氮添加对中亚热带杜鹃灌丛凋落物生产和叶分解的影响
2021-04-25李家湘樊晓亮徐文婷谢宗强
张 蔷, 李家湘, 樊晓亮, 徐文婷, 谢宗强,5,*
1 中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093 2 北京市生态环境监测中心,北京 100048 3 中南林业科技大学林学院,长沙 410004 4 河北雾灵山国家级自然保护区, 兴隆 067300 5 中国科学院大学,北京 100049
近几十年来, 由于化石燃料的燃烧和工业化肥的使用等人类活动的影响, 大气氮沉降急剧增加, 成为全球气候变化的一个重要因子[1]。在亚洲, 活性氮的释放从1961年到2000年增加了近五倍, 预计到2030年会超过105 Tg/a[2]。中国的氮沉降量近年来也快速增长, 成为继欧美之后的三大氮沉降热点地区之一, 其中亚热带地区已位于高沉降通量区域, 年氮沉降量超过30 kg/hm2[3- 4]。氮沉降的持续增加会改变自然界中元素的生物地球化学循环过程, 对生态系统产生多方面的影响, 如改变植物生产力和生理生态机制[5- 6], 导致土壤酸化、土壤元素失衡以及生物多样性丧失等一系列生态问题[7- 9]。因此, 氮沉降及其产生的生态后果已成为生态学领域研究的热点内容。
凋落物作为生态系统的重要组成部分, 在生物碳库向土壤碳库的转换过程中起到关键作用,其生产和分解过程受到大气氮沉降的深刻影响[10- 12]。一方面, 氮沉降会增加分解过程中的外源氮输入, 影响分解微生物的生长和活性, 从而直接影响凋落物分解的化学过程; 另一方面, 氮沉降改变了土壤理化性质, 影响植物生长和凋落物自身化学特性(如C/N比), 进而改变凋落物的产量和分解速率[13- 15]。近十几年来, 关于氮沉降对凋落物生产和分解的影响已开展了较多研究[16- 17]。已有的研究结果显示, 在背景氮含量较低的地区, 模拟氮沉降会增加凋落物产量; 但也有研究发现施氮对凋落物生产量的促进作用不明显, 这与林分类型、氮输入量、林龄及试验持续时间等多方因素有关[18- 19]。对于凋落物分解, 由于凋落物理化性质、分解阶段以及外界环境条件等因素的差异, 氮沉降的影响结果也不尽相同[20]。一般来说, 凋落物和环境氮贫乏, 氮沉降对分解起促进作用; 而在氮含量丰富的地区, 氮沉降则可能无实质影响或者出现抑制作用[21- 22]。在凋落物分解前期, 氮沉降通常起促进作用; 但随着分解的进行, 促进作用逐渐减弱, 且在后期常常表现为抑制作用[23]。
虽然近年来关于氮沉降对凋落物影响的研究屡现不鲜,但是,相关研究主要集中于森林和草地生态系统[24- 27], 氮沉降对灌丛生态系统凋落物生产和分解的作用规律尚不清楚。作为陆地生态系统的主要类型之一, 灌丛生态系统在全球气候调节和碳循环中发挥着不可替代的作用, 同时在群落演替过程中扮演着极其重要的角色[28]。在中国, 灌丛分布面积达74.3×106hm2, 占国土面积的7.7%, 是我国山地和干旱地区极为常见的植被类型[29]。近年来由于退化植被的恢复和重建, 我国灌丛植被的面积不断增加, 碳汇功能日益显著, 据估计灌丛碳汇约为森林碳汇的1/3。此外, 灌丛通常生长于土壤较为贫瘠的地区, 群落结构相对简单,对环境变化表现出较高的敏感性[30]。因此, 在大气氮沉降不断增加的大背景下, 有必要摸清灌丛生态系统的响应规律,以便评估其对全球气候变化产生的影响。杜鹃(Rhododendronsimsii)作为在我国南方地区广泛分布的物种, 以其为优势种的群落是亚热带最常见的灌丛类型之一[31]。因此以中国中亚热带杜鹃灌丛为研究对象, 探讨不同氮添加水平下其凋落物产生和分解的初期响应规律, 以期为揭示气候变化背景下灌丛生态系统的响应机制提供数据支撑。
