武夷山不同林龄甜槠林土壤呼吸特征及影响因素
2021-04-25陈燕菲吴承祯范海兰林勇明
赵 青,刘 爽,陈燕菲,吴承祯,范海兰,林勇明,李 键,*
1 福建农林大学林学院,福州 350002 2 福建省高校森林生态系统过程与经营重点实验室,福州 350002 3 福建省生态产业绿色技术重点实验室,南平 354300 4 武夷学院生态与资源工程学院,南平 354300
土壤作为陆地生态系统中最大的碳库,包含2000 Pg土壤无机碳及1500 Pg土壤有机碳[1]。全球尺度上,每年土壤通过呼吸作用释放出的CO2约为化石燃料燃烧释放CO2的10倍以上[2],土壤呼吸(Soil respiration,RS)产生的微小变化都可能对大气中CO2浓度、全球碳平衡产生极大的影响[3]。森林作为陆地生态系统的主体,其RS占总呼吸的69%,森林RS变化直接影响着全球碳循环的动态变化[4],已成为陆地生态系统碳循环过程和全球气候变化研究的热点问题之一。
RS受气温、降水、土壤质地、植被类型、人类活动等多种因素的综合影响[5- 8],其中林龄因素作为影响林下植被生物量、凋落物量的重要因素,对气温、土壤温度、生物量等非生物、生物因素的响应均存在差异性。随着林龄的增大,森林物种组成、群落结构、动植物的丰富度都会发生变化[9- 10],不同林龄植被群落生产力、年凋落量、生物量积累不同,进而影响森林生态系统的碳排放和碳循环过程[11]。Ewel等关于热带森林RS研究表明,随着林龄增大,RS随之增大[12];Saiz 在对不同林龄北美云杉RS研究表明,RS随林龄增大,呈先减小后保持稳定的趋势[13];王娟等对不同林龄锐齿栎林RS研究表明,RS变化与林龄的关系并不显著[14]。由此可见,林龄对RS动态变化影响的研究结果仍存在较大的不确定性。
武夷山自然保护区是我国中亚热带常绿阔叶林保存较为完好的代表性区域[15],甜槠林作为武夷山自然保护区常绿阔叶林群落的优势种之一[16],在中亚热带森林生态系统碳平衡中占有重要的地位[17]。因此,探究甜槠天然林RS随林龄变化特征及其影响机制,对于准确评估本区域森林生态系统碳收支及碳循环具有重要的意义。目前关于该区域甜槠林研究主要集中在甜槠林群落结构特征[18]、生物量[19]、凋落物[20]、养分循环[21]等方面,其RS研究主要在探究温度、水分等非生物因素对RS日动态的影响,土壤温度在非生物因素中对RS日变化的影响最大[22]。生物因子特别是植物根系通过“生物泵”的形式向地下生态系统转移C,是土壤C输入的主要途径[23],然而在不同年龄序列上植物根系对RS的调节机制研究相对较少,而随着土壤水分、温度的改变势必会影响植物根系分布、根系呼吸的组成,进而对土壤呼吸产生影响。以武夷山自然保护区4个年龄阶段天然甜槠林(18、36、54、72 a)为研究对象,旨在探究(1)甜槠林RS及其组分随季节变化趋势;(2)探究影响RS的主要因素,特别是生物因素随林龄的变化趋势及其与RS间的响应。
1 材料与方法
1.1 研究区概况及样地设置
