903年被发现,HALL等[9]研究证实其符合孟德尔遗传定律,通常发生于胚芽形成时(约胚胎第8周)。BDD通常以常染色体显性的方式进行遗传,并且女性外显完全而男性外显不完全,大约3/4的患者表现为双手拇指对称性短指畸形[10]。这种末节指骨的先天性畸形是由于骨膜持续生长过程中,指骨的骨骺线过早的骨化引起了闭塞,同时又失去了侧指甲的褶皱,导致短而宽的指骨覆盖于已成型的指甲板,引起BDD的发生[12]。致病机制方面,HOXD13基因中聚丙氨酸链的短缩导致的错义突变可诱发BDD及相关的综合征的出现[11]。此外,位于第20号染色体q13.3上的GNAS基因突变也参与构建人体先天性肢端畸形[12~13]。郭芮吉等[14]分别应用全外显子测序和Sanger测序对我国3名BDD/BDE患者及其家系进行基因突变检测,结果两者方法检测结果一致,提示3名患者均存在GNAS基因13号外显子上c.A1145T杂合错义突变,再次验证了GNAS基因突变导致BDD存在于某些家系中。一些与BDD相关的综合征如Rubinstein-Taybi综合征[15]、Poland综合征[16]、Robinow综合征[17]如表1所示。
3 HOXD13及相关疾病的研究
HOX基因编码一个高度保守的转录因子家系,这些转录因子在胚胎发育中起着重要作用[13]。人类HOX基因至少有39个,分为A、B、C、D四簇,依次位于染色体7pl4、17q21、12q13和 2q31,每个基因簇含9~12个基因,长度大约120 kDa,其中D簇是脊椎动物肢体发育的关键,位于该簇端粒和中心粒基因两侧的增强子调节着HOXD13基因的转录[3,18]。HOXD13基因编码同源异型结构域转录因子,与肢体生长发育有着紧密的联系,其定位于2q31.1(HOXD基因簇的5的最末端),通常含两个外显子,全长1365 bp,编码区长1008 bp,编码的蛋白质由335个氨基酸组成。其产物在3端是一个180 bp的同源盒功能域,5端则含两个聚丝氨酸链和一个聚丙氨酸链[19]。按照5端→3端方向进行转录,在每个基因簇中的基因其自然位置与发育过程中的表达方式相对应,在特定的位置依次进行表达或沉默。因此接近尾端(5端)的HOXD13基因一般较晚表达[20~21]。HOXD13基因常表现于体轴远端及神经外胚层末端,“掌管”着早期胚胎肢芽和指骨结构的发育,亦是造成并多指畸形(synpolydactyly,SPD)、BDD、BDE的重要因素[22~23]。此外,HOXD13基因还参与其他系统的形成,KAJIOKA等[24]通过原位杂交检测出HOXD13在生殖结节间质中表达,参与外生殖器的形成。WIESEMANN等[6]报道了一例同时患有SPD的母子。母亲在HOXD13基因中有一个新的移码突变(c.708 - 708delc),在胎儿中也发现了这种突变,胎儿的染色体微阵列是正常的,胎儿除了SPD外还伴有先天性脐膨出。但HOXD13移码突变与SPD胎儿脐膨出之间的联系尚不明确,有待进一步研究证实。同时,HOXD13基因的甲基化参与了部分肿瘤的进展,如乳腺癌和胆管细胞癌[25~26]。此外,该基因的表达水平与肿瘤的发病率呈正相关,这一点在散发性乳腺癌患者的低生存率、预后方面的研究中得以证实[27~28]。
4 BDD与HOXD13的联系
如前所述,HOXD13是编码与肢体末端生长发育相关的同源异型结构域转录因子基因。BDD均可由HOXD13基因突变引起。NELSON等[29]通过鸡的动物实验发现HOXD13基因的突变在四肢形成期得到了相应的表达,这种突变包括错义及无义突变等。短指畸形大部分是由HOXD13基因中聚丙氨酸链的延展或缩短所造成,聚丙氨酸链结构在转录因子中广泛存在,可进行转录调控或介导蛋白质间的相互作用[30~31]。短链聚丙氨酸链(7个丙氨酸残基)形成α螺旋,长链(>10个丙氨酸残基)形成β折叠[32]。聚丙氨酸链的延展或缩短可阻碍HOXD13基因与转录因子的相互作用,进一步诱发蛋白质的快速加工过程[33]。聚丙氨酸链延展参与了SPD的形成,而聚丙氨酸链的缩短所致的错义和无义突变均可引起BDD及相关的综合征,并与NOTCH信号通路共同参与诱导骨的增殖、分化[34]。以下就分基因突变和信号通路两个方面阐述BDD与HOXD13基因的联系。