1 研究地区和研究方法
1.1 研究地概况
研究地点位于湖南浏阳大围山自然保护区(28.35°—28.48°N, 114.03°—114.22°E)。大围山地处罗霄山脉北段, 以中山地貌为主, 气候属于中亚热带季风湿润气候区, 年均气温11—16 ℃, 年均降雨量1800—2000 mm, 无霜期243 d。研究地土壤垂直分带明显: 海拔800 m以下基本为红壤, 800—1200 m主要为山地黄壤, 1200 m以上为山地黄棕壤。
在植被区划上, 研究区属于中亚热带典型常绿阔叶林北部亚地带, 区内地带性植被为常绿阔叶林[31]。因长期人为干扰,大围山自然保护区森林植被曾遭到严重破坏。直到20世纪80年代,林场实行封禁管理,植被才开始逐渐恢复。如今海拔1200 m以上的区域植被逐渐演替为中山山地灌丛, 主要的建群种有杜鹃、水马桑(Weigelajaponicavar.sinic)、湖南白檀(Symplocoshunanensis)和圆锥绣球(Hydrangeapaniculata)等, 其中以杜鹃为建群种的灌丛分布最广。因此本试验以杜鹃灌丛群落作为研究对象, 进行模拟氮沉降试验。试验样地表层土壤(0—10 cm)的理化性质见表1[32]。

表1 试验样地土壤理化性质(平均值±标准误差)
1.2 试验设计
试验样地设置于大围山自然保护区海拔约1400 m的位置, 依据生境一致,地形起伏不大等原则,选择连续分布的杜鹃灌丛布设样地。试验采用随机区组设计, 整个试验样地由3个区组组成, 区组之间的缓冲距离为10 m; 每个区组布设4个5 m × 5 m的小区, 小区之间的缓冲距离为5 m。
参照试验区域的实际氮沉降水平(18.23—38.88 kg hm-2a-1)[33], 设置不同浓度梯度的氮添加试验。本研究共设置4个处理, 各处理的施氮量分别为: 对照(CK, 0 g m-2a-1)、低氮(LN, 2 g m-2a-1)、中氮(MN, 5 g m-2a-1)和高氮(HN, 10 g m-2a-1), 分别对应每个区组的4个小区,每种处理3个重复。试验为期两年, 试验时间为2012年8月到2014年8月, 施氮时间为每年3—11月,频次为每月1次。试验所用氮肥为硝酸铵(NH4NO3), 施氮时将相应量的硝酸铵溶于20 L水中进行喷洒, 对照样地喷洒等量的水。
1.3 研究方法
凋落物收集试验: 2012年8月, 在试验样地每个小区样方随机布设3个0.5 m × 0.5 m的收集器, 共需布设36个。收集器为孔径1 mm的尼龙网, 网底距地面约10 cm。收集从2012年8月到2014年8月两年间产生的凋落物,在生长季(5—11月)一般每两月一次,10月和11月为凋落高峰期,每月收集一次。将收集的凋落物带回实验室,使用烘干机在65 ℃恒温下烘至恒重, 称量得到凋落物总重量。然后将凋落物分为4类:树叶、枝条、繁殖器官(花果)和其他组分, 分别测定各组分的重量。样地凋落物产量为每块样地收集器中凋落物重量的平均值。
