五大连池火山森林群落多样性与稳定性
2021-04-21谢立红曹宏杰黄庆阳王继丰王建波倪红伟
谢立红,曹宏杰,黄庆阳,杨 帆,王继丰,王建波,倪红伟,2
(1. 黑龙江省科学院 自然与生态研究所 湿地与生态保育国家地方联合工程实验室,黑龙江 哈尔滨 150040;2. 黑龙江省林业科学院,黑龙江 哈尔滨 150040)
植物群落的空间(地带性)特征和时间(演替动态)特征都会反映在物种多样性这一重要的群落信息上[1]。森林群落的物种组成与结构是生态系统功能和过程的基础,既能反映群落的种间关系,也可表现环境对物种的影响,同时也是衡量植物群落稳定的重要尺度和方式[2]。物种多样性是物种丰富度和分布均匀性的综合反映,体现了群落的结构类型、组织水平、发展阶段、稳定程度和生境差异[3-4],分析植物群落的结构和物种多样性,对揭示群落的更新、稳定性与演替规律具有重要的意义[5]。稳定性是群落内部各个植物种群、动物种群、微生物种群、土壤环境、气候等相互作用和生物运动的结果,是在群落演替进化过程中形成和表现的[6]。稳定性是植物群落结构与功能的综合特征,是生态系统存在的必要条件和重要功能表现[7]。物种多样性和稳定性是植物群落的2个属性,它们之间的相互关系和相互影响一直受到生态学家的关注[8]。火山喷发为研究植被演替尤其是原生演替提供了难得的条件[9]。五大连池火山处于大小兴安岭和松嫩平原的交错地带,至今历经了7次火山喷发,形成了14座火山,拥有大陆上保存完整、分布集中、形态典型、种类最齐全的新老期火山地质地貌[10]。五大连池火山区生态条件独特复杂,由熔岩裸地到演化中的不同生境内可见低等植物和高等植物(演替中的不同植被生态系列)[11]。五大连池完好的内陆单成因火山地貌,原生而完整的植被演替过程,且地处植被交错区(大小兴安岭植被交错带),是研究火山干扰和植被演替与生物多样性系统发育等的理想场所[12]。近年来,对火山森林群落的物种多样性有大量研究。如牟长城等[13]研究了长白山林区森林/沼泽交错群落的植物多样性,郝占庆等[14-17]研究了长白山北坡植物群落物种多样性,姜萍等[18]研究了长白山南坡森林群落组成-结构以及树种多样性。然而,对五大连池火山森林群落的多样性和稳定性研究尚未有报道。本研究以五大连池4座老期火山为研究对象,从森林群落多样性指数、年龄结构和优势树种的存活曲线入手,分析森林群落多样性与稳定性,为五大连池火山森林植被的演替、恢复与可持续发展提供科学依据。
1 研究地区与研究方法
1.1 研究区概况
五大连池火山群 (48°30′~48°50′N,126°00′~126°45′E)中心区由新期火山活动形成的巨大熔岩流——石龙、2座年轻火山和巨大的熔岩形成的石龙台地和火山堰湖群构成,四周由老期火山活动形成的玄武岩台地构成,台地上环布12座老期火山和众多熔岩流,是中国第1个以火山自然环境及生态系统为保护对象的自然保护区[19]。
