茶树3个MAPKKK基因的克隆及其在采后加工中的表达
2021-04-22沈潮如杨润梅曹红利
沈潮如 杨润梅 岳 川 曹红利
(福建农林大学园艺学院/茶学福建省高校重点实验室,福建 福州 350002)
在茶叶加工过程中,萎凋是白茶品质形成的关键工序,做青是乌龙茶形成天然花果香的关键。在萎凋、做青等工序中,鲜叶受光照、高温、脱水、翻(抖)动等胁迫产生应激响应,水分逐渐散失,茶叶中的酶发挥作用,咖啡碱、茶多酚等物质发生转化,茶叶品质逐渐形成。研究表明这些加工应激响应过程是构成茶叶品质风味形成的关键因素[1]。目前关于白茶萎凋和乌龙茶做青的研究已有较多报道,多集中在不同萎凋方式和条件、不同做青工序等条件下理化成分变化以及香气形成机理[2-3]。随着技术的发展近年来的研究热点集中在利用多组学对白茶萎凋及乌龙茶做青过程品质形成机理进行探究[4-5]。大量研究发现,茶叶中与香气等品质相关的基因表达会在白茶萎凋和乌龙茶加工中具有显著变化,推测这些基因参与了茶叶采后加工的品质形成[6-9]。然而,加工过程中外界刺激与品质形成间的分子响应调控模式仍未知,因此,在明确与品质相关基因在加工过程中的表达模式后,有必要进一步深入研究茶叶品质形成调控的信号转导通路。
丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)级联是动植物中广泛存在的保守信号转导通路,主要由MAPKKK、MAPKK和MAPK的依次磷酸化作用将信号级联放大,通过逐级磷酸化将外界刺激信号转导至细胞内部,以级联顺序磷酸化和活化不同的上游受体和下游靶基因来响应胁迫[10-12]。MAPKKK是一类丝氨酸(Ser)/苏氨酸(Thr)激酶,位于MAPK级联上游,数量最庞大,通过磷酸化下游MAPKK活化环S/T-XXXXX-S/T基序中S/T残基激活MAPKK。根据MAPKKK催化结构域的不同,植物中MAPKKK分为MEKK亚族、Raf亚族和ZIK亚族,其保守域分别为G(T/S)Px(W/Y/F)MAPEV、GTxx(W/Y)MAPE和GTPEFMAPE(L/V)(Y/F)[13-14],研究发现MEKK亚族的蛋白质结构保守性与ZIK亚族和Raf亚族相比相对较低[15]。大量研究表明,MAPKKK基因家族在植物应对干旱、高盐、低温等逆境中发挥重要作用[16-17],在气孔发育[18]、细胞分裂[19]等过程中也发挥着重要的调节作用。MAPKKK的研究主要集中于模式植物和大田作物,如水稻(Oryzaativa)[14]、拟南芥(Arabidopsisthaliana)[18]、玉米(Zeamays)[20]、棉花(Gossypiumhirsutum)[21]、大豆(Glycinemax)[22]等。目前,茶树(Camelliasinensis)中关于MAPK级联途径基因的报道较少,本研究前期发现CsMAPK3参与茶树非生物胁迫响应[23],但茶树中大量的相关基因有待挖掘,同时它们在茶叶采后加工过程中刺激响应与茶叶品质形成的调控作用有待揭示。