凋落叶分解试验: 本研究群落的优势种为杜鹃和白檀, 二者生物量占灌木层总生物量的90%以上[34- 35], 因此分别选用杜鹃凋落叶和白檀凋落叶进行分解试验。凋落叶分解试验采用网袋法, 选择10 cm × 15 cm大小的尼龙网袋,上面网眼为2 mm、下面网眼为0.3 mm,以防止袋内凋落物掉落。每袋装新鲜风干落叶5g, 在每个样方内随机地布设 30个分解袋, 四角利用铁钉固定, 以防移动。分解袋于2013年1月完成布设, 2013年4月到2013年11月间每月回收一次(6月、8月因故未回收), 次年3、5月份各回收一次。在收取时去除分解袋上附着的落叶、根系、土块等异物, 置于干净的塑料袋内及时送至实验室, 于65 ℃下烘干称重, 根据如下公式计算得到凋落叶质量残留率:
凋落叶干物质残留率=Wt/W0× 100%
式中:Wt为t时刻凋落叶干重,W0为凋落叶初始干重。
凋落叶分解速率采用Olson指数衰减模型[36]计算:
y=ae-kt
式中:y为凋落叶干物质残留率(%);k为分解系数;t为分解时间(a)。根据该模型计算凋落叶分解50%和95%所需时间。
此外,烘干的凋落物粉碎后过100目筛, 保存供化学分析(总C、总N、总P测量)。
1.4 数据分析处理
利用单因素方差分析方法(one-way ANOVA)比较不同处理样地之间凋落物产量和干物质残留率的差异。利用重复测量的方差分析(repeated measures ANOVA)比较不同氮处理下凋落物产量季节动态和凋落物分解养分动态的差异, 并分析施氮和测定时间及二者的交互作用对凋落物产量和分解的影响。各组数据进行方差分析前均进行方差齐性检验。
使用Excel 2010进行数据整理,采用R 4.0.2 和System 9.2软件进行数据的统计分析, 显著水平为0.05。相关图表的绘制使用SigmaPlot 14.0和Word 2010。
2 结果与分析
2.1 氮添加对凋落物产量的影响
对不同施氮处理的杜鹃灌丛进行连续监测, 收集从2012年8月到2014年8月产生的凋落物, 计算得到年平均凋落物产量。结果显示, 4种处理样地的年平均总凋落物产量大小顺序为LN((2541.89±112.5) kg/hm2)>MN((2342.97±519.8) kg/hm2)>HN((2087.22±391.8) kg/hm2)>CK((1936.54±358.9) kg/hm2) (图1)。LN、MN和HN处理样地的总凋落物量分别比对照样地高出31.26%、20.99%和7.78%, 表明氮添加处理增加了凋落物产量, 但施氮浓度越高凋落量增加的幅度越小, 且各处理之间的差异不显著(P>0.05)。
在杜鹃灌丛中, 凋落物各组分所占比例大小依次为凋落叶>花果>凋落枝>其他(图2)。凋落叶占总凋落量的(74.98±2.6)%—(77.20±1.1)%, 是凋落物的主要组成部分; 花果占(13.73±2.0)%—(15.23±2.5)%; 凋落枝占(6.28±2.4)%—(8.49±0.6)%; 剩余其他杂物占(1.20±0.4)%—(2.13±1.3)% (表2)。此外, LN、MN和HN处理样地的凋落物各组分的凋落量均高于对照样地, 其中凋落叶产量的大小顺序为LN>MN>HN>CK, 凋落枝和花果产量的顺序为MN>LN>HN>CK, 但各处理间的差异未达到显著水平(P>0.05)。