4.1 HOXD13基因的突变
JOHNSON等[35]在对HOXD13基因的研究中发现两种位于HOXD13基因同源盒的错义突变:c. 940 A > C(p.I314L)和c. 923 C > G(p.S308C),均产生于HOXD13基因的第二外显子同源结构域,其中位于同源结构域第47位的亮氨酸对DNA结合引起了BDD的发生。c. 940 A > C突变的患者第Ⅲ掌骨短缩、第Ⅴ指并指,同样的突变在一个连续Ⅵ代出现短指和多指的家族中也被检出[36];c. 923 C >G突变产生于一个早期报道为BDD的家族,家族中大多数患者掌骨缩短同时伴有拇指远节指骨的缩短[37]。通過体外DNA-蛋白质结合试验显示:与野生型HOXD13蛋白相比,发生c. 923 C >G突变的蛋白结合DNA的能力并未出现明显的改变,而是其突变的HOXD13蛋白与其核心结合序列5-TTAC-3的亲和力增强,与结合序列5-TTAT-3的亲和能力减弱,这种复合效应可以由引起改变的氨基酸侧链上甲基化位点与相关碱基之间相互作用,从而引起BDD的发生[23]。ZHAO等[33]在一个短指畸形家族中发现由HOXD13聚丙氨酸链缩短导致的错义突变,该家族所有的肢端畸形呈现对称性分布,并显示与BDD和SPD密切相关,BROWN等[30]用两点连锁分析法将其致病基因定位于D2S1379-D2S324,5HOXD基因测序指出,HOXD13基因中聚丙氨酸链发生缩短突变:由正常的15个丙氨酸缩短为8个。先前有研究认为HOXD13基因聚丙氨酸链在一定范围之内的延长或缩短是一种多态现象,但当缺失的丙氨酸数目>7个就可能引起肢端畸形[33]。随后KUSS等[38]在小鼠相关模型中进一步提出HOXD13基因的突变经视黄醇通道下调了视黄酸的水平,诱导指状组合型软骨的生成,从而改变了正常的排列结构。在对一短指畸形家系的研究中发现,患者携带有HOXD13基因内的一个无义突变c.820C > T(p.R274X)[39]。
4.2 NOTCH信号通路
NOTCH信号通路对早期胚胎发育具有极大影响,也参与诱导骨的增殖、分化,是调控骨代谢的重要途径之一。通过荧光素酶报告系统和实时定量PCR发现HOXD13基因的过表达会引起CHSY1基因的上调[40]。CHSY1基因是通过调控NOTCH信号通路影响软骨的发育[41]。早期在斑马鱼动物实验得出:骨骼和胸鳍的发育过度生长,表明CHSY1基因的缺失会导致NOTCH信号通路上调;WILSON等[42]用同样的方法在小鼠上观察到软骨发育不良、骨密度下降、远节指间关节移位致使BDD等现象,进一步证明了NOTCH信号通路在指骨发育中的重要作用,也证实了CHSY1与HOXD13基因有着密切的关系,但究竟以何种方式相联系,还需要进一步研究[43~44]。
5 BDD的防治措施
目前BDD主要依赖超声进行筛查,然而肢端发育的异常在胚胎早期很难进行观察及干预。现今对于该类畸形最好的办法还是在妊娠期采用超声技术进行排查。对于生育的畸形患儿可以采用外科技术及义肢进行修复或弥补,但治疗方式现今尚无统一的标准,对于单纯短指一般不需特殊处理,但如果严重影响了手指的功能或伴有并指畸形,则需要手术进一步有效干预,手术一般选择两岁后进行。手术的目的一是为了恢复手的功能;二是改善形态,并尽量避免或减轻后遗症,目前有指骨延长[45]、植骨成形[46]、修复重建[47]等治疗方法,对伴有SPD的患者可采用分指术,很大程度可以缓解畸形,让患者投入到正常的生活和工作中。随着科技的进步,近年来提出的仿生技术(丙烯酸材料假指关节),可根据患者手指正常生理长度及外形定制生物型仿生手指,加上当今3D打印技术及神经科学的发展,人工生物肢体技术将有可能在BDD的治疗中发挥着重要作用[48]。同时,精准医学的理论构建也有了新的支持,这无疑提供了一种新的治疗理念,开拓了新的思路。
6 展望