本研究以福鼎大白茶茶树和铁观音茶树品种为材料,克隆获得3个MAPKKK基因cDNA全长序列,对其生物信息学进行全面分析,采用荧光定量PCR检测其因的组织表达特异性以及在白茶萎凋和乌龙茶做青过程中的表达,旨在为揭示茶叶采后加工品质形成的调控机理提供理论参考。
1 材料与方法
1.1 试验材料
以福建农林大学园艺学院南区教学茶场福鼎大白茶为材料,于2018年5月采取一芽二叶为原料,放置室内自然萎凋,室温控制在22~25℃,空气相对湿度70%左右。在萎凋过程的0、0.25、1、4、12、24、36和48 h进行取样。以铁观音中小开面二三叶嫩梢为材料,按照闽南清香型乌龙茶加工工艺进行制作(表1),在此过程中取鲜叶、晒青叶、一摇、二摇、三摇和杀青前等各工序的在制品为样品。于2018年10月分别采取盆栽福鼎大白茶茶树植株的根、茎、叶、花、果实,作为分析不同组织基因表达材料。所有上述样品设置3次生物学重复,取样后用锡箔纸包裹液氮速冻,放-80℃冰箱保存备用。
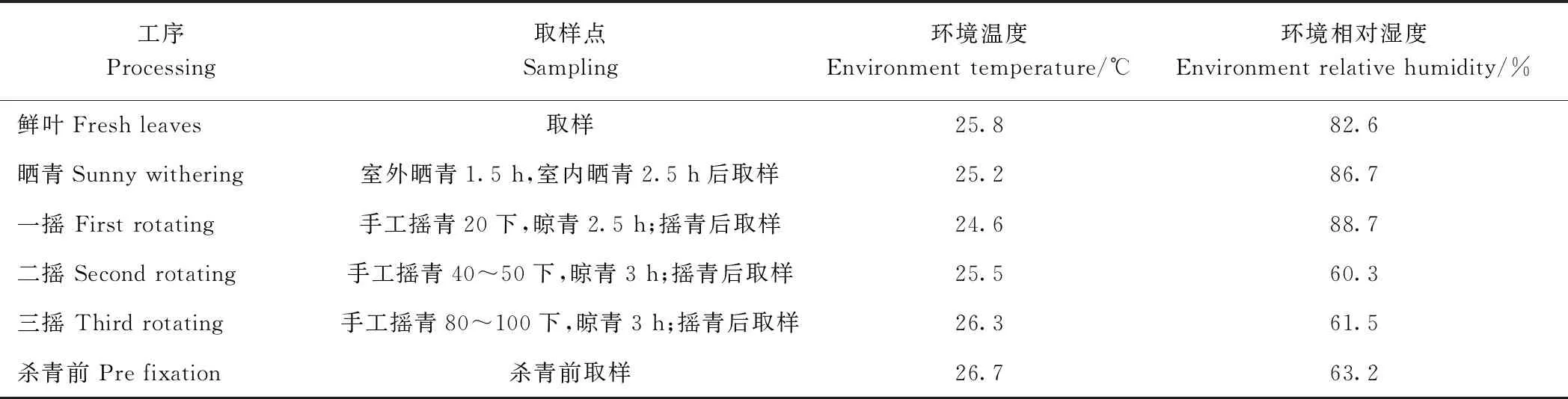
表1 乌龙茶加工工艺
GeIDoc-ItTS3Imager凝胶成像仪、CFX96 Touch荧光定量PCR仪、T100TMThermal Cycler PCR仪,美国Bio-Rad公司;琼脂糖水平电泳仪和电泳槽,北京六一公司;AIRTECH 紫外超净工作台,苏州安泰公司;NanoDrop ND2000 超微量分光光度计,上海Thermo Scientific公司。
1.2 试验方法
1.2.1 总RNA的提取与cDNA 的合成 按照RNAprep Pure 多糖多酚植物RNA提取试剂盒(北京天根生化科技有限公司)说明,取0.1 g样品在液氮中低温研磨后,提取茶树总RNA。采用PrimeScriptTM1st strand cDNA Synthesis Kit 试剂盒(日本TaKaRa公司)合成cDNA用于后续反转录PCR(reverse transcription PCR, RT-PCR);采用Easy Script @One-Step gDNA Removal and cDNA Synthesis SuperMix试剂盒(北京全式金生物技术有限公司)合成cDNA模板用于实时荧光定量PCR(real-time quantitative PCR, qRT-PCR)试验。