研究地点位于武夷山自然保护区内(27°33′N,117°27′E)。该地区位于武夷山脉北部,平均海拔1200 m,主峰为黄岗山,海拔2158 m。气候属典型的亚热带季风气候,年均气温18.1℃,年降水量在2000 mm左右,年相对湿度为82%—85%,无霜期为252—272 d[24]。区内现存世界同纬度面积最大、保存最完整的中亚热带森林生态系统,地带性植被为亚热带常绿阔叶林,分布在海拔400—1100 m处,且绝大部分为天然林和天然更新次生林,壳斗科为构成森林群落的主要建群种、优势种。区内土壤垂直分布规律显著:海拔700 m以下为红壤、700—1100 m为黄红壤、1100—1900 m为黄壤、1900 m以上为山地草甸[18- 19]。
实验区样地位于武夷山自然保护区生态定位站以东的甜槠林群落内,海拔高度为700 m,群落内甜槠林生物量中各层次所占比例大小依次为:乔木层>凋落层>灌木层>草本层,群落的shannon-Wiener多样性指数为3.814,Simpson指数为0.908,盖度在90%以上。为评估不同林龄甜槠林对土壤呼吸的影响,本研究采用空间代时间法[25],以甜槠天然林4个林龄等级(18、36、54、72 a)为研究对象,甜槠林林分年龄由森林管理经营记录结合树木每木检尺获得,同时测定各林龄甜槠林的叶面积指数、树高、胸径、相对密度等指标(表1)。为尽量避免气候、土壤类型的差异,确保RS差异原因为林龄的差别,研究设定每个林分之间的距离小于10 km。
林下植被概况:72 a甜槠林胸径最大,冠层郁闭度最高,林分密度为1189株/hm2,乔木层组成种类主要为青冈栎(Cyclobalanopsisglauca)、苦槠(Castanopsissclerophylla)、木荷(Schimasuperba)、石栎(Lithocarpusglabra)、南岭栲(Castanopsisfordii),同时混生光叶石楠(Photiniaglabra)、山杜英(Elaeocarpussylvestris)、山矾(Symplocossumuntia)等树种。灌木层主要树种有冬青(Ilexchinensis)、细枝柃(Euryaloquaiana)等,草本层较稀疏主要由淡竹叶(Lophatherumgracile)、芒萁(Dicranoprerisdichotoma)等构成;相较于7 2a甜槠林,54 a甜槠林胸径、林分冠层郁闭度明显变小,林分密度为1231株/hm2,冠层覆盖度为56%,乔木层组成种类主要为木荷(Schimasuperba)、马尾松(Massonpine)、山杜英(Elaeocarpussylvestris)、杉木(Cunninghamialanceolata)、冬青(Ilexchinensis)等,灌木层主要为肿节少穗竹(Oligostachyumoedogonatum)、马银花(Rhododendronovatum)、宜兴苦槠(Pleioblastusyixingensis);草本层群落物种主要由里白(Diplopterygiumglaucum)、鸡血藤(SpatholobussuberectusDunn)等草本植物构成;36 a甜槠林林分密度为为1279株/hm2,冠层覆盖度为48%,林下有较多的灌木、草本类植物,多为小叶赤楠(Syzygiumgrijsii)、桃叶石楠(Photiniaprunifolia)、赤楠(Syzygiumbuifolium)、香槟杜鹃(R.champinicane)等;18 a甜槠林胸径最小,林分密度为为1478株/hm2,冠层覆盖度为39%,伴生树种多为木荷(Schimasuperba)、马银花(Rhododendronovatum)、芒萁(Dicranoprerisdichotoma)、狗脊(Woodwardiajaponica)、鹿角杜鹃(Rhododendronlatoucheae)等树种,高大乔木少。
