低温胁迫对大豆花期叶片蔗糖代谢及产量的影响
2021-04-27赵晶晶郑殿峰
赵晶晶,周 浓,郑殿峰
(1重庆三峡学院生物与食品工程学院,三峡库区道地药材绿色种植与深加工重庆市工程实验室,重庆404120;2广东海洋大学,广东湛江524088)
0 引言
在制约植物地理分布和作物生产力的诸多因素中,低温胁迫是最重要的一项[1]。东北平原是中国最重要的大豆产区,即使在全球变暖的背景下,低温胁迫仍是东北平原作物生产的主要非生物胁迫,会影响作物的正常生长发育和产量形成[2-3]。因此,研究低温胁迫对大豆花期叶片蔗糖代谢及产量的影响,对指导东北地区大豆生产实践具有重要的意义。
碳代谢是指作物体内有机物质合成、转化和降解的代谢过程[4],该过程能够为作物新组织提供所需的碳源[5],其代谢强度在作物生长发育过程中的动态变化直接影响着光合产物形成、运输以及矿质营养吸收等[6]。大豆光合产物以淀粉的形式储存在叶片中,夜间经碳代谢作用分解为蔗糖运输到各器官中为新组织提供所需的碳源,故整个碳水化合物的合成和水解过程都需要多种酶的参与。转化酶和蔗糖合成酶(SS)可以将蔗糖水解为己糖进入各种代谢途径[7],从而完成物质在源-库器官之间的代谢转运过程。Erdal[8]发现短暂的盐胁迫会使玉米叶片内淀粉和蔗糖含量迅速下降,葡萄糖和果糖含量升高,叶片内合成酶活性降低、分解酶活性升高[9],致使蔗糖的生物合成过程受阻。前人对于大豆苗期[10]、花期[11]、结荚期[12]和籽粒充实期[13]的低温胁迫已取得了一定的研究成果。产量是衡量作物生长状况的重要指标,也是作物对非生物胁迫响应的综合体现,Matsukawa[13]以及Yamamoto和Narikawa[14]认为低温胁迫致使豆类作物减产的原因主要概括为三个方面:(1)豆科作物苗期发生低温胁迫会延缓其前期的生长发育,致使植株长