本 研 究 选 取 东 焦 得 布 山 (48°39′13ʺN,126°16′30ʺE)、 小 孤 山 (48°40′45ʺN,126°22′06ʺE)、 尾山(48°47′23ʺN,126°15′26ʺE)和南格拉球山 (48°44′13ʺN,126°00′46ʺE)4 座老期火山。研究区属温带大陆性季风气候,年平均气温-0.5 ℃,年平均降水量476.3 mm,年均无霜期121 d。研究区主要森林植被类型为温带落叶阔叶混交林,主要包括蒙古栎Quercus mongolica林和落叶阔叶林等。乔木优势树种南坡为蒙古栎和黑桦Betula davurica,北坡为紫椴Tilia amurensis和色木槭Acer mono等。研究区分布有暗棕壤性火山灰土和黑土性火山灰土[20]。
1.2 研究方法
1.2.1 样地设置与植被调查 于2018年7中旬至8月中旬植物生长旺盛期,采用样地调查法,在老期火山东焦得布山(高海拔525 m、中海拔475 m、低海拔425 m),小孤山(高海拔450 m、中海拔425 m、低海拔400 m),尾山(高海拔510 m、中海拔470 m、低海拔430 m)和南格拉球山(高海拔580 m、中海拔520 m、低海拔460 m)的南坡和北坡,每坡设置低、中、高3个海拔样地。乔木层共取24个样方,每个样方面积20 m×20 m,分别记录乔木种类、个体数、胸径、树高和群落的总郁闭度及所有乔木层树种的幼苗更新情况,用生长锥钻取胸径≥2.5 cm的乔木;灌木层分别设置4个2 m×2 m的小样方,共96个样方,记录灌木的密度、盖度、高度;草本层分别设置5个1 m×1 m的小样方,共120个样方,记录草本的密度、盖度、高度。
1.2.2 样地资料处理 将野外采取的年轮样芯,带回实验室固定在木槽内自然风干,待木芯完全风干后,用乳白胶固定在木槽上。固定后的芯样用砂粒由粗到细的砂纸打磨抛光,直到年轮清晰可见。用LINTAB年轮分析仪测年[21]。
1.2.3 多样性指数测度方法 采用Margalef、Simpson、Shannon-Wiener和Pielou等指数比较4座火山森林群落的丰富度、多样性和均匀度,多样性指数计算参考文献[3]。采用方差分析法(ANOVA)对各植物群落物种多样性指数进行差异性检验。多样性指数值均为平均值±标准误。植物种类的重要值可体现植物在群落中的相对重要性:乔木层重要值(IV1)=(相对密度+相对优势度+相对高度)/3;灌木和草本层重要值(IV2)=(相对高度+相对盖度+相对密度)/3。
1.2.4 稳定性研究方法 森林群落的年龄结构是群落变化发展的内在依据,因此,通过对森林群落年龄结构分析,可以测度群落的稳定性和动态[22]。优势种或建群种的种群稳定对群落稳定有决定作用[23]。选取群落乔木层年龄结构、乔木层重要值最高种群的年龄结构判定森林群落的稳定性。本研究中龄级划分采用胸径≥2.5 cm(利用年轮样芯测定年龄)的乔木划分,龄级划分标准以20 a为1个龄级,Ⅰ龄级为0~20 a、Ⅱ龄级为20~40 a,Ⅲ龄级为40~60 a,其他龄级以此类推[24]。统计分析后绘制群落的年龄结构图和种群的年龄结构图,以此判断群落的稳定性。
2 结果与分析
2.1 森林群落结构特征
从表1可见:乔木树种北坡最多的山体为10种,南坡最多的山体为6种,群落树种组成简单。研究区南北坡向上森林群落在结构数量上都有差异,北坡各山体间乔木层和草本层的物种数目相差较大,而南坡各山体间森林群落各层次在结构数量上差异不明显(东焦得布山草本层除外)。
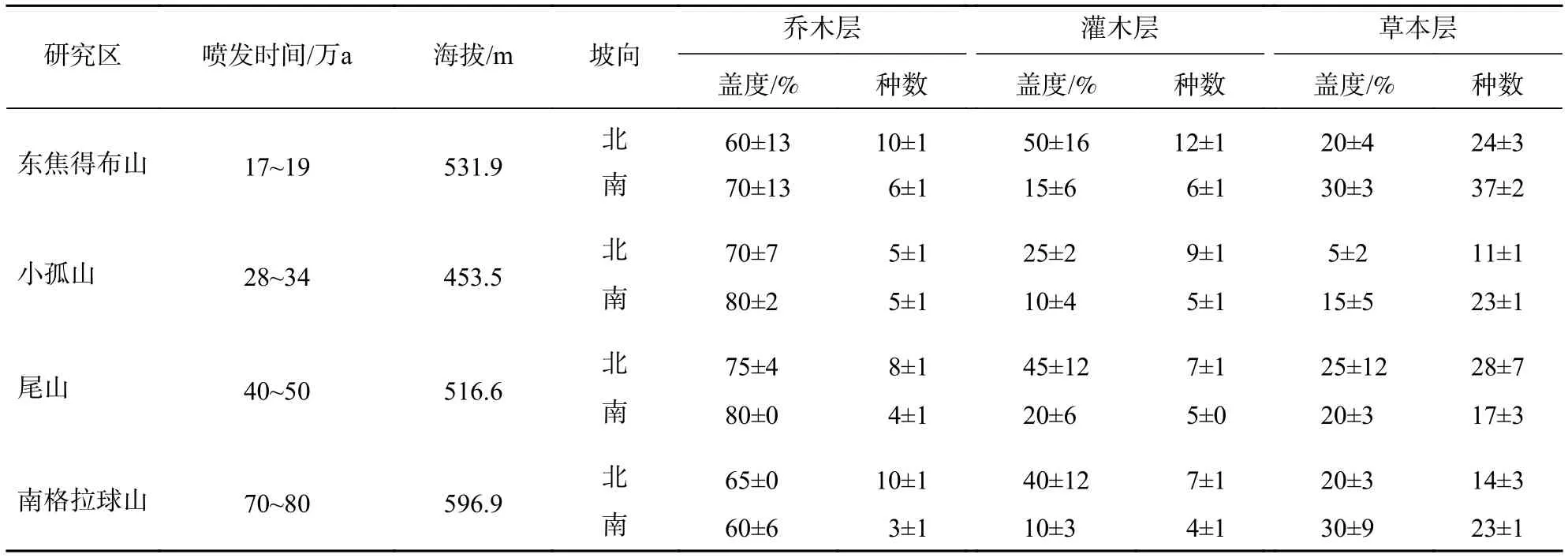