在4种不同林龄(18、36、54、72 a)甜槠天然林内各选择生长发育好、受人为影响小、且立地条件相似的3块20m×20m样方作为固定样地,共计12块样地,进行土壤呼吸及环境因素测定。
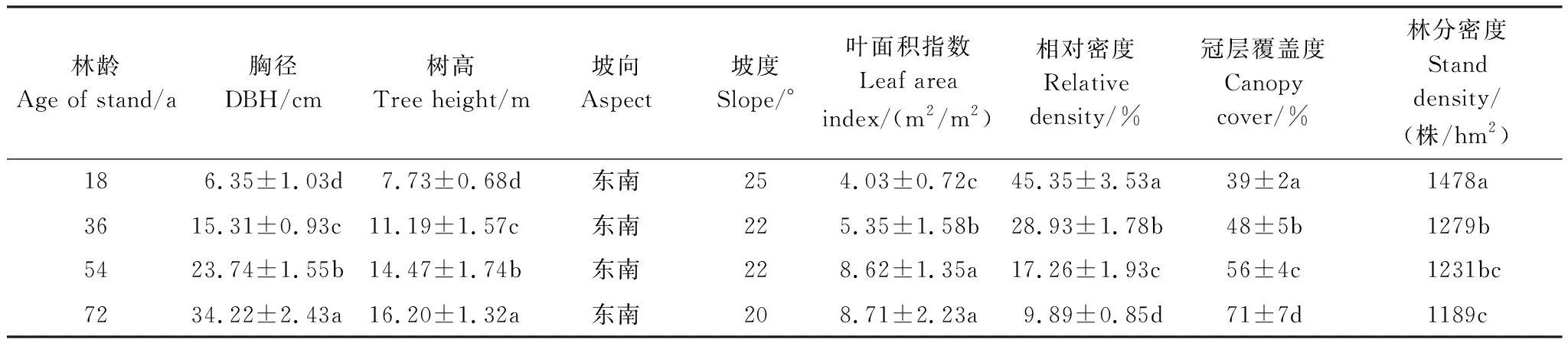
表1 研究区样地概况
1.2 土壤呼吸及相关环境因子测定
2017年3月在每个样地中心位置为起点呈Z字形随机设置5个高10 cm,直径为20 cm的PVC土壤呼吸圈,将PVC土壤呼吸圈一头削尖,嵌入地表深度为5 cm,且保证实验阶段土壤圈测定位置不变。在2017年3月—2018年2月每月下旬选取3d采用土壤碳通量系统(LI-8100,LI-COR)进行24 h连续测定(时间一般从当日9:00—11:00间开始到次日满24 h),每隔2 h测定一次,每次测定3 min,重复测定3次,测定土壤月、季节(2017年4月,春季;2017年7月,夏季;2017年10月,秋季;2018年1月,冬季)呼吸速率。为避免土壤、根系、凋落物对测量结果的影响,在每月测定前1d将土壤圈内植被、凋落物及其他杂物取去除,用于观测土壤总呼吸速率(RS)。同时选用壕沟法,将土壤进行挖沟断根处理用于测定土壤各组分呼吸速率。挖沟法具体为:在每个样方附近预先设置1 m×1 m样方作为土壤异养呼吸观测点,在土壤呼吸速率测定前5个月,即2016年10月在样方四周挖掘深度1 m,宽度60 cm环形壕沟,将1 m×1 m石棉网放入所挖壕沟,用4块1 m×1 m石棉网将样方围住,防止植被根系向外伸长,5个月后植物根系断死测定土壤异养呼吸值就会比较精确。土壤总呼吸(RS)与异养呼吸(RH)之差为土壤自养呼吸(RA)。土壤温度、含水量采用LI-8100自带温度、含水量探头(ML2x)进行同步测定,测定深度均为5 cm。
1.3 凋落物及根系生物量侧定
在各调查样地内随机设置5个1 m×1 m的凋落物收集框,收集框为孔径为1 mm的尼龙网制成的方口盘,网底距地面0.5 m,于研究时段每月下旬收集1次凋落物。将收集的凋落物按叶、枝、繁殖器官、果称鲜重后带回实验室在85℃恒温箱中烘干至恒重,由干/鲜重比值计算出含水率,最后换算出各月单位面积内凋落物产量(图1)。
根系生物量测定采用土钻法及全根挖掘法进行取样[26],每月下旬与土壤呼吸测定同步。在每个土壤呼吸圈附近用5 cm直径的根钻随机钻取土芯,共钻取10个,采集0—20 cm、20—40 cm、40—60 cm三层垂直土层土芯,将采集的土芯分层混合装袋,带回实验室用筛孔为20目筛网在水中反复清洗,用游标卡尺测量捡出细根(根径≤2 mm),粗根(根径>2 mm),风干后称鲜重,然后烘干至恒重。
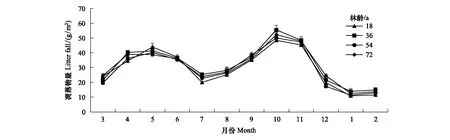
图1 不同林龄甜槠天然林凋落物年动态Fig.1 Annual dynamics of monthly litter in Castanopsis eyrei forests with different forest ages
1.4 数据处理