势弱小,叶片出现萎蔫脱水现象;(2)在豆科作物的花期发生低温胁迫会影响叶片光合产物的输出,使总状花序的花和荚不能从叶片(源器官)得到充足的光合同化物,致使顶端的花荚发育不良,甚至大量脱落;(3)籽粒充实期发生低温胁迫会造成植株的籽粒充实不良,这就是低温胁迫造成豆科作物减产的主要原因。荚数是豆类作物高产的关键因素,因为单株荚数直接影响其产量,间接影响单株籽粒数,但其对环境胁迫十分敏感,如低温、干旱、淹水或高温等非生物胁迫均不同程度地影响其数量的多少[15]。王萍等[16-17]以3个不同熟期大豆品种为试验材料,发现大豆花期遭遇低温胁迫会使产量降低的主要原因是三粒荚数和单株荚数减少,导致单株粒数下降,并且恒温低温处理对大豆单株荚数和单株粒数形成的影响远大于变温低温处理。Funatsuki等[18]发现花期冷害胁迫可以使耐寒、中度敏感和冷害敏感品种的产量依次减少35.56%、55.00%和67.34%,花期冷害除了导致豆科作物的荚数减少,还会导致子粒发育不良,空瘪荚数增加。Bernacchi等[19]和Morgan等[20]认为,源和库共同控制作物的产量形成,源活性的增强需伴随库强度潜力的增加,保证光合源(叶片)具有较强的光合物质生产能力是大豆获得高产的前提,花期是决定大豆产量高低的关键生育期,而关于低温胁迫对大豆花期叶片蔗糖代谢及产量的研究较少[11],故本研究于大豆花期进行低温胁迫,通过测定叶片蔗糖代谢相关指标及产量的变化,填补该方面的空白。光合作用是作物碳代谢的物质生产和决定环节,低温胁迫主要通过损伤作物碳代谢进程影响作物产量。本试验采用自然与人工模拟环境相结合的手段,从蔗糖含量以及代谢相关酶活性两方面揭示了大豆应激花期低温胁迫的代谢机制,丰富了相关的理论研究,对指导东北地区大豆的生产实践具有重要意义。
1 材料与方法
1.1 实验设计