1.2.2CsMAPKKK基因cDNA全长克隆 根据前期转录组数据的注释结果[24],筛选茶树MAPKKK基因序列,序列经拼接后再进行Blastx同源比对,并结合DNAMAN软件查找开放阅读框(open reading frame, ORF)。在ORF上下游设计引物(表2),以1.2.1节所得cDNA为模板,进行PCR扩增。按照PrimeSTAR HS试剂盒(日本TaKaRa公司)操作步骤进行,程序为98℃预变性5 min;98℃变性10 s,56℃退火10 s,72℃延伸3 min,30个循环;72℃延伸5 min。PCR产物经1.0%琼脂糖凝胶电泳,用凝胶成像系统(美国Bio-Rad公司)检测和记录结果。扩增产物回收,转入大肠杆菌,筛选阳性克隆进行基因测序,测序结果通过NCBI数据库比对,最终获得基因的完整编码序列。
1.2.3 CsMAPKKK生物信息学分析 采用ProtParam(http://expasy.org/tools)在线软件分析预测蛋白理化性质、跨膜结构域。用Signal P5.0预测信号肽利用WoLF PSORTS(https://www.genscript.com/)进行亚细胞定位。利用MEME(http://meme-suite.org/tools/meme)预测保守元件。使用SOPMA(https://npsa-prabi.ibcp.fr)和Swiss Model(https://swissmodel.expasy.org/)分析三级结构。通过NCBI-CDD和SMART分析保守结构域,使用DNAMAN 8.0软件进行多序列比对。利用MEGA7.0软件,采用邻接法(neighbor-joining,NJ)构建系统发育进化树。
1.2.4CsMAPKKK基因表达分析 以茶树不同组织、不同萎凋和做青过程的样品总RNA为模板,用Easyscript One-step gDNA Removal and cDNA synthesis superMix 试剂盒(北京全式金生物技术有限公司)合成cDNA,以茶树CsPTB为内参基因,设计qRT-PCR引物(表2),测定目标基因的表达水平变化。采用Bio-Rad的CFX96 Touch荧光定量PCR仪进行扩增,反应体系参照transstart Tip Green qPCR superMix试剂盒(北京全式金生物技术有限公司)方法,程序:94℃预变性30 s;94℃变性5 s;60℃退火30 s;40个循环。结果采用2-ΔΔCt算法进行分析。采用Excel 2016软件分析数据及作图,SPSS 22.0进行显著性差异分析(P<0.05)。
2 结果与分析
2.1 CsMAPKKK基因克隆与理化性质分析