实验期间相邻3 d所测数值相差不大,因此将测定前2 d的土壤呼吸速率、5 cm深土壤温度、土壤含水量数据平均成1 d,进行土壤呼吸速率与土壤温湿度模型拟合,第3天数据作为实测样本对所建模型进行验证。利用SPSS 19.0软件进行数据分析,Excel 2010进行数据的统计、前期计算以及作图。
1.5 模型构建
土壤呼吸与土壤温度、土壤含水量之间单因素模型选用常见的经验模型:分别采用指数、线性、对数、幂指数、二次曲线模型进行拟合,用线性模型、指数模型、非线性模型来拟合土壤温度、土壤含水量对土壤呼吸的共同影响,线性模型进行土壤各层细根生物量与土壤呼吸速率拟合。
Rs=aebXQ10=e10b
(1)
Rs=aX+bRs=aln(X)+bRs=aXbRs=aX+bX2+c
(2)
Rs=aebTWcRs=a+bT+cWRs=a(T×W)+b
(3)
Rs=a+bX2+cT
(4)
式中,RS为土壤呼吸速率(μmol m-2s-1),X分别可带入5 cm深土壤温度(℃)、土壤含水量(%)进行计算,T为5 cm深土壤温度(℃),W为土壤含水量(%);X2为0—60 cm土层细根生物量(t/hm2);a,b,c为拟合参数;Q10值为土壤呼吸的温度敏感性指数,指土壤温度每升高10℃时土壤呼吸速率的变化值。
均方根误差(RMSE):表示模型的预测精度(实际值和模型预测值之差),值越小,模型精度越高。
(5)
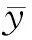
2 结果与分析
2.1 不同林龄甜槠林土壤呼吸特征及影响因素研究
4种林龄甜槠林RS季节变化呈明显的单峰趋势(图2),表现为在夏季(6、7月)最高,冬季最低(12、1月),如36 a甜槠林在6月份达到一年中达到RS最大值7.18 μmol m-2s-1,72 a甜槠林在7月份达到一年中RS最大值7 μmol m-2s-1。18、36、54、72 a甜槠林RS季节变化范围(及年平均值)为1.54—6.35 μmol m-2s-1(3.54 μmol m-2s-1)、2.33—7.18 μmol m-2s-1(4.32 μmol m-2s-1)、2.21—6.89 μmol m-2s-1(3.95 μmol m-2s-1)、1.98—7 μmol m-2s-1(4.01 μmol m-2s-1),RS季节变化与林龄大小无关。由表2可知,不同林龄甜槠林异养呼吸速率占总呼吸的比例比较大,秋季土壤异养呼吸占总呼吸的比例在四个季节最高。
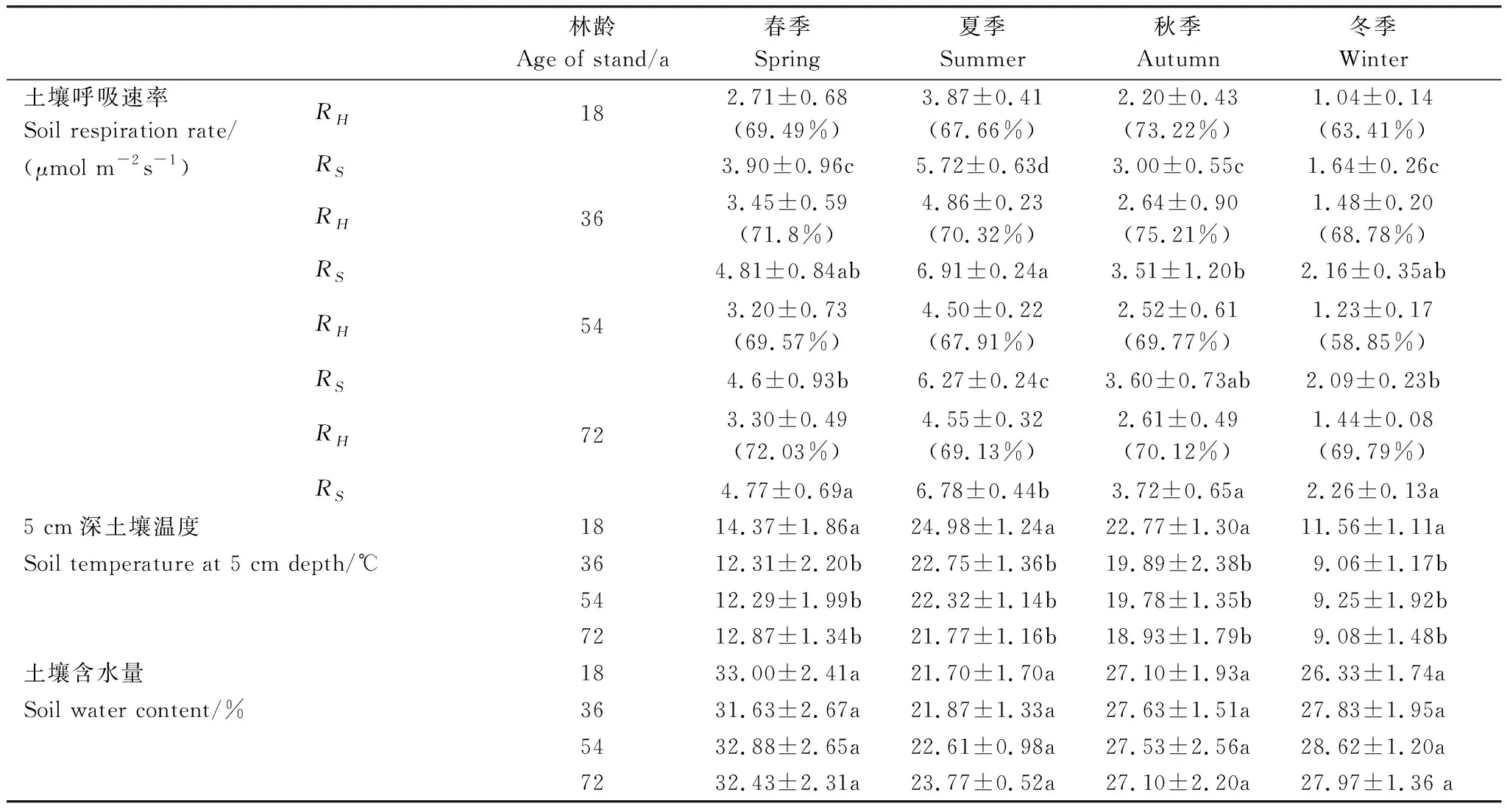
表2 不同林龄甜槠林土壤呼吸速率、5 cm深土壤温度、土壤含水量季节变化
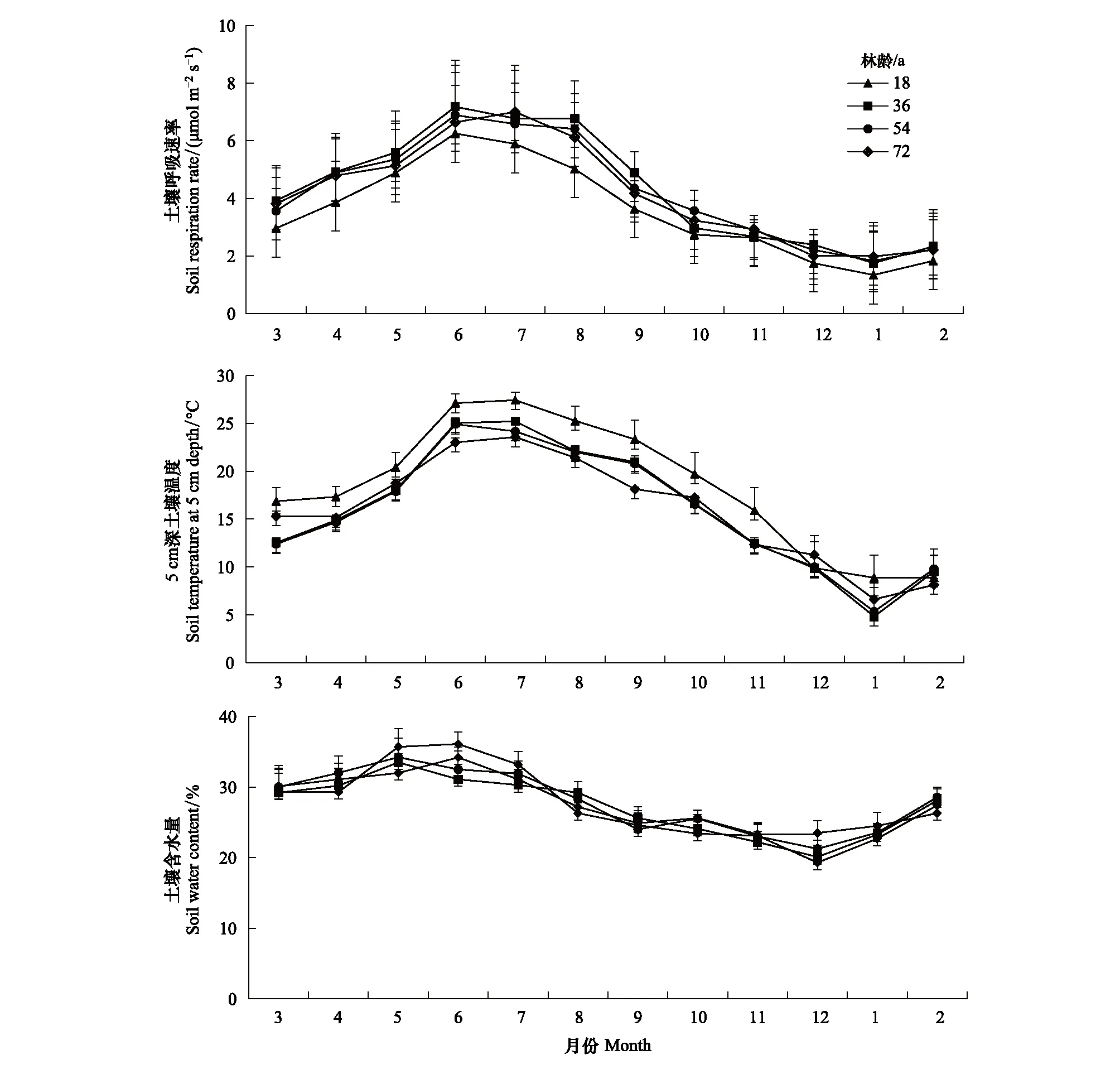
图2 不同林龄甜槠林总土壤呼吸速率、5 cm深土壤温度、土壤含水量月变化动态Fig.2 Monthly changes of total soil respiration rate, soil temperature at 5 cm depth and soil water content in Castanopsis eyrei forests with different forest ages