本研究以敏感品种(‘合丰50’)和耐冷品种(‘垦丰16’)为研究材料,两供试品种均为中熟品种、亚有限结荚习性、生育期相同。于2017年在黑龙江省农业科学院进行盆栽试验,种子播种在桶口直径28.0 cm、桶底直径20.0 cm、高23.0 cm的塑料桶内,呈五角星型种植,每穴2粒种子,大豆子叶期(VC)定苗5株,以盆为试验单元,各处理采用完全随机试验设计。
试验土壤类型为草甸黑钙土,土壤基本状况为:碱解氮含量为166.1 mg/kg,有效磷含量为38.1 mg/kg,速效钾含量为173.0 mg/kg,有机质含量为35.1 g/kg,pH 6.75。然后按N:P:K=1:2.49:1.66比例,一次性施入0.15 kg硫酸铵(含N 21%)、0.13 kg硫酸钾(含K 50%)和磷酸二铵(含P 46%~48%),充分混匀。
于花期进行低温处理1~4天,低温条件为昼夜15℃恒温[5,7],相对湿度为75±2%,光照条件正常,自然环境生长为对照(CK),即‘合丰50’和‘垦丰16’的CK表示为H-CK和K-CK,C表示低温处理,1~4表示处理天数,即‘合丰50’的低温处理表示为H-C1~H-C4、‘垦丰16’的低温处理表示为K-C1~K-C4。低温处理第4天后,将低温处理的植株搬到自然环境下进行恢复1~4天,连续进行取样用于生理生化指标的测定,处理和对照各设4次重复。
1.2 取样及测定项目
1.2.1 产量及其构成因素测定 大豆成熟期(R8期)时,各处理随机选取10株植株进行株高、底荚高度、单株荚数、单株粒数以及单株粒重(产量)等的测定。
1.2.2 碳代谢相关指标的提取及测定 于供试品种低温处理后第1天开始第1次取样,以后连续取8天。各处理选取大小均匀一致的5株植株带回室内,取大豆的功能叶片(倒三叶)速冻于液氮中,再放入-20℃冰箱中贮存,待样品全部收集完毕后,使用蒽酮比色法测定淀粉含量[21];使用间苯二酚法测定蔗糖和果糖含量[21];使用3,5-二硝基水杨酸法测定叶片内酸性转化酶(AI)、中性转化酶活性(NI)、蔗糖合成酶(SS)和蔗糖磷酸合成酶活性(SPS)[21]。