如图1所示,通过ORF上下游引物进行qRT-PCR扩增获得3个CsMAPKKK基因全长。在NCBI数据库比对出它们有完整的ORF,且与其他物种的MAPKKK18、MAPKKK-NPK1具有较高的相似性,将3个CsMAPKKK基因分别命名为CsMAPKKK18、CsMAPKKK18-like和CsMAPKKK-NPK1,cDNA全长序列分别为1 005、1 209和1 077 bp。 在ProtParam中对3个CsMAPKKK氨基酸序列进行预测分析,结果显示如表2所示。CsMAPKKK18、CsMAPKKK18-like及CsMAPKKK-NPK1分别编码334、402和358个氨基酸;分子量分别为36.59、44.03和39.56 kDa;不稳定系数分别为46.55、44.64和47.28,均大于40,表明3个CsMAPKKK蛋白均为不稳定蛋白;亲水性(grand average of hydropathy, GRAVY)系数分别为-0.381、-0.298和-0.307,表明均为疏水性蛋白;理论等电点(pI)分别为6.01、5.16和7.09,CsMAPKKK18和CsMAPKKK18-like蛋白的pI小于7.0,为酸性蛋白。TMTHMM跨膜结构预测显示CsMAPKKK18、CsMAPKKK18-like与CsMAPKKK-NPK1均不含有跨膜结构域;信号肽输出位点预测显示只有CsMAPKKK18和CsMAPKKK18-like有信号肽输出位点;磷酸化位点预测显示分别含有31、46和32个Ser、Thr或Tyr位点;WoLF PSORTⅡ预测亚细胞定位结果表明最大可能分别定位于液泡、线粒体和细胞核。

表2 基因引物序列

注:1:CsMAPKKK18-like;2:CsMAPKKK18-NPK1;3:CsMAPKKK18;M: Marker 2 000。
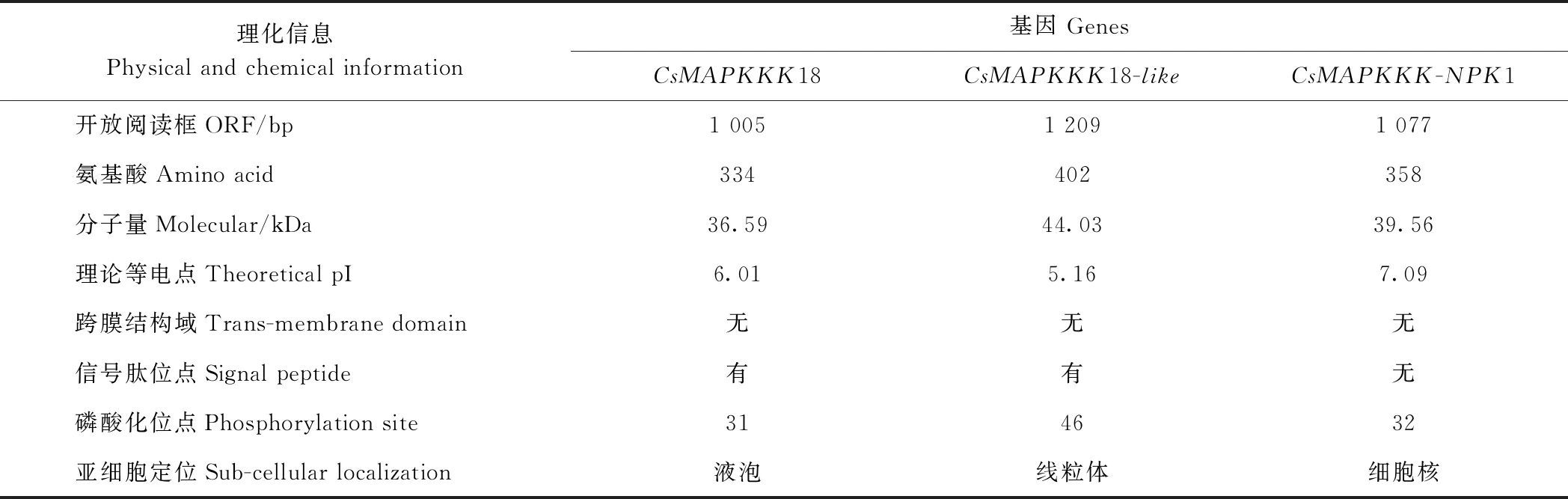
表3 CsMAPKKKs基本理化信息
2.2 CsMAPKKK保守结构域分析
氨基酸多重比对分析结果如图2所示,CsMAPKKK同蓖麻、李、榴莲、葡萄具有较高的保守性,均具有MEKK特征性保守区域G(T/S)Px(W/Y/F)MAPEV(图中黑色框),其中CsMAPKKK18与CsMAPKKK18-like保守域序列均为GTPMFMAPEV,CsMAPKKK-NPK1保守域序列为GTPLWMAPEV。除了特征性保守域,还利用MEME软件对其他结构域进行预测,结果显示(图3),CsMAPKKK18与CsMAPKKK18-like在结构上更相似,分别含有8个和10个保守域,CsMAPKKK-NPK1含有9个保守域。
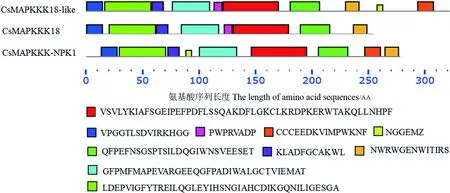
图3 茶树3个CsMAPKKK蛋白的MEME保守元件预测
2.3 CsMAPKKK三级结构预测分析
用SWISS-MODEL对MAPKKK蛋白的三级结构进行模拟(图4),根据拟南芥MEKK蛋白为模板建模,其序列一致性在35%~40%之间,且全球模型质量估计值(global model quality estimation, GMQE)值为0.5~0.6,具有理想的预测效果。3个CsMAPKKK蛋白三级结构高度相似,均具有MAPKKK典型的三级结构,图中黑色箭头分别指示其MEKK特征结构域。