不同林龄甜槠林土壤温度变化与RS变化趋势一致,土壤含水量在6月达到最高值,12月为最低值。方差分析表明(表2),林龄对冬季RS影响不显著(P>0.05),秋季18 a甜槠林土壤呼吸速率值与其余3种林龄差异显著(P<0.05),4种林龄甜槠林土壤春季含水量高于秋季,很可能是因为武夷山自然保护区雨季集中在春夏季,秋冬季相对较少。18 a甜槠林土壤温度与其余3种林龄差异显著(P<0.05),林龄对土壤含水率并没有显著影响(P>0.05)。
2.2 模型建立
2.2.1土壤温度与土壤呼吸单因子模型
为探究温度、水分与土壤呼吸速率之间的关系,采用土壤温度与RS,土壤含水量与RS进行指数、线性、对数、幂指数、二次曲线模型拟合(表3)。
结果表明,18 a甜槠林所有拟合方程的R2均达到0.80以上,其中二次曲线模型拟合(0.826)结果最好,在拟合方程的选取上,二次函数方程更为复杂,拟合R2仅比幂指数大0.005,差距不明显,幂指数模型为18 a甜槠RS预测的最佳模型;36 a甜槠林所有模型拟合R2均大于0.80,对数模型拟合程度(0.840)最好,为36 a甜槠RS预测的最佳模型;54 a甜槠林指数模型拟合R2(0.852)最好,成为54 a甜槠RS预测的最佳模型;72 a甜槠林二次曲线模型拟合R2(0.862)最好,但拟合R2仅比线性模型拟合R2高0.002,为了方便计算,线性模型成为72 a甜槠RS预测的最佳模型。
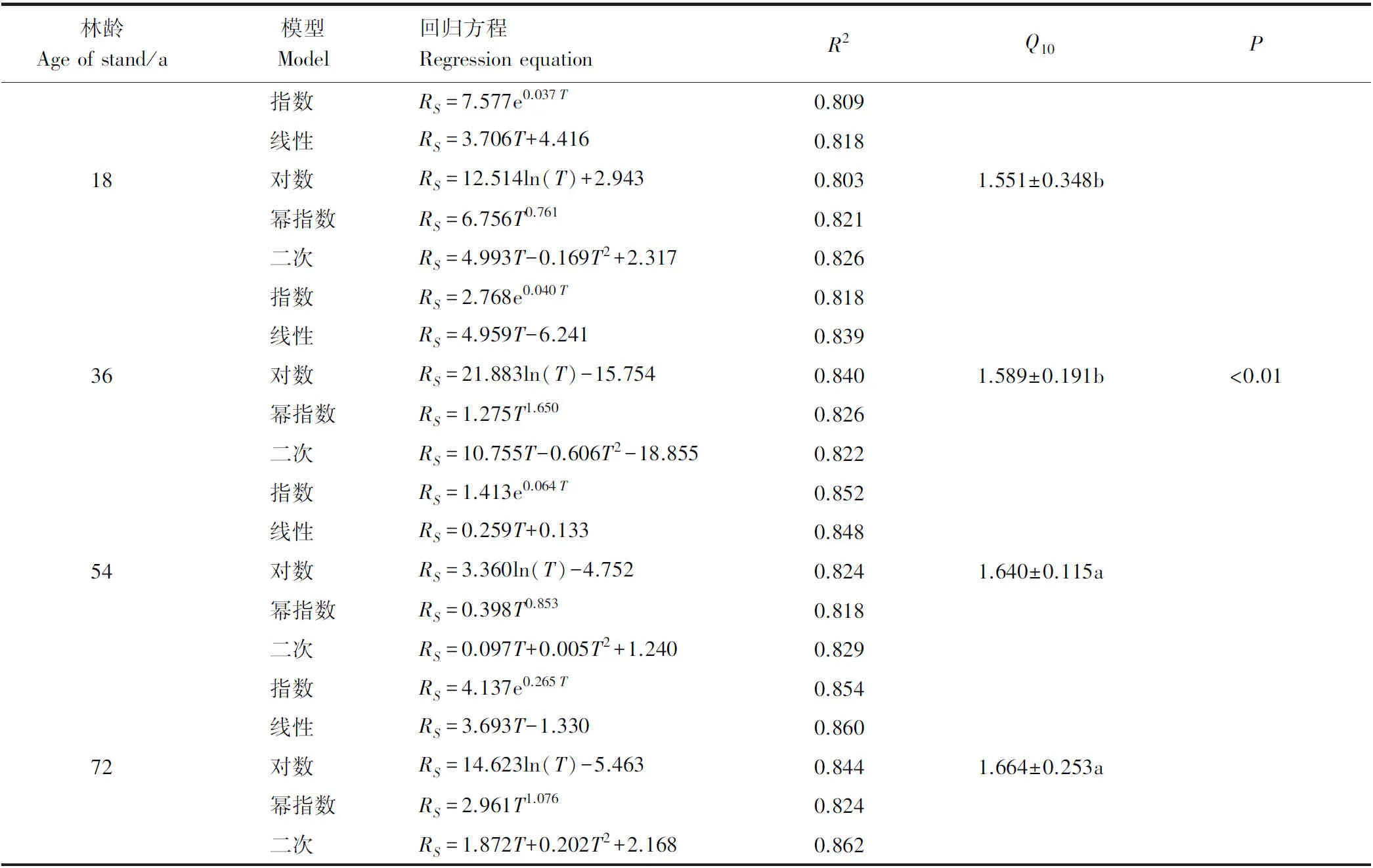
表3 5 cm深土壤温度与土壤呼吸速率拟合方程
2.2.2土壤含水量与土壤呼吸单因子模型
表4为土壤含水量与RS模型拟合方程。结果表明,甜槠林四种林龄土壤含水量与RS模型拟合结果小于5 cm深土壤温度与RS之间模型拟合R2。18 、36、54、72 a甜槠林土壤含水量与RS拟合的最佳模型分别为指数、幂指数模型(0.532)、指数模型(0.481)、指数模型(0.518)、线性模型(0.523)。
根据拟合结果,虽然温度是影响甜槠各林分RS的主要原因,但湿度因素作用仍不可忽略,RS动态变化是土壤温度、土壤水分共同作用的结果,因此需将二者共同引入方程进行模拟,提高RS的精确性。
2.2.3土壤温度、土壤体积含水量与土壤呼吸速率多因素模型及预测
表5为土壤温度、土壤含水量与RS双因素模型拟合结果。从拟合效果来看,18 a甜槠林中R2模型拟合R2(0.877)最高,36 a甜槠林中R1模型拟合R2(0.871)最高,与土壤温度、RS最佳单因子拟合模型相比,R2略有提高,但差距不明显;54 a甜槠林中R2模型拟合R2(0.901)最高,72 a甜槠林中R2模型拟合R2(0.903)最高。3种拟合模型R2较土壤含水量与RS拟合方程提高明显,可见,湿度因素对甜槠林RS作用不明显,温度因子是影响甜槠林RS的关键因子,但湿度因子的加入也提高了模型的准确性。
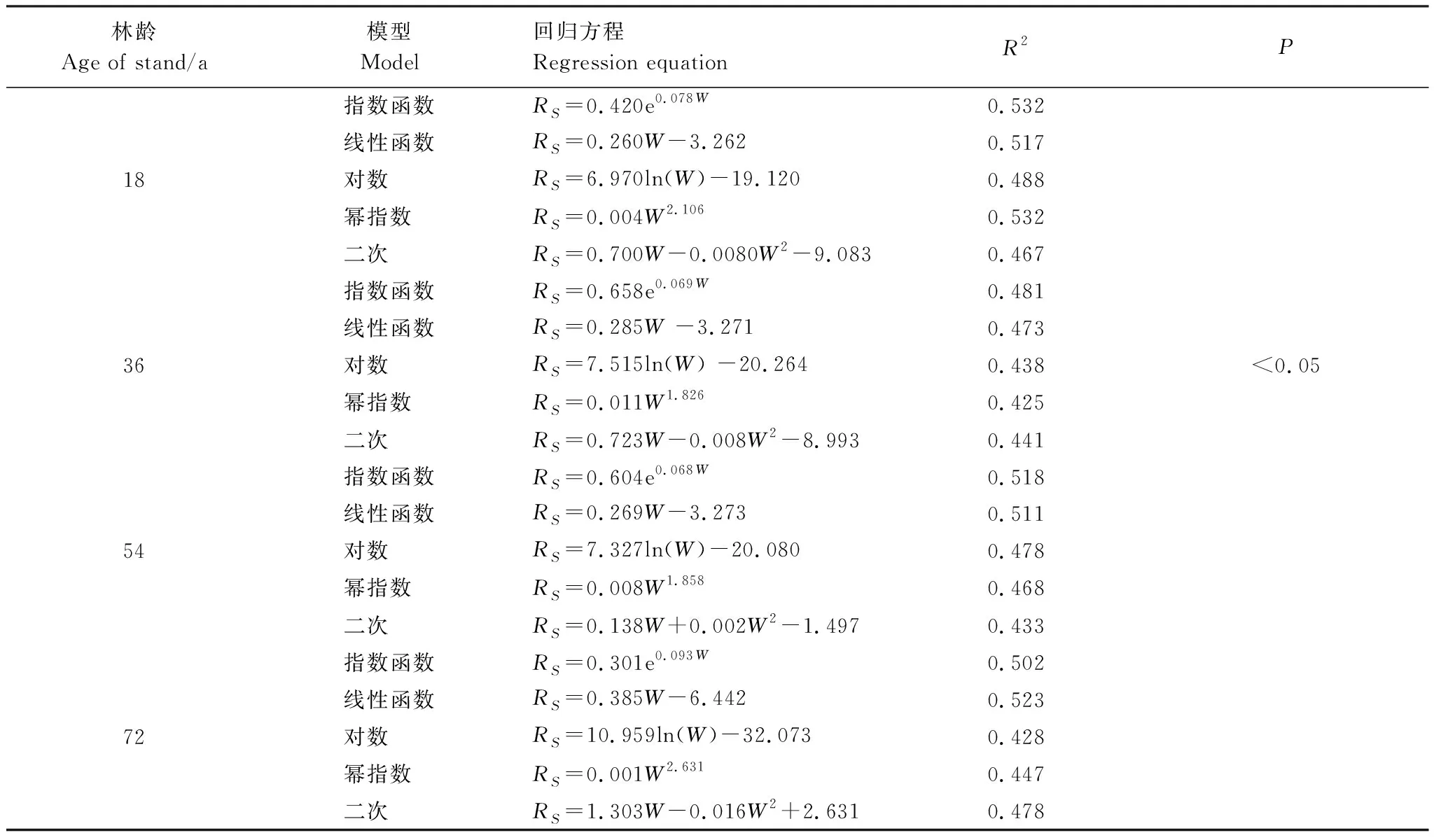
表4 土壤含水量与土壤呼吸速率拟合方程
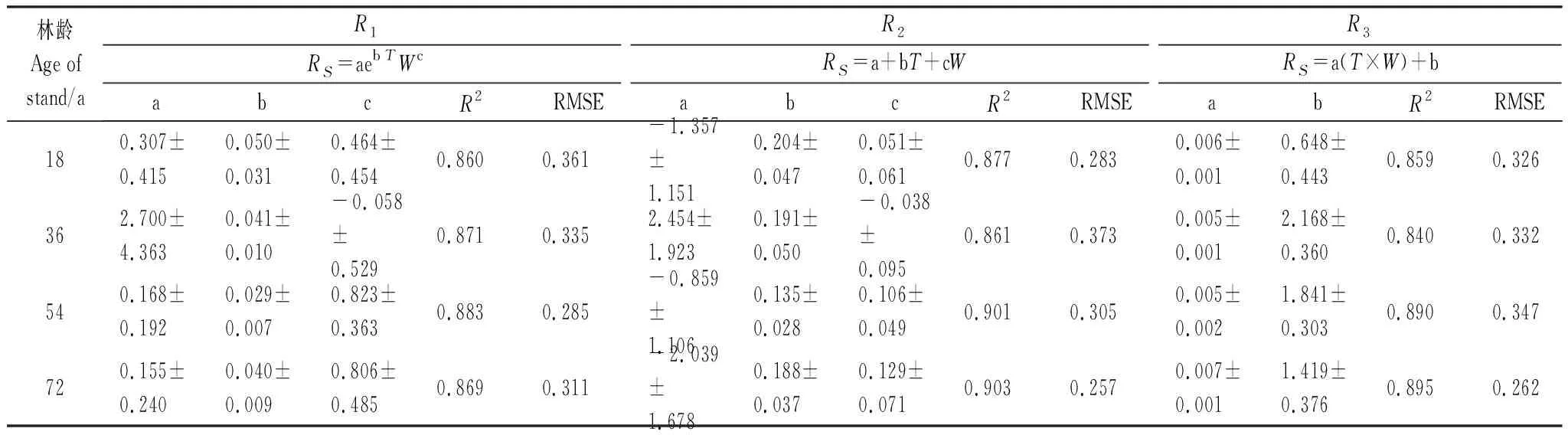
表5 5 cm深土壤温度、含水量与土壤呼吸速率拟合方程
18、36、54、72 a甜槠林实测值与预测值误差范围分别为:±1.5 μmol m-2s-1、±1.5 μmol m-2s-1、±1 μmol m-2s-1、±2 μmol m-2s-1。总体来看36 a预测效果最好,其次为18 a。同时可以看出,用双变量预测模型预测RS时,可以克服单变量指数模型在土壤温度较高时对土壤呼吸速率值的高估以及土壤水分较高时对RS的低估。