1.3 数据分析
采用Microsoft Excel 2013进行数据的录入、整理以及绘图,用SPSS 19.0做方差分析。
2 结果与分析
2.1 低温胁迫对大豆产量及其构成因素的影响
由表1可以看出,与对照(H-CK和K-CK)相比,低温处理均显著降低了大豆产量,其大小顺序依次为CK>C1>C2>C3>C4,不同品种间呈现相同的变化规律,其中,‘合丰50’的减产率依次分别为10.29%、21.70%、27.35%和30.88%,‘垦丰16’的减产率依次分别为9.58%、18.08%、22.60%和24.22%,除H-C1处理外,其他处理与对照(H-CK和K-CK)间均达到显著差异。此外,‘合丰50’和‘垦丰16’的单数荚数和单株粒数也有不同程度的减少,其中,低温胁迫4天时,‘合丰50’的单株荚数和单株粒数较H-CK分别减少了26.56%和34.45%,‘垦丰16’的单株荚数和单株粒数较K-CK分别减少了14.74%和35.40%,方差分析可知,处理与对照(H-CK和K-CK)之间差异均达到了显著差异,这说明花期遭遇低温胁迫不利于大豆单株荚数和单株粒数的形成,而大豆单数荚数或单株粒数任何一个因素减少,均会造成大豆单株产量降低。
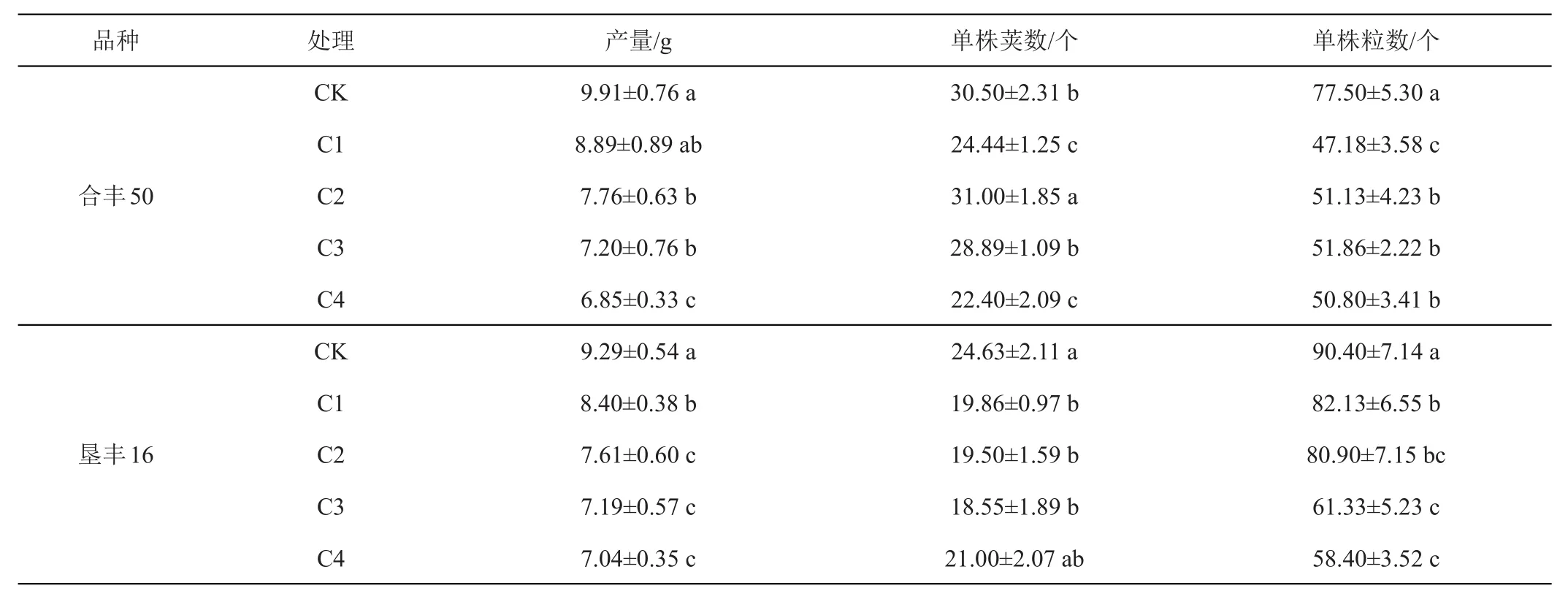
表1 低温胁迫对大豆产量及产量构成因素的影响