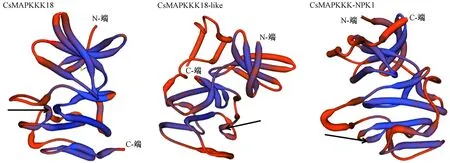
注:黑色箭头指示保守结构域G(T/S)PX(W/Y/F)MAPEV。
2.4 GsMAPKKK的进化树分析
以蓖麻(Ricinuscommunis)、可可(Theobromacacao)、葡萄(Vitisvinifera)、番茄(Solanumlycopersicum)、甜椒(Capsicumannuum)等11个植物的同源MAPKKK蛋白和3个茶树的CsMAPKKK蛋白构建进化树分析,结果显示(图5),CsMAPKKK-NPK1与蓖麻、葡萄、栓皮栎树、榴莲、可可、梅、李聚为一类,与蓖麻和葡萄关系最近;CsMAPKKK18-like与CsMAPKKK18聚为一类,相似性达85%,这两个蛋白与番茄、甜椒等亲缘关系较近。

图5 茶树3个MAPKKK蛋白与其他植物MAPKKK基因编码的氨基酸序列的系统进化树
2.5 CsMAPKKK基因茶树不同组织部位中的表达分析
由图6可知,CsMAPKKK18在根部的表达量最高,其次是花,都显著高于茎、叶和果中的表达量,其中叶和果中的表达无显著差异且表达量都很低。CsMAPKKK18-like的表达模式与CsMAPKKK18相似,根中表达量最高,其次是花,茎、叶、果中的表达无显著差异且三者的表达量极低。CsMAPKKK-NPK1同样在根中的表达最高,显著高于茎、叶、花和果中的表达量,且在茎、叶、花和果中的表达无显著差异。
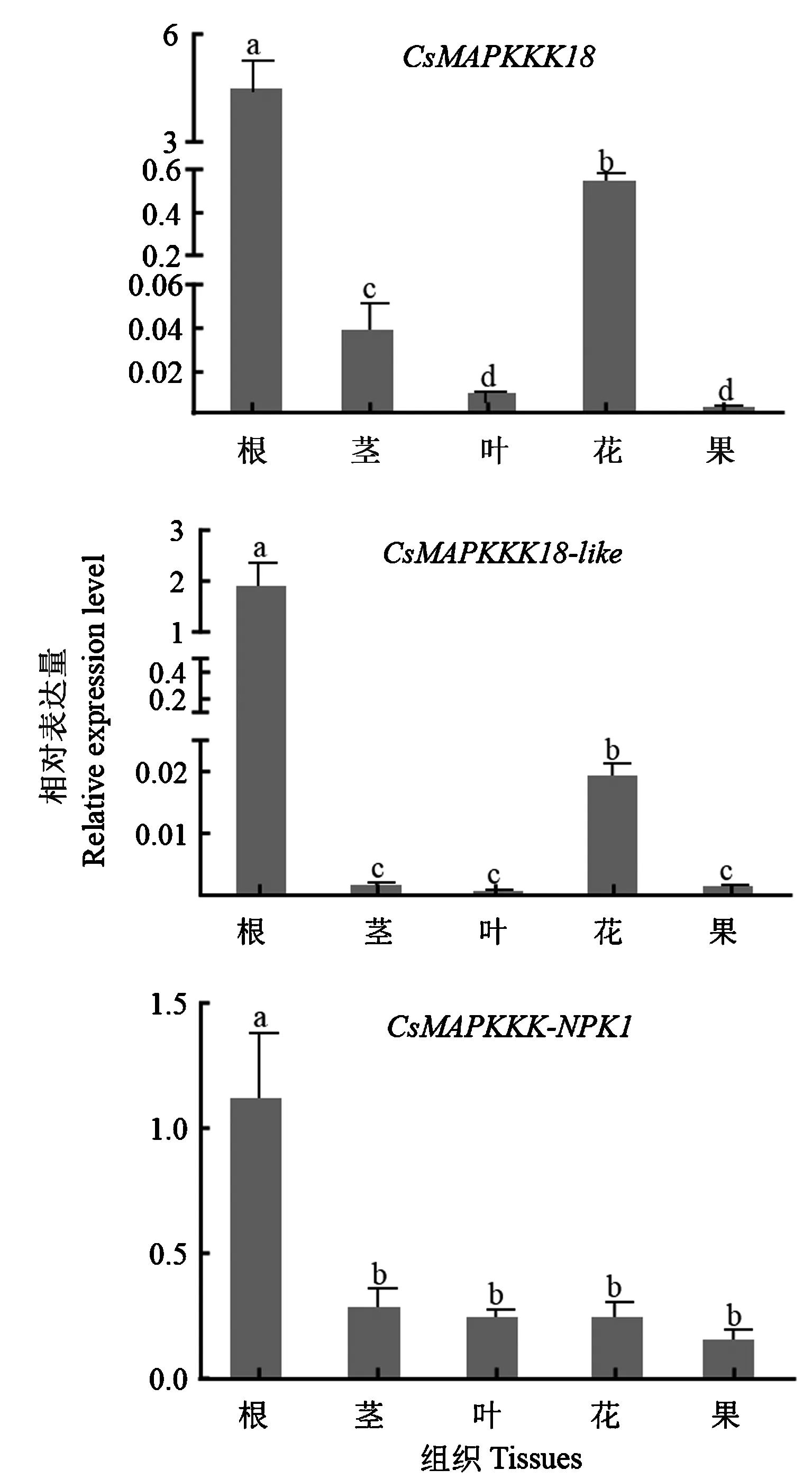
注:不同小写字母表示显著性差异(P<0.05)。下同。
2.6 CsMAPKKK白茶不同萎凋时间的表达分析
在白茶萎凋过程中的表达结果显示(图7),以鲜叶(0 h)为对照,CsMAPKKK18在其他各萎凋阶段均显著上调,且在萎凋1 h表达量显著上调至64.9,但在4 h时表达量迅速下调至5.8,而12 h 时表达量又迅速上调至64.3,之后表达量下降,但表达水平都显著高于0 h。CsMAPKKK18-like表达量呈现先升后降的趋势,当萎凋时间为1 h 时表达量最高,达3.44,而后表达量迅速下降,到24 h以后表达量显著低于鲜叶(0 h)。CsMAPKKK-NPK1在萎凋前期(4 h以前)表达量上调缓慢,在萎凋12 h以后表达量迅速上调,到48 h 时表达量最高,约为0 h的30倍。综上说明CsMAPKKK18和CsMAPKKK-NPK1在白茶萎凋0~48 h的过程中受诱导上调表达,而CsMAPKKK18-like在萎凋前期诱导,后期抑制表达。

图7 茶树3个CsMAPKKK在白茶萎凋过程中的表达