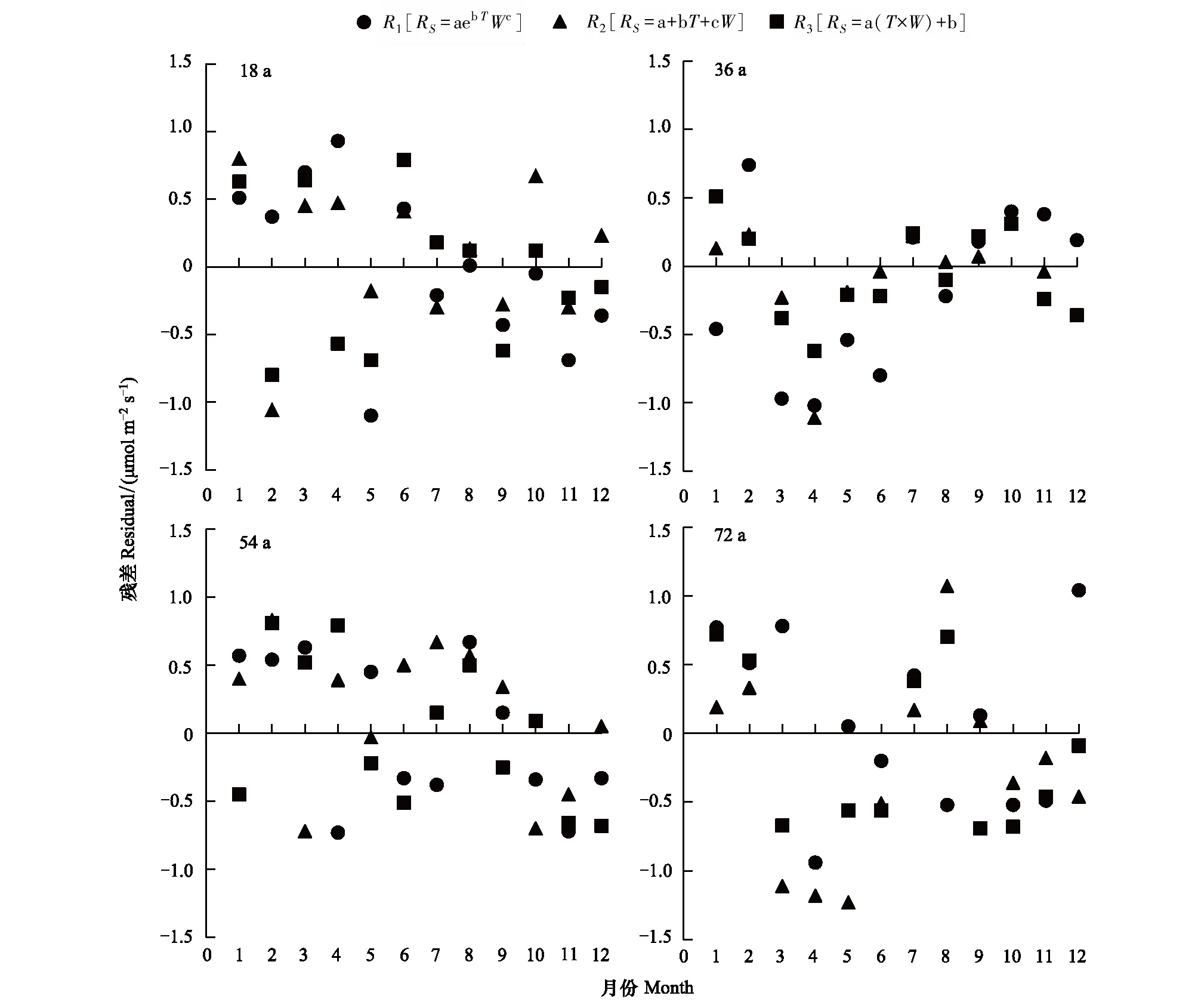
图3 不同林龄甜槠林土壤呼吸速率拟合残差值Fig.3 Fitting residual difference of soil respiration rates in Castanopsis eyrei forests with different stand ages
2.3 不同林龄甜槠林细根生物量分布及其与土壤呼吸速率关系
4种林龄甜槠林细根生物量季节变化基本趋势大致为:夏季>秋季>春季>冬季(图4)。四个季节中,0—20cm、20—40cm土层细根生物量均为36 a甜槠林最大,这与RS最大值出现在36 a甜槠林中具有相似表现。甜槠林根系总生物量随林龄的增大而增加(图5),其中粗根生物量变化趋势与根系生物量变化趋势相同,细根生物量随林龄的增大呈单峰趋势,表现为从18 a起,甜槠林细根生物量开始增加,到36 a达到细根生物量最大值,而后细根生物量逐渐降低。甜槠林细根生物量在0—20cm处最大(图6),表明甜槠林细根主要集中在表层土壤。
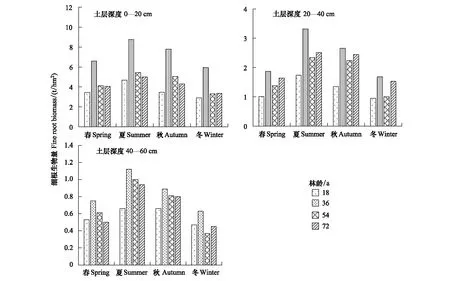
图4 不同林龄甜槠林不同土层深度根系生物量季节变化Fig.4 Seasonal changes of root biomass in Castanopsis eyrei forests with different forest ages and different soil depth
RS与各林龄甜槠林不同土层细根生物量相关性如表6所示,除18 a甜槠林外,其余3种林龄甜槠林各土层细根生物量与土壤呼吸速率拟合R2均在0.60以上,36 a甜槠林各土层细根生物量与RS在四种林龄中拟合最好。0—20 cm土层细根生物量对土壤呼吸速率的解释率54 a(70.2%)、72 a(70.4%)高于20—40 cm土层细根生物量对土壤呼吸速率解释率54 a(65.1%)、72 a(63.1%),36 a甜槠林0—20 cm、40—60 cm土层细根生物量对土壤呼吸速率解释率相差不大。0—60 cm土层根系生物量与5 cm深土壤温度可共同解释了RS变异的88.3%—91.8%,高于5 cm深土壤温度、含水量与RS的拟合值(表7)。
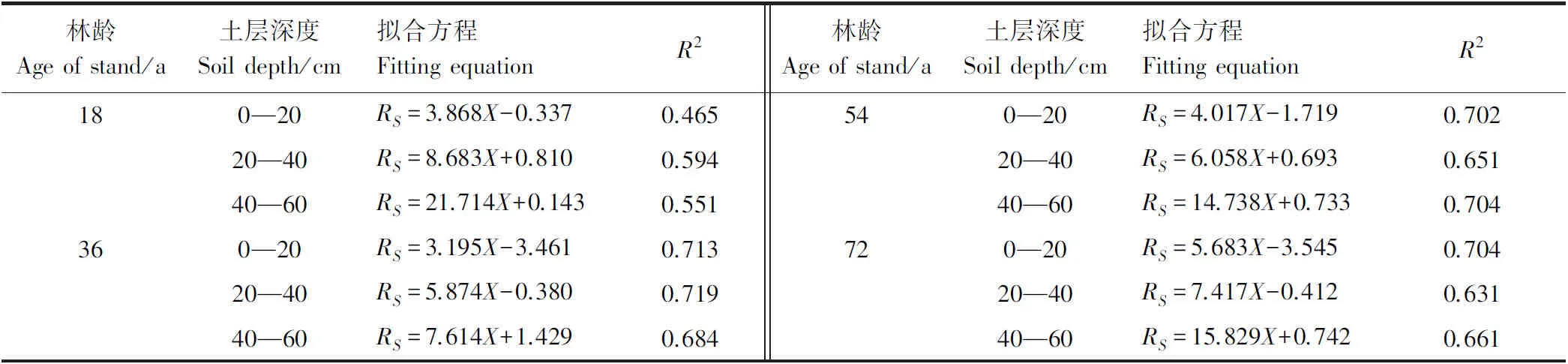
表6 不同林龄甜槠林各土层深度细根生物量与土壤呼吸速率表线性关系
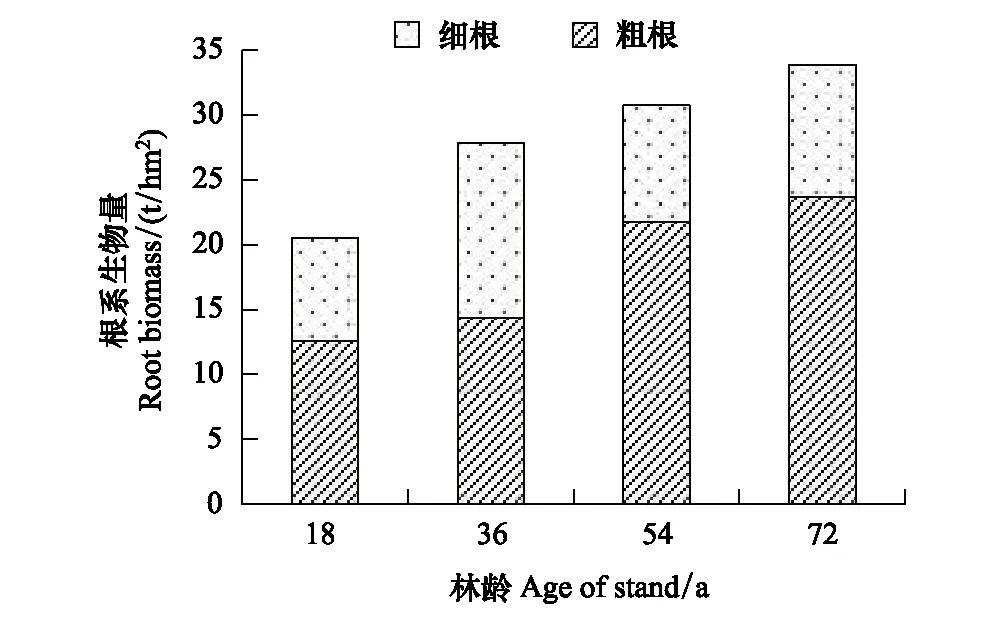
图5 不同林龄甜槠林根系生物量 Fig.5 Root biomass of Castanopsis eyrei forests in different forests
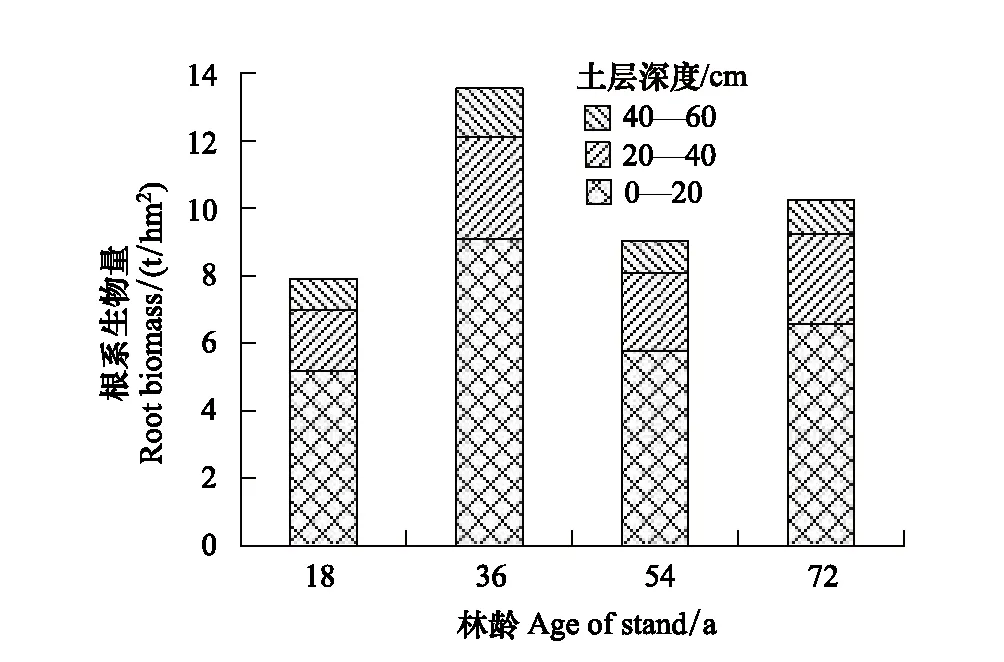
图6 不同林龄甜槠林细根生物量垂直分布 Fig.6 Vertical distribution of fine root biomass of Castanopsis eyrei forests in different forests
3 讨论与结论
3.1 不同林龄甜槠天然林土壤呼吸速率变化及其主要驱动因子