综上,指骨发育是一个动态的过程,受到基因及信号通路的调控。HOXD13作为BDD发生的关键基因,其作用机制、分子水平的探索研究,对了解遗传性肢体畸形有着极其重要的意义。治疗方面,再生医学展示了良好的前景,满足精准医疗的要求。但如果基因检测技术应用于胚胎发育的初期,使其得到相关的检测及早期的干预,将有助于指导此类患者的合理治疗。随着BDD在分子水平上的研究,指骨发育的过程有望得到进一步细致的探索,并且可以通过更多分子水平上的手段得到更多对其治疗的新思路和新方法;研究方面,对可能具有致病基因的突变进行功能性的实验研究,基因筛查对遗传病因的鉴定,为进一步建立有效的干预手段提供基础的同时,也为临床表型分类和遗传咨询提供了精确的科学依据。
参 考 文 献
[1] TEMTAMY S A,AGLAN M S.Brachydactyly[J].Orphanet J Rare Dis,2008,3:15.
[2] RICHERT B,ANDR J.Nail disorders in children:diagnosis and management[J].Am J Clin Dermatol,2011,12(2):101-112.
[3] LE CAIGNEC C,PICHON O,BRIAND A,et al.Fryns type mesomelic dysplasia of the upper limbs caused by inverted duplications of the HOXD gene cluster[J].Eur J Hum Genet,2020,28(3):324-332.
[4] PRIMON M,HUNTER K D,PANDHA H S,et al.Kinase regulation of HOX transcription factors[J].Cancers (Basel),2019,11(4):E508.
[5] GAO B,GUO J,SHE C,et al.Mutations in IHH,encoding Indian hedgehog,cause brachydactyly type A-1[J].Nat Genet,2001,28(4):386-388.
[6] RADHAKRISHNAN P,NAYAK S S,PAI M V,et al.Occurrence of synpolydactyly and omphalocele in a fetus with a HOXD13 mutation[J].J Pediatr Genet,2017,6(3):194-197.
[7] MUNDLOS S.The brachydactylies:a molecular disease family[J].Clin Genet,2009,76(2):123-136.
[8] 王旭,沙艳伟,梅利斌,等.一短指畸形家系临床特征调查及ROR2基因突变分析[J].中国优生与遗传杂志,2017,25(1):104-106.
[9] HALL C M.International nosology and classification of constitutional disorders of bone (2001)[J].Am J Med Genet,2002,113(1):65-77.
[10] DATHE K, KJAER K W, BREHM A, et al. Duplications involving a conserved regulatory element downstream of BMP2 are associated with brachydactyly type A2[J].AM J Hum Genet,2009,84(4):483-492.
[11] GUO X Y,SHI T F,LIN M R,et al.A nonsense mutation in HOXD13 gene from A Chinese family with non-syndromic synpolydactyly[J].Tohoku J Exp Med,2019,249(2):93-100.