2.7 CsMAPKKK基因在乌龙茶做青过程中的表达分析
在乌龙茶做青过程中,以鲜叶为对照,荧光定量PCR表达结果显示(图8),CsMAPKKK18表达量在晒青后显著下调为0.3,而一摇、二摇、三摇和杀青前的表达量都是显著上调,且在二摇和三摇分别上调至23.1和26.2;CsMAPKKK18-like在三摇表达量显著上调为1.6,但在晒青、二摇和杀青前显著下调表达,表达量分别为0.4、0.5和0.5;CsMAPKKK-NPK1在晒青显著下调到0.3,在一摇、二摇和三摇都显著逐渐上调,其表达量分别为1.7、2.1和3.2,推测CsMAPKKK18和CsMAPKKK-NPK1与乌龙茶做青过程中机械力胁迫诱导下茶叶品质的形成密切相关。
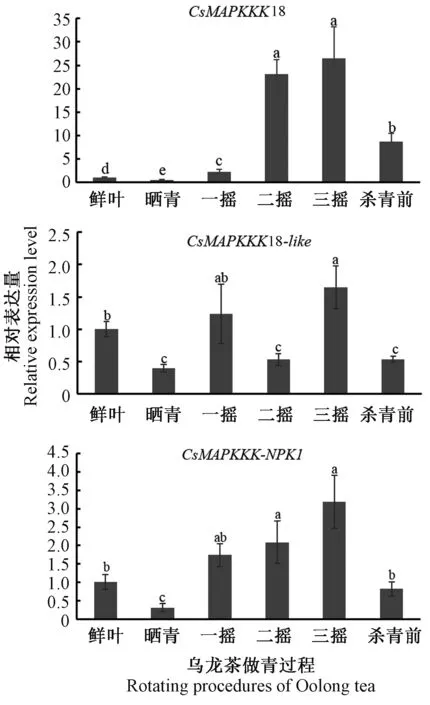
图8 茶树3个CsMAPKKK在乌龙茶做青过程中的表达
3 讨论
MAPKKK家族是MAPK级联途径中成员最多的一个基因家族,多种植物已从全基因组数据中鉴定出完整的MAPKKK家族成员,如拟南芥中含80个[13],水稻中含75个[25],小麦中含155个[26],玉米中含74个[20],葡萄中含45个[27-28],棉花中含78个[21],番茄中含89个[29],黄瓜中含59个[30]。目前,仅报道茶树1个CsMAPK3基因[23],而茶树MAPKK和MAPKKK基因鲜见报道。因此,茶树中还有大量MAPKKK家族成员待进一步挖掘鉴定。本研究获得3个茶树MAPKKK基因,它们均具有MEKK特征性保守区域G(T/S)PX(W/Y/F)MAPEV,属于MEKK亚家族。研究表明,大部分ZIK亚族在N端具有激活域,几乎所有的Raf亚族在C端具有激活域,N端具有调节域,而MEKK亚族的激活域有可能在N端、C端或中央[31],CsMAPKKK激活域有待进一步研究。亚细胞定位预测显示,3个CsMAPKKK蛋白都定位于不同亚细胞结构上,推测这3个基因在功能上有差异。
本研究检测了这3个CsMAPKKK在茶树不同组织部位的表达,结果显示,3个CsMAPKKK在茶树根、茎、叶、花以及果等组织中均有表达,且均是在根中表达量最高。该结果与番茄[29]、黄瓜[30]、小麦[26]等研究中的MAPKKK组织特异性表达结果一致。其中CsMAPKKK18与CsMAPKKK18-like的组织表达模式具有相似性,根和花的表达量显著高于其他组织,推测CsMAPKKK18与CsMAPKKK18-like具有功能上的相似性。