本研究表明4种林龄甜槠天然林RS呈单峰型曲线,表现为夏季(6、7月)最高,冬季最低(1月),且与土壤温度具有一致的季节变化特征,这与亚热带森林RS研究结果一致[22,27],可能与研究区内,不同林龄间气温、降水量等气候条件基本相同,土壤温度、土壤含水量季节变化模式也基本相同有关。不同林龄甜槠林RS变化与季节因素无关(P>0.05),这与韩营营对不同林龄白桦天然林RS研究结果一致[28]。相比之下,橡树林随林龄增加RS降低[29],山杨人工林也呈相同的趋势[30],这表明随温度带不同、树种不同、微环境不同,林龄对RS的响应可能存在差异。不同林龄甜槠林秋季土壤异养呼吸占总呼吸的比重最高,可能与研究区秋季气温开始降低,自养呼吸占总呼吸的比例下降(表2),同时凋落物增加(图1)为土壤动物、微生物提供了大量的碳底物[31],微生物分解凋落物释放CO2增多有关。
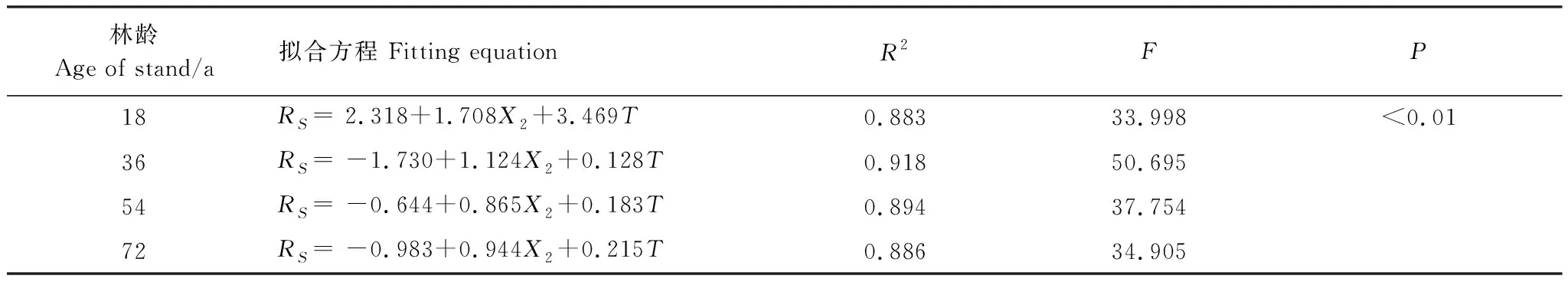
表7 5 cm深土壤温度、0—60 cm细根生物量与土壤呼吸速率线性关系
土壤温度被认为是解释不同林龄RS差异的一个重要因素。本研究中4种林龄甜槠天然林RS与土壤温度间存在极显著的指数关系(P<0.01),土壤温度可以解释不同林龄甜槠林RS的80.26%—86.22%变异,不同林龄甜槠林土壤温度对RS的影响显著高于土壤含水量的影响,表明在该区域中土壤温度是影响RS的关键因子,这与中亚热带常绿阔叶林的研究结果一致[32]。随着林龄的增加,一方面甜槠天然林乔木层植物个体、密度增大,林分郁闭度和密度通过影响植被林下光照对土壤温度产生影响[33]。4种林龄中,18 a甜槠林5 cm深土壤温度明显高于其他3种林龄(P<0.05),这可能是因为18 a甜槠林冠层盖度较低(表1),而冠层盖度高低直接影响了土壤表面可接收的太阳辐射[34],使得土壤温度、土壤有机碳密度、微生物根系呼吸产生差异,进而影响RS变化,前人研究表明冠层盖度与土壤温度间存在极显著关系,且在土壤较为干旱时,冠层盖度对RS的影响最大[35]。另一方面,凋落物分解产生的土壤有机质是有机碳补充的重要来源,随着林龄的增大,植被凋落物量不断增多[31],促进了土壤有机碳的积累,为微生物呼吸提供了底物,使得RS增大。36 a甜槠林土壤呼吸速率最高,这可能是因为36 a甜槠林林下凋落物丰富,冠层盖度较18 a甜槠林高,但5 cm深土壤温度较54、72 a甜槠林低,很可能通过增加凋落物产量对根系、微生物呼吸产生影响,进而影响RS。
Q10值是用来表征RS对温度变化的敏感性指标,众多研究表明,Q10值随时空变化存在差异[36- 37]。本研究结果表明,不同林龄甜槠林Q10值在1.551—1.664之间,这处在亚热带森林土壤呼吸Q10值(1.56—2.39)范围中[38]。72、54 a甜槠林Q10值显著高于36、18 a甜槠林Q10值,且与土壤温度变化呈负相关关系,表现为Q10值在高温下较低,低温下较高,表明低温地区植被土壤呼吸对温度变化更为敏感,这与前人研究结果一致[23,38]。结果表明54、72 a甜槠林Q10值显著高于18、36 a甜槠林,这与韩营营等[28]、YU等[33]研究结果一致,但与于舒[39]对不同林龄油松次生林Q10值随林龄的增加而降低不一致。这些结果都强调了林龄因素对Q10值的影响,因此,在不考虑林龄因素的影响下,土壤温度与RS的拟合估计值可能会低估或高估于实测值。