[12] USARDI A,MAMOUNE A,NATTES E,et al.Progressive development of PTH resistance in patients with inactivating mutations on the maternal allele of GNAS[J].J Clin Endocrinol Metab,2017,102(6):1844-1850.
[13] KRUMLAUF R.Hox genes,clusters and collinearity[J].Int J Dev Biol,2018,62(11-12):659-663.
[14] 郭芮吉,韓刚,方霞,等.GNAS新错义突变导致D/E型短指畸形[J].组织工程与重建外科杂志,2019,15(3):152-154.
[15] YU S,WU B B,QIAN Y Y,et al.Clinical exome sequencing identifies novel CREBBP variants in 18 Chinese Rubinstein-Taybi Syndrome kids with high frequency of polydactyly[J].Mol Genet Genomic Med,2019,7(12):e1009.
[16] PRATYUSH S, APOORV, AHMER Z, et al.Brachysyndactyly in Poland Syndrome[J].Cureus,2020,12(8):e9755.
[17] CHEN J,MIAO Y,XIE Y,et al.EP17.09:Prenatal diagnosis of a fetus with Robinow syndrome[J].Ultrasound Obstet Gynecol,2019,54(S1):336.
[18] SONG J Y,PINEAULT K M,DONES J M,et al.Hox genes maintain critical roles in the adult skeleton[J].Proc Natl Acad Sci USA,2020,117(13):7296-7304.
[19] BRISON N,DEBEER P,TYLZANOWSKI P.Joining the fingers:a HOXD13 Story[J].Dev Dyn,2014,243(1):37-48.
[20] KONDO T,DUBOULE D.Breaking colinearity in the mouse HoxD complex[J].Cell,1999,97(3):407-417.
[21] BRUNETTI L,GUNDRY M C,SORCINI D,et al.Mutant NPM1 maintains the leukemic state through HOX expression[J].Cancer Cell,2018,34(3):499-512.e9.
[22] HUANG J,LIU H Y,WANG R R,et al.A 3.06-Mb interstitial deletion on 12p11.22-12.1 caused brachydactyly type E combined with pectus carinatum[J].Chin Med J (Engl),2019,132(14):1681-1688.
[23] ZAIB T, WEI J, KOMAL S. A heterozygous duplication variant of the HOXD13 gene caused synpolydactyly type 1 with variable expressivity in a Chinese family[J].Bmc Medical Genetics, 2019,20(1):203.
[24] KAJIOKA D,SUZUKI K,NAKADA S,et al.Bmp4 is an essential growth factor for the initiation of genital tubercle (GT) outgrowth[J].Congenit Anom (Kyoto),2020,60(1):15-21.
[25] KE D, RONG Y, LINA J.Combined diagnosis of breast cancer in the early stage by MRI and detection of gene expression[J]. Experimental & Therapeutic Medicine, 2018,16(2):467-472.
[26] PACO A,DE BESSA G S,FREITAS R.Methylation in HOX clusters and Its applications in cancer therapy[J].Cells, 2020,9(7):1613.
[27] ZHONG Z B ,SHAN M, QIAN C, et al.Prognostic significance of HOXD13 expression in human breast cancer[J]. International Journal of Clinical & Experimental Pathology, 2015, 8(9):11407-11413.
[28] DE BESSA GARCIA S A,ARA′UJO M,PEREIRA T,et al.HOX genes function in breast cancer development[J].Biochim Biophys Acta Rev Cancer,2020,1873(2):188358.
[29] NELSON C E,MORGAN B A,BURKE A C,et al.Analysis of Hox gene expression in the chick limb bud[J].Development,1996,122(5):1449-1466.
[30] BROWN L Y,BROWN S A.Alanine tracts:the expanding story of human illness and trinucleotide repeats[J].Trends Genet,2004,20(1):51-58.