如表2所示,随着低温处理时间的延长,植株的株高逐渐降低,而底荚高度逐渐升高,低温胁迫对大豆基部茎粗没有显著影响。各处理株高的大小顺序依次为CK>C1>C2>C3>C4,‘合丰 50’株高的降低率分别为1.81%、2.03%、2.18%和7.05%,‘垦丰16’株高的降低率分别为3.00%、3.68%、7.98%和8.52%,方差分析可知,H-C4与H-CK、K-C3与K-CK、K-C4与K-CK之间差异极显著。就底荚高度而言,C1处理起负调节作用,C2、C3和C4处理起正调节作用,不同品种间呈现相同的变化规律,底荚高度的增加不利于植株下部产量的形成,进而降低了植株的单株籽粒重。
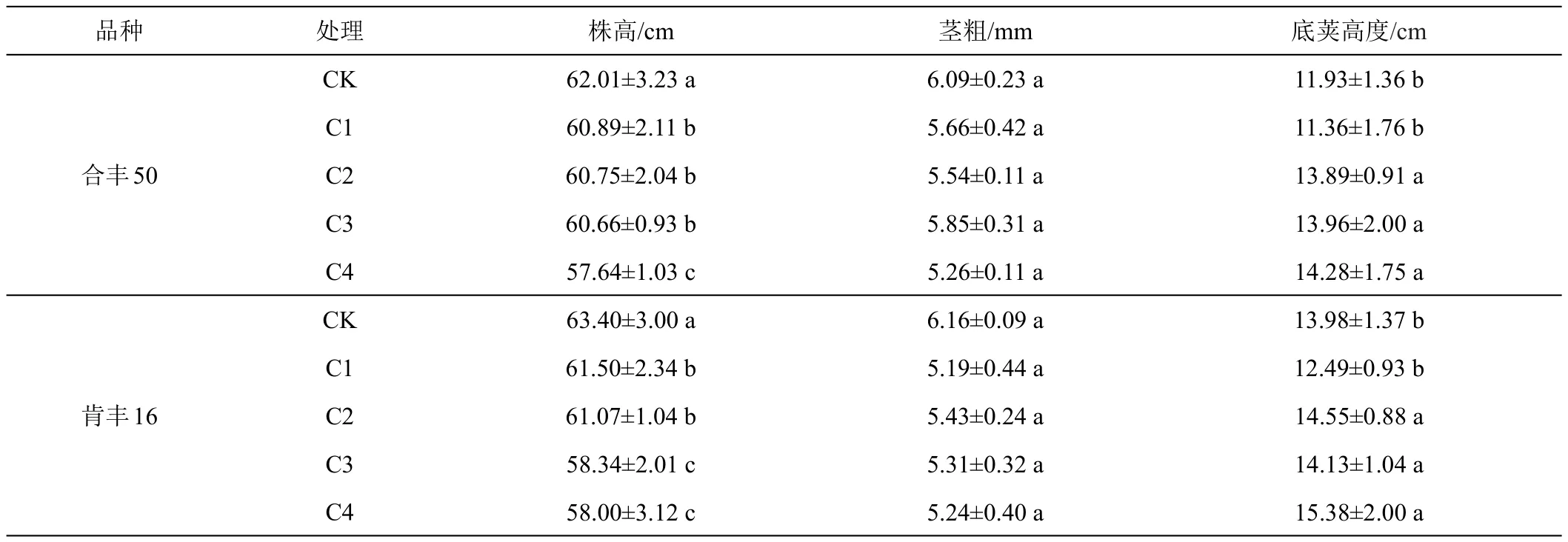
表2 低温处理对大豆植株形态指标的影响
大豆在花期遭遇低温胁迫时,不仅单株荚数和单株粒数有不同程度地减少,其粒荚数构成也发生了较大变化(图1)。‘合丰50’花期遭遇低温胁迫时,其空瘪荚、二粒荚数、三粒荚数和四粒荚数均大幅度下降,而一粒荚数略有增加;‘垦丰16’花期遭遇低温胁迫时,其空瘪荚、一粒荚数、二粒荚数、三粒荚数和四粒荚数均不同程度地减少,花期低温胁迫对冷敏感‘合丰50’荚粒数的影响较耐冷‘垦丰16’严重,推断其可能是大豆减产的主要因素之一。
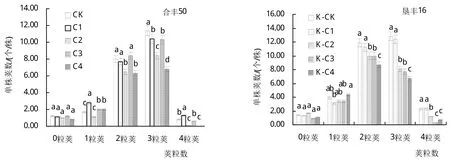
图1 低温处理对大豆单株荚粒数的影响
如图2所示,低温处理后,‘合丰50’各处理的第10节和11节产量对总产量的贡献率高于CK,其增加幅度为1.30%~28.70%;C1和C3的第5节~8节、C2和C4的第12节~14节产量对总产量的贡献率一直低于CK。低温处理后,‘垦丰16’各处理的第4节~6节产量对总产量的贡献率高于CK,其增加幅度为16.45%~69.88%;C1的第9节~12节、C2的第7节~14节、C3的第11节和第12节产量对总产量的贡献率低于CK。总之,不同的大豆品种,其减产原因不同。
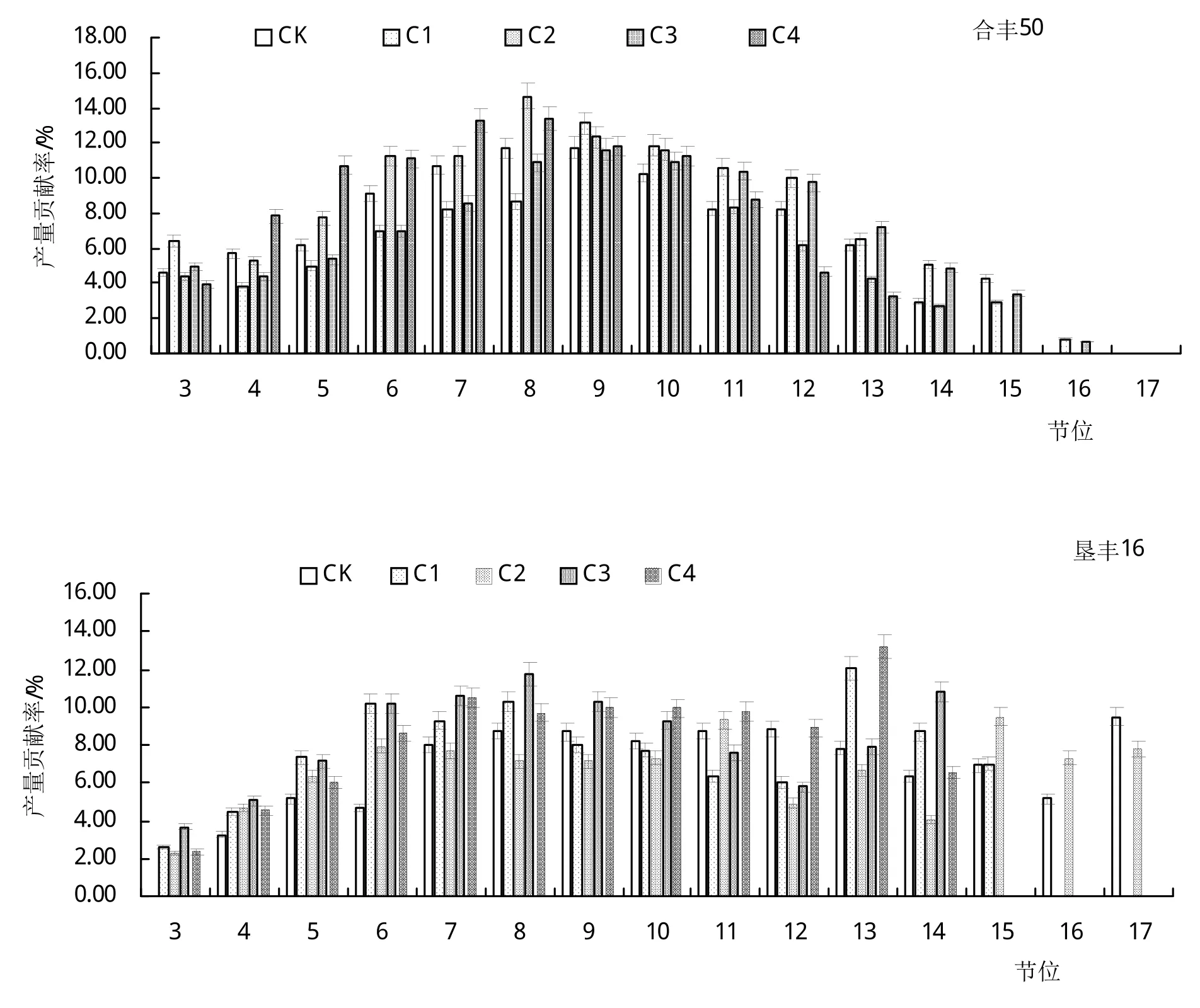