进一步分析了3个基因在白茶萎凋和乌龙茶做青过程中的表达模式,为深入揭示茶叶采后加工品质形成机理提供参考。在白茶0~48 h萎凋过程中,3个CsMAPKKK均受诱导表达。CsMAPKKK18和CsMAPKKK18-like在萎凋前期(1 h)的表达量最高,其中CsMAPKKK18在4 h表达量骤降,12 h表达量又上升到高水平,此后表达量降低但保持稳定,而CsMAPKKK18-like在4 h后表达量显著下调。CsMAPKKK-NPK1在白茶萎凋前期表达量上调缓慢,12 h后表达量增加最多达30倍。在乌龙茶做青过程的5个时间点,3个CsMAPKKK都有不同程度的诱导。其中CsMAPKKK18表达量上调最大达26倍;CsMAPKKK18-like在三摇显著上调,而晒青、二摇和杀青前均为显著下调;CsMAPKKK-NPK1在晒青显著下调,在一摇、二摇和三摇都显著上调。可见这3个基因的表达水平变化与茶叶采后加工的进程密切相关。白茶萎凋和乌龙茶做青过程中,茶叶所受的刺激模式不尽相同,白茶萎凋主要以水分散失导致的渗透胁迫为主,而乌龙茶做青中既有失水产生的胁迫,也有机械力的作用。MAPK级联在渗透胁迫和机械力胁迫中发挥着重要的作用,因此,这些基因的上调表达能够增强茶叶自身对外界刺激的响应。同时,蛋白激酶等编码基因的上调表达,也可能在茶叶品质形成中发挥重要的调控作用。前期研究发现,乌龙茶做青和白茶萎凋中,多种与茶叶品质相关的基因的表达均上调,如β-淀粉酶基因(CsBAMs)、类胡萝卜素裂解双加氧酶基因(CsCCDs)、类胡萝卜素裂解双加氧酶(CsNCED2)等[6-7,32]。做青机械振动能显著提高茶树橙花叔醇合成酶CsNES基因和芳樟醇合成酶CsLIS的表达[33-34],茶树萜烯类合成相关基因CsMVK,CsDXS和CsGGPS在白茶萎凋过程中显著上调[9]。茶叶加工过程中,茶叶会产生不同程度的渗透胁迫,逐渐失水,膜系统逐渐破坏,此过程中基因表达非常活跃,不同程度的非生物胁迫和生物胁迫产生的防御作用是影响萜类物质合成的主要因素,而萜类物质是构成茶叶香气、滋味等品质的重要作用因子[35]。然而,与品质相关的基因在加工过程中如何被刺激,即外界刺激如何调控茶叶鲜叶在制品品质形成的信号通路仍有待研究。MAPK级联与植物的抗逆响应密切相关,茶叶对外界胁迫的响应促进了茶叶品质的形成,因此,推测由MAPK级联调控的应激响应通路对茶叶采后加工品质的形成发挥了重要作用。在MAPK级联作用下,调控下游与茶叶品质形成密切相关的基因的表达。
4 结论
本研究克隆到3个茶树CsMAPKKK基因家族成员CsMAPKKK18、CsMAPKKK18-like和CsMAPKKK-NPK1,生物信息学分析显示3个基因均属于MEKK亚家族成员,预测定位在不同的亚细胞结构。进化树分析显示CsMAPKKK18与CsMAPKKK18-like与番茄、甜椒亲缘关系较近,MAPKKK-NPK1与蓖麻和葡萄关系更近。3个基因在茶树根中表达较高,且在茶叶采后加工的白茶萎凋和乌龙茶做青过程中表达活跃。本研究结果为进一步研究MAPK级联途径在茶叶采后加工过程中的功能和品质机理形成提供了参考。