图1 不同氮处理下杜鹃灌丛总凋落物年产量 (平均值±标准误差)Fig.1 Annual total litter biomass of Rhododendron simsii shrubland in different nitrogen treatments (Mean±SE)CK、LN、MN和HN表示4种氮添加处理, 代表的施氮浓度分别为0、2、5和10g m-2 a-1

图2 不同氮处理下杜鹃灌丛凋落物各组分的年产量 Fig.2 Annual biomass of litter component in Rhododendron simsii shrubland under different nitrogen treatments

表2 不同氮处理下杜鹃灌丛凋落物的组成及其比例
2.2 氮添加对凋落物季节动态的影响
图3显示, 凋落物各组分表现出不同的季节变化动态。总的来看, 凋落物总量和凋落叶具有较一致的变化趋势, 即在每年10—11月份达到峰值, 在其他月份凋落量则较少, 表明叶凋落量在一定程度上主导着杜鹃灌丛群落的凋落物总量。凋落枝的产量在每年10月份达到最大值。花果凋落物的产量具有明显的季节动态, 在每年5月份达到峰值, 在其他月份的产量则很少或者没有。剩余其他杂物的凋落量变化规律不明显。不同施氮处理下, 凋落物各组分凋落量随时间的动态变化趋势基本一致(图3)。氮添加对总凋落量和叶凋落量的促进作用较为明显, 其中低氮添加的促进作用最强, 但影响没有达到显著水平(P>0.05)。
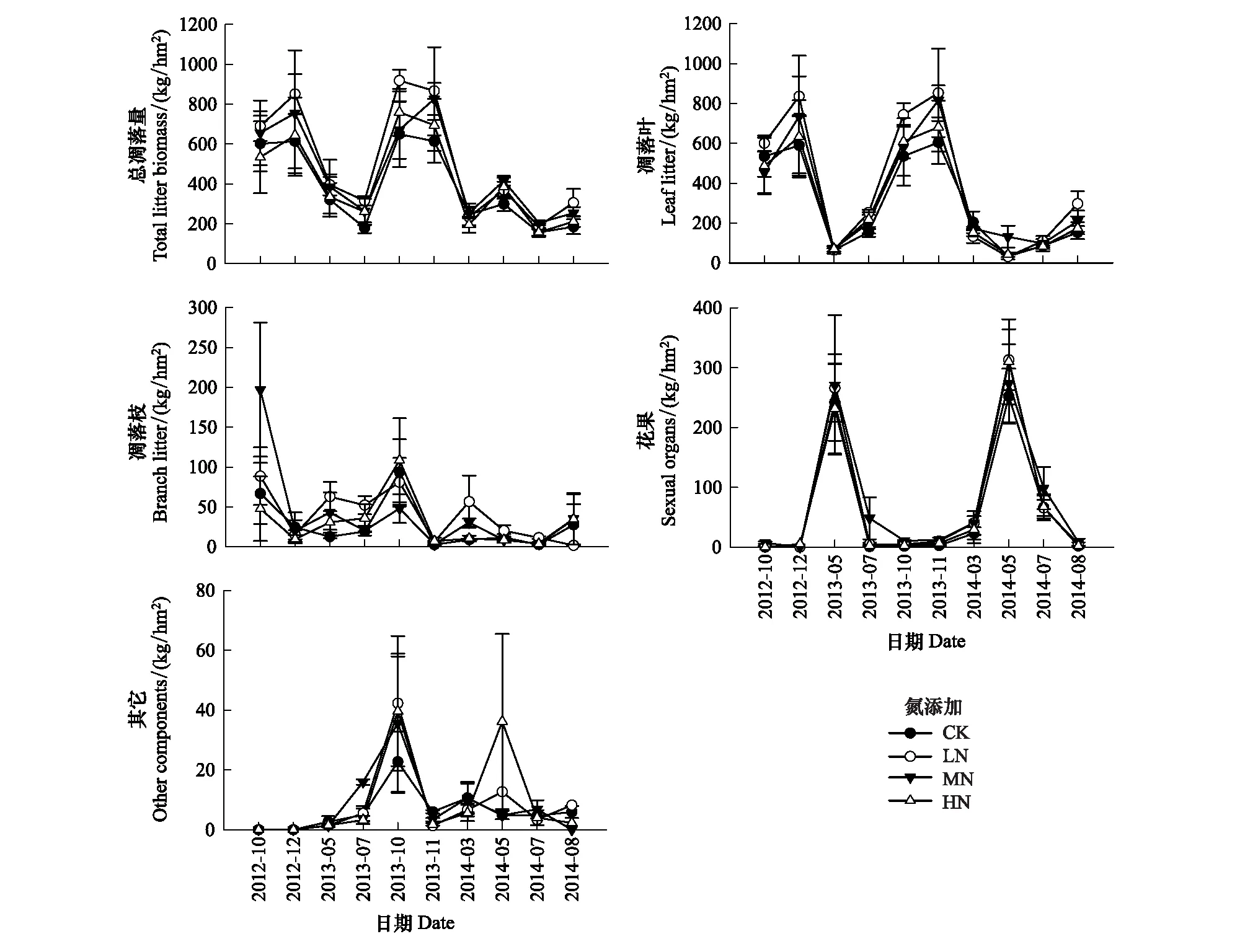
图3 氮添加对杜鹃灌丛凋落物各组分动态变化的影响Fig.3 Effects of nitrogen addition on dynamics of litter component in Rhododendron simsii shrubland
2.3 氮添加对凋落叶分解的影响
本研究分别选取群落优势种杜鹃和湖南白檀进行凋落叶分解实验。结果显示, 分解过程中凋落叶干物质残留率随时间呈下降趋势(图4)。不同物种间凋落叶分解速率差异显著(P<0.01), 其中杜鹃凋落叶分解较慢, 干物质残留率高, 经510 d分解后对照样地剩余64.4%; 白檀凋落叶分解快, 干物质残留率低, 经510天分解后对照样地剩余54.8%。在不同的分解阶段, 氮添加对两个优势种凋落叶分解速率的影响不一样(图4)。对于杜鹃凋落叶, 在分解前期MN处理对分解的促进作用较强, 在分解后期则是LN处理的促进作用较明显; 对于白檀凋落叶, 氮添加在分解前期的作用规律不明显, 在分解后期则表现出一定的促进作用。

图4 不同氮添加处理下凋落叶分解过程中干物质残留率的变化Fig.4 Change of drymass remained during decomposition process of leaf litter under different nitrogen treatments *表示不同氮处理间凋落叶干物质残留率差异显著 (P<0.05)