图2 低温处理对大豆不同节位产量贡献率的影响
2.2 低温胁迫对大豆叶片碳水化合物含量的影响
从图3可以看出,大豆花期进行低温胁迫可以导致叶片中淀粉和蔗糖含量减少,随着处理时间的延长,其下降幅度逐渐增加;果糖含量随着处理时间的延长逐渐提高,不同基因型品种间呈现相同的变化趋势。低温胁迫1~4天时,与对照相比(H-CK和K-CK),‘合丰50’叶片蔗糖含量分别依次减少了33.23%、41.44%、59.80%和66.74%,‘垦丰16’叶片蔗糖含量分别依次减少了23.27%、42.51%、51.98%和55.34%,可见,冷敏感品种对低温胁迫响应更敏感。
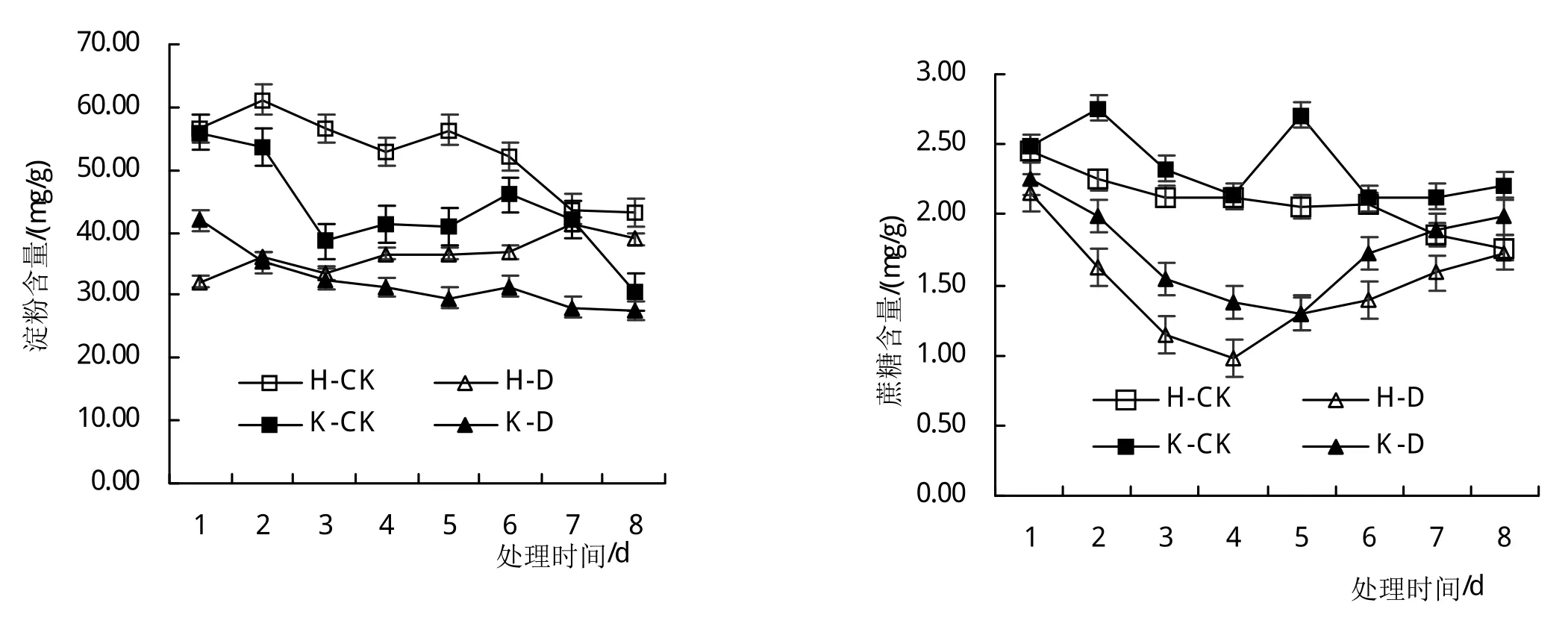
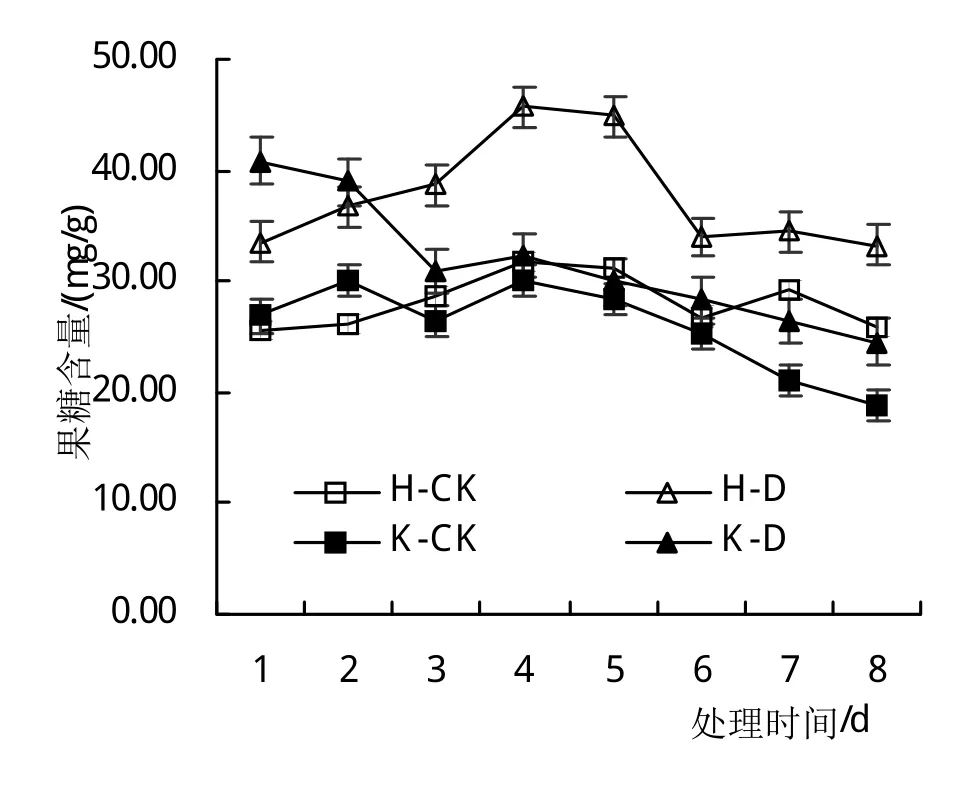
图3 低温胁迫大豆叶片碳水化合物含量的影响
随着低温胁迫后恢复时间(5~8天)的延长,叶片中淀粉和蔗糖含量逐渐增加,始终高于对照,果糖含量逐渐减少,始终低于对照,其中,叶片蔗糖含量在恢复4天后可恢复到对照水平,而淀粉和果糖含量在短时间内难以恢复到对照(H-CK和K-CK)水平,不同基因型品种间呈现相同的变化趋势。
2.3 低温胁迫对大豆叶片蔗糖代谢相关酶活性的影响
由图4分析可知,低温胁迫增加了叶片内酸性转化酶(AI)、中性转化酶(NI)和SS活性,低温胁迫第4天,与对照相比(H-CK和K-CK),‘合丰50’上述指标依次增加了59.92%、17.12%和168.18%,‘垦丰16’上述指标依次增加了45.27%、80.62%和150.23%;降低了SPS活性,随着低温胁迫时间的延长,SPS活性降低幅度逐渐增加,低温胁迫对耐冷品种叶片内蔗糖代谢相关酶活性的调控效应影响小于敏感品种。