本研究中,土壤含水量仅可以解释甜槠林RS变异的42%—54%,可见,研究区内甜槠林湿度因子对RS的影响较小。这与相关学者研究结果不一致,刘鹏等表明当土壤含水率大于8%时,RS与土壤含水率无显著相关关系[40];刘宝等在对3种林龄闽楠林RS研究发现土壤含水率与RS之间不存在显著线性相关关系(P>0.05)[41]。本研究中土壤含水量与RS间相关关系拟合程度不高,很可能是因为研究区中有丰富的降水,土壤含水量高,湿度因素不是该区域的限制因素,但也发现54、72 a甜槠林较36 a甜槠林土壤含水量高时,RS有所降低,表明随着林龄增加有机质积累量增大,土壤容重降低,持水能力增高时,可能会抑制土壤根系呼吸[42],使得RS降低,但其阈值需要进一步确认。湿度因子与土壤呼吸间的关系复杂,很难单独分析水分对RS的影响,一般而言,湿度较小的地区如干旱半干旱地区,土壤湿度与RS拟合较好,温湿度双因素拟合对RS解释性及预测性更强[43]。
3.2 不同林龄甜槠林根系生物量变化及其对土壤呼吸的影响
本研究表明随着甜槠林林龄的增长,根系生物量(细根+粗根)呈增大趋势,0—60 cm细根生物量表现为随林龄增大,呈先增加后减小的趋势。影响细根分布因素一方面与树种、土壤物理性质(通气性)、土壤水分等有关[44];另一方面,土壤温度随土层的改变也是导致细根主要分布于土壤表层的重要原因[45],0—20 cm土壤表层温度条件适宜,通气性好,有机质含量高,适宜土壤动物及微生物的生长和代谢,细根生物量多分布于此层。李帅锋等[46]研究不同林龄思茅松人工林根系生物量认为,由于群落结构、物种组成变化,思茅松细根生物量随林龄增大而减小。本研究结果与此结论不同,36 a甜槠林细根生物量在0—20 cm土层占比最大,随着林龄增大细根生物量占比逐渐减小,这可能是由于36 a甜槠林林下植被凋落量大,表层营养丰富,利于细根生长,随着林龄增大,细根向深处生长需要一定的时间;同时,36 a甜槠林的林下植被相较于54、72 a甜槠林植被生长好,表层土壤养分含量高,因而细根生物量大。土壤各层次细根生物量的分布差异反映出土壤资源有效性的变化情况[46],森林土壤资源有效性的变化会直接导致地下碳分配格局的改变,进而影响RS的动态变化。
不同林龄甜槠林不同深度土层细根生物量与RS拟合结果表明,RS与各层土壤根系生物量呈线性正相关关系,除18 a甜槠林外,其余三种林龄各土层细根生物量与RS拟合R2均在0.60以上,表层土壤(0—20 cm)细根生物量是引起地下根系生物量差异的主要原因。相关研究表明,亚热带常绿阔叶林根系呼吸速率与细根生物量呈极显著相关关系[47],夏季适宜的环境因子促进细根生物量生长,使得土壤呼吸速率与细根生物量达到最大值[48]。本研究表明试验区内0—60 cm土壤细根生物量、土壤温度与RS拟合高于土壤温度、含水量与RS拟合值,土壤根系生物量相较于土壤含水量对甜槠林RS影响更大。
RS具有明显的时空异质性,在空间上彼此接近的同种植被,测定的土壤呼吸速率都可能存在很大的差异。本研究分析了武夷山自然保护区4种不同林龄甜槠林RS的年际变化及影响RS的生物、非生物因子,由于本实验仅在同一海拔、同一立地条件对不同林龄甜槠林进行土壤呼吸研究,因此所得结果与武夷山自然保护区甜槠林立地特异性有关。本研究强调了甜槠林RS中林龄的作用,指出在对未来森林植被土壤呼吸及碳汇功能研究时,应考虑林龄及生物因子的影响。