为进一步说明氮添加对凋落叶分解速率的影响, 利用Olson指数衰减模型对干物质残留率和分解时间进行拟合, 得到的拟合方程R2均达到显著水平(表3), 说明方程拟合效果较好。杜鹃凋落叶各处理的分解速率表现为LN>HN>CK>MN, 其凋落叶分解95%所需时间分别为7.69、11.11、12.00、17.65 a, 低氮添加促进了凋落叶分解;白檀凋落叶各处理的分解速率表现为HN>MN>LN>CK,其凋落叶分解95%所需时间分别为5.08、7.89、9.09、11.11 a, 氮添加浓度越高白檀凋落叶分解越快。此外, 两种凋落叶分解95%所需时间(5.08—17.65 a)为分解50%所需时间(1.17—4.08 a)的约4倍, 表明凋落叶分解前期快、后期慢。

表3 不同氮处理水平下凋落叶分解残留率与时间的指数方程
2.4 氮添加对凋落叶分解过程中养分动态的影响
对杜鹃和白檀凋落叶的初始化学性状进行测量, 结果表明二者具有较大差异, 白檀叶片初始N、P含量明显高于杜鹃, 且其C/N、C/P和N/P较杜鹃叶片更低(表4)。

表4 杜鹃和白檀凋落叶初始化学性状
凋落叶养分释放动态如图5所示, 杜鹃和白檀凋落叶C含量在分解过程中均表现出先下降后回升的趋势(图5), 但在整个实验阶段C含量变化比较平缓, 特别是杜鹃凋落叶C含量随分解时间变化并未表现出显著性差异(表5,P=0.87)。氮添加对杜鹃和白檀凋落叶分解过程中C元素含量的影响没有表现出明显规律, 分解300天时,杜鹃C含量表现为CK 图5 不同氮添加处理下凋落叶分解过程中C、N、P元素含量的变化Fig.5 Change of C, N and P concentrations during decomposition process of leaf litter under different nitrogen treatments 表5 不同氮添加处理下凋落叶C、N、P元素含量的重复测量方差分析 在整个试验阶段, 杜鹃和白檀凋落叶的N含量均呈现先上升后下降随后比较平稳的趋势(图5), P含量则呈逐渐上升的趋势(图5), N、P含量随时间变化均表现出显著差异(表5,P<0.05)。氮添加对杜鹃凋落叶分解过程中N、P元素的释放没有显著影响, 但是显著促进了白檀凋落叶分解过程中N、P元素的释放(表5,P<0.05)。 两年的氮添加试验结果表明, 进行施氮处理的杜鹃灌丛样地凋落物产量高于对照样地。外源氮输入可以增加土壤的有效氮水平, 从而在一定时间内促进植物生产力, 比如林业经营上经常通过施氮肥来促进树木生长[18]。以往的大量研究表明, 氮沉降可以促进陆地生态系统生物量的增长, 但由于不同生态系统类型受氮素限制的程度不同, 氮沉降对生产力的促进程度也有所差异[37- 38]。与森林生态系统相比, 灌丛对环境的变化更为敏感, 也更易受外界环境影响[30]。本研究结果显示施氮提高了杜鹃灌丛群落的生产力, 反映出大气氮沉降也许会在一定程度上增加植被的碳汇功能。但是施氮浓度越高, 杜鹃灌丛凋落物生物量增加的幅度越小, 这一结果与前人的研究相一致[39- 40]。造成这一现象的原因可能有两个方面, 其一是本研究区域土壤氮含量丰富(表1), 且本底氮沉降处于较高水平(18.23—38.88 kg hm-2a-1)[33], 当施氮浓度较高超过植物生长需求时, 会引起生态系统氮饱和现象, 造成土壤酸化、光合作用氮利用率降低,继而降低植被生产力[41]。