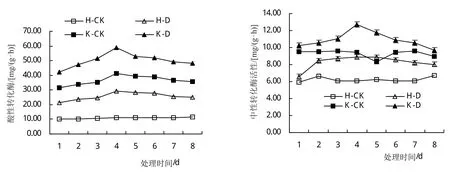
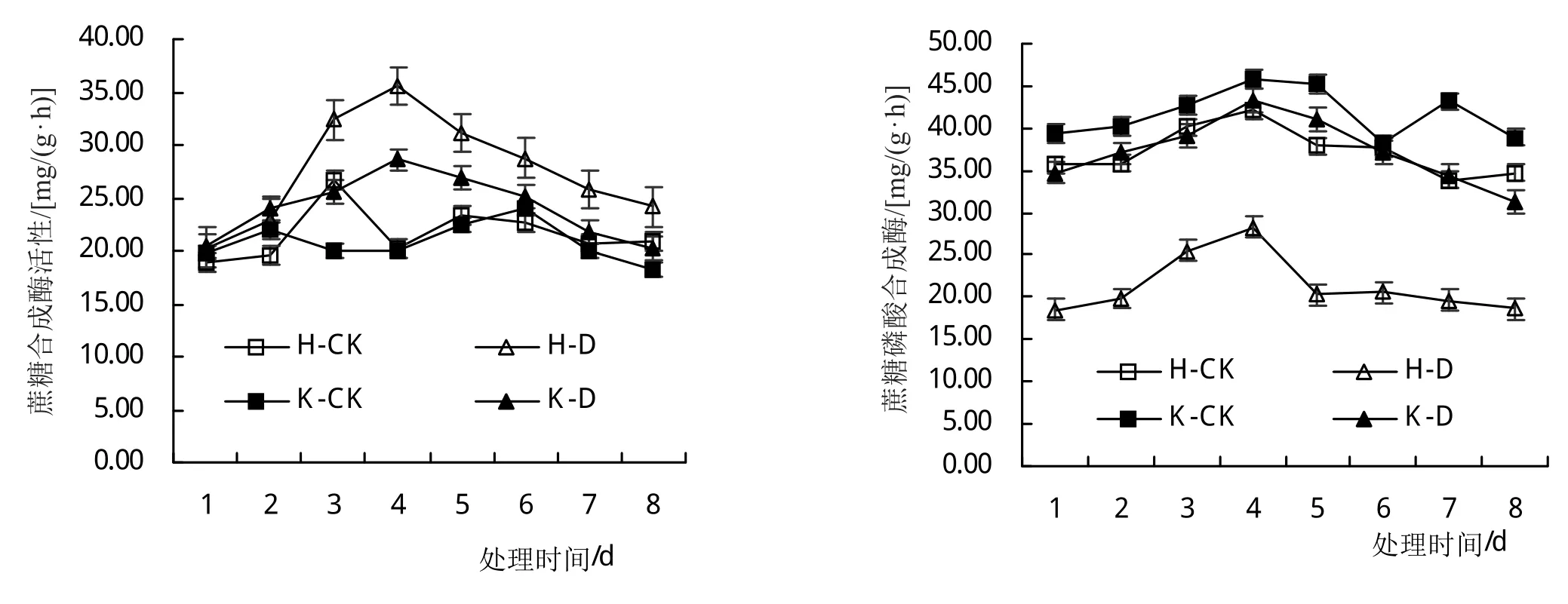
图4 低温胁迫对大豆叶片蔗糖代谢相关酶活性的影响
低温胁迫后恢复期间(5~8天),叶片中AI、NI和SS活性逐渐减少,但始终高于对照,其中,恢复第4天,与对照相比(H-CK和K-CK),‘合丰50’上述指标依次增加了16.03%、17.11%和33.03%,‘垦丰16’上述指标依次增加了17.29%、49.64%和21.94%;SPS活性逐渐增加,但始终低于对照,不同基因型品种间呈现相同的变化趋势。
3 讨论
许多学者已从不同的生理机制验证了低温胁迫导致作物减产的原因,认为花期低温胁迫造成大豆产量损失的原因是低温胁迫影响了叶片光合产物的输出,致使大豆的总状花序的花和荚不能从源器官(叶片)内获得充足的光合同化物,使植株顶端的花荚发育不良,严重时甚至大量脱落[9]。本研究也得到了相似的结果,低温胁迫降低了大豆的株高和产量,提高了其底荚高度,其中,‘合丰50’和‘垦丰16’的减产范围分别为10.29%~30.88%和9.58%~24.22%,底荚高度的增加不利于植株下部产量的形成,并且受胁迫植株的二粒荚数、三粒荚数和四粒荚数显著降低,这导致大豆的单株荚数和单株籽粒数有不同程度地减少,这与前人的研究结果相同[16-17]。
碳代谢作为植物体内有机物质合成、转化和降解的主要代谢途径,其代谢强度在作物的正常生长发育过程中的动态变化直接影响着植物体内光合产物形成、运输以及矿质营养的吸收等。短暂的干旱胁迫或者遮光可以使植株叶片内淀粉和蔗糖含量迅速下降接近为零,果糖含量升高,库器官内蔗糖合成酶活性降低而分解酶活性升高,使植物叶片内的蔗糖生物合成过程受阻[22]。