[31] CATALFO P,MUSUMECI M L,LACARRUBBA F,et al.Congenital malalignment of the great toenails:a review[J].Skin Appendage Disord,2018,4(4):230-235.
[32] BLONDELLE S E,FOROOD B,HOUGHTEN R A,et al.Polyalanine-based peptides as models for self-associated beta-pleated-sheet complexes[J].Biochemistry,1997,36(27):8393-8400.
[33] ZHAO X L,SUN M,ZHAO J,et al.Mutations in HOXD13 underlie syndactyly type V and a novel brachydactyly-syndactyly syndrome[J].Am J Hum Genet,2007,80(2):361-371.
[34] 呂赵劼,王志浩,卢淑娴,等.短指(趾)症及指(趾)骨发育的分子调控机制[J].遗传,2019,41(12):1073-1083.
[35] JOHNSON D,KAN S H,OLDRIDGE M,et al.Missense mutations in the homeodomain of HOXD13 are associated with brachydactyly types D and E[J].Am J Hum Genet,2003,72(4):984-997.
[36] CARONIA G,GOODMAN F R,MCKEOWN C M,et al.An I47L substitution in the HOXD13 homeodomain causes a novel human limb malformation by producing a selective loss of function[J].Development,2003,130(8):1701-1712.
[37] MAN N,NIMER S D.p300 suppresses leukemia development in NUP98-HOXD13 driven myelodysplastic syndrome[J].Oncotarget,2018,9(42):26603-26604.
[38] KUSS P,VILLAVICENCIO-LORINI P,WITTE F,et al.Mutant HOXD13 induces extra digits in a mouse model of synpolydactyly directly and by decreasing retinoic acid synthesis[J].J Clin Invest,2009,119(1):146-156.
[39] JAMSHEER A,SOWI?倰[1]SKA A,KACZMAREK L,et al.Isolated brachydactyly type E caused by a HOXD13 nonsense mutation:a case report[J].BMC Med Genet,2012,13:4.
[40] GUO M,LIU Z,WILLEN J,et al.Epigenetic profiling of growth plate chondrocytes sheds insight into regulatory genetic variation influencing height[J].Elife,2017,6:e29329.
[41] TIAN J,LING L,SHBOUL M,et al.Loss of CHSY1,a secreted FRINGE enzyme,causes syndromic brachydactyly in humans via increased NOTCH signaling[J].Am J Hum Genet,2010,87(6):768-778.
[42] WILSON D G,PHAMLUONG K,LIN W Y,et al.Chondroitin sulfate synthase 1 (Chsy1) is required for bone development and digit patterning[J].Dev Biol,2012,363(2):413-425.
[43] LUO Z,RHIE S K,FARNHAM P J.The enigmatic HOX genes:can we crack their code?[J].Cancers(Basel),2019,11(3):323.
[44] SHIELDS B J,SLAPE C I,VO N,et al.The NUP98-HOXD13 fusion oncogene induces thymocyte self-renewal via Lmo2/Lyl1[J].Leukemia,2019,33(8):1868-1880.
[45] DANILKIN M Y.Phalangeal lengthening techniques for brachydactily and posttraumatic digital stumps with the use of a modified external mini-fixator[J].Tech Hand Up Extrem Surg,2016,20(2):61-66.
[46] 楊茜,王斌.先天性短指畸形的发病机制、分类及治疗进展[J].组织工程与重建外科杂志,2015,11(6):389-395.
[47] DU Z J,CUI Y,HAO J,et al.New technique for correcting bony deformity in racquet thumb[J].Ann Plast Surg,2020,84(4):390-393.
[48] AMSUESS S,GOEBEL P,GRAIMANN B,et al.A multi-class proportional myocontrol algorithm for upper limb prosthesis control:validation in real-life scenarios on amputees[J].IEEE Trans Neural Syst Rehabil Eng,2015,23(5):827-836.
(收稿日期:2021-01-09 修回日期:2021-01-26)
(编辑:梁明佩)