其二是氮添加提高了土壤的氮素有效性, 这会引起土壤养分平衡的破坏, 使磷元素等有效性较低的养分成为植被生长的限制性因素, 从而造成植物生长受抑制、生产力下降[42]。张弛等[39]对亚热带常绿阔叶林的研究表明, 在高浓度氮添加的同时施加磷肥可以明显提高森林生产力。此外, 灌丛对外源氮输入的响应需要一定时间, 从氮输入到被根系吸收, 再到应用于光合作用, 这是一个复杂的机制, 短期内可能难以表现出显著的效应[43]。 在不同氮添加水平下, 杜鹃灌丛的总凋落量和叶凋落量均呈现出明显的单峰型季节变化动态。这与亚热带其他地区常绿阔叶林凋落量的季节动态模式多呈双峰型不同, 主要是由于本研究群落落叶树种较多, 其生物量占到灌木层总生物量的95%以上, 在每年秋季10—11月, 由于气温下降, 叶片衰老而大量脱落, 从而出现凋落峰值[35,44]。总凋落物和凋落叶的季节动态曲线相似, 是因为落叶占了总凋落量的75.75% (表2), 在一定程度上主导着杜鹃灌丛的凋落物总量。此外, 凋落枝的峰值也出现在每年秋季, 其动态除与季节性凋落有关外, 受气象条件特别是降雨、强风干扰以及冬季积雪的影响也较大[45]。花果的凋落量在每年5月达到峰值, 原因是本群落优势树种杜鹃的花期在4—5月份, 在5月中下旬会产生大量的繁殖器官凋落物。 凋落叶干物质残留率随时间变化整体呈下降趋势, 其中白檀凋落叶分解速率较杜鹃更快。凋落物的分解主要受到外界环境及其自身理化性质的影响[20], 本试验中两种凋落叶的分解环境相似, 不同区组间凋落叶分解速率差异不显著(P>0.05), 两个物种分解速率不同主要是由于凋落叶自身性质的差异。白檀凋落叶分解速率更快的原因可能有两个方面, 一是白檀叶片的初始N、P含量较高, C/N比值较低(表4), 丰富的养分有利于微生物的生长, 从而促进凋落叶分解; 二是白檀叶片一般为膜质或薄纸质, 与革质的杜鹃叶片相比更易于分解。此外, 杜鹃和白檀凋落叶均表现为前期分解速率较快, 这与凋落物中易分解的碳水化合物的快速降解、水溶性物质的淋溶有关; 而随着凋落物的进一步分解, 木质素、纤维素等难分解的化学成分逐渐积累, 分解速率趋于缓慢[46]。 研究结果显示,氮添加对白檀凋落叶分解表现出一定的促进作用, 与对照样地相比, 施氮使凋落叶分解周期缩短了18.18%—54.28%。氮添加可以通过多个途径对凋落物分解产生影响, 如改变凋落物的化学元素含量, 改变植被组成和植物生长状况, 改变土壤微生物群落等[14, 18]。外源氮输入增加了微生物可利用氮, 促进其生长繁殖,有利于凋落物分解; 另外, 施氮还会促使微生物群落组成向更加高效但对氮有更高需求的方向转变, 这也许是施氮促进凋落物分解的另一个原因[20]。杜鹃凋落叶分解对氮添加的响应与白檀有所不同,Olson指数模型拟合结果显示, 低浓度氮添加缩短了杜鹃凋落叶分解周期, 但当施氮浓度升高时则会对凋落叶分解产生抑制作用。这可能是由于氮素不是本区域杜鹃凋落叶分解的主要限制因子, 杜鹃叶片N/P=17.42 (表4), 一般来说N/P<14时受N限制, N/P>16时受P限制[47]。李仁洪等[48]对慈竹凋落物分解的模拟氮沉降研究也显示, 氮沉降会对竹子叶和枝凋落物的分解产生促进作用, 但是当施氮水平升高时, 促进作用会逐渐减弱; 陈翔[49]的研究也发现, 适量的氮添加可以促进落叶松凋落物分解, 但随着氮素水平的上升, 这种促进能力会减缓甚至产生抑制作用。