Miao 等[23]和 Qi等[1]以番茄为试验材料,经低温胁迫后,发现番茄叶片内淀粉、蔗糖和果糖含量升高,转化酶和SS活性降低,SPS活性被显著提高,而胁迫恢复过程中的叶片内淀粉含量和AI活性难以恢复到对照水平[24]。木薯叶片在低温胁迫初期,其叶片内会积累较多的碳水化合物,但随着气温的逐渐降低,叶片内碳水化物的积累量会逐渐减少[24]。SPS作为催化蔗糖合成中的关键酶,在低温胁迫环境下,月季成熟叶片内SPS和AI活性呈下降的趋势,Martindale等[25]认为作物叶片内蔗糖积累的主要原因是蔗糖合成能力增强。本研究结果表明,大豆花期进行低温胁迫导致叶片中淀粉和蔗糖含量减少,果糖含量增加,提高了AI、NI和SS活性,抑制了SPS活性,其中,‘合丰50’叶片蔗糖下降幅度为33.23%~66.74%,‘垦丰16’叶片蔗糖下降幅度为23.27%~55.34%,低温胁迫后恢复期间,上述指标呈相反的变化趋势,不同基因型品种间呈现相同的变化趋势。低温胁迫造成大豆叶片内蔗糖含量减少的主要原因是蔗糖合成途径受到抑制,而分解途径加快,从源库角度来看,源器官(叶片)内蔗糖含量的减少不利于其向外运输,花作为大豆花期的主要库器官,无法从源器官得到充足的光合同化物,致使其大量脱落[13],导致最终产量降低。这与Martindale等[25]的研究结果不同,这说明植物对低温胁迫的响应受自身遗传条件、胁迫强度、胁迫时期等因素影响。
4 结论
大豆花期低温胁迫后,随低温胁迫时间的延长,叶片淀粉和蔗糖含量逐渐减小,果糖含量逐渐增加,叶片内AI、NI和SS活性提高,SPS活性被抑制;低温恢复期间上述指标呈现相反的变化趋势,这说明大豆花期遭遇低温胁迫,加快了蔗糖的分解代谢,而抑制了其合成途径,致使叶片内的蔗糖含量急剧减少,并且其危害不能在短时间(自然环境下恢复4天)内恢复,致使单株荚数或单株粒数均有不同程度的减少,最终降低了大豆的单株产量。