本研究中白檀和杜鹃凋落叶分解对氮添加表现出不同的响应规律,事实上,已有的研究显示氮沉降是否改变凋落物分解速率会受到试验树种、氮处理类型及试验时长等多种因素的影响[50]。例如项文化等[51]对亚热带杉木林的研究表明施氮会显著提高凋落物分解速率; 而宋学贵等[52]对川西南常绿阔叶林的研究显示氮添加对凋落物分解表现出显著的抑制作用; 但据莫江明等[53]统计, 约有一半的研究表明氮处理并没有在统计学上显著改变凋落物分解速率。 在凋落叶分解过程中, 不同氮添加水平下杜鹃和白檀凋落叶N含量均呈先上升后下降的趋势, 即表现为富集—释放模式, 这与在森林和草原进行的施氮试验结果一致[27, 46]。凋落物元素残留率随时间出现波动变化, 表明在分解过程中环境对分解物的养分元素存在补充和再释放的过程, 这可能与分解过程中微生物的活动节律有关[54]。有研究显示, 在分解初期微生物会进行固氮来维持自身的生长, 这些微生物附着在分解物上会导致氮富集; 而后随着微生物大量繁殖, 氮消耗加快, 又会造成氮释放[55]。另外, Roskoski[56]在阔叶林中进行的研究发现, 凋落物分解过程中可能存在自生固氮菌的侵入, 这也许是分解前期产生N富集的原因之一。与N元素的波动变化趋势不同, 本研究中叶片P元素在分解过程中主要表现为富集模式, 这与仲米财等[55]在中亚热带杉木林的研究结果有所不同。事实上, 在不同的研究中凋落物释放P的临界值存在差异, Cromack等[57]将C/P为240作为凋落物固持P的临界值, 而Lousier等[58]发现大叶钻天杨(Populusbalsamifera) 和欧洲山杨(Populustremula)凋落物释放P的临界值为230。本研究中杜鹃和白檀凋落叶的初始C/P分别达到492和283(表4), 均大于上述两个临界值, 因此在分解过程中P含量呈增加趋势。而且由于杜鹃凋落叶C/P远高于白檀, 其叶片在分解过程中的P富集现象也更明显。 氮添加对凋落叶分解过程中C、N和P元素的释放分别具有不同的作用结果。其中,施氮对C元素的释放没有显著影响, 这一结果与在亚热带杉木人工林进行的模拟氮沉降研究结果相似[59]。Magill等[60]在美国东北部Harvard林开展的氮添加试验也发现, 施氮使凋落物中木质素的含量显著增加, 但对纤维素含量无显著影响。但是, 氮添加显著促进了白檀凋落叶分解过程中N、P养分的释放, 而对杜鹃凋落叶N、P的释放没有显著影响。本试验中杜鹃和白檀凋落叶的分解环境相似, 但是二者的化学性状差异较大, 白檀凋落叶的N/P比值低于杜鹃, 施氮会改善其周围氮缺乏的状况, 从而促进微生物群落的生长繁殖, 加快其N、P元素释放。在美国夏威夷雨林开展的模拟氮沉降试验显示, 氮输入会使植物体产生大量的胞外磷酸酶, 导致有机物分解出更多的磷酸盐[61]; 在热带雨林开展的施氮试验也发现, 施氮肥会加速凋落物分解过程中的磷释放[62]。但是Elser等[63]在全球范围的研究发现, 当生态系统达到氮饱和后, 持续进行氮输入可能会抑制磷元素的周转。所以, 氮添加对凋落物养分释放的影响既取决于分解者的种类和活性, 也受到气候、树种等诸多因素的影响[59, 64]。

3 讨论
3.1 氮添加对杜鹃灌丛凋落物产量的影响
3.2 杜鹃和白檀凋落叶分解及其对氮添加的响应
3.3 凋落叶分解过程中的养分动态及其对氮添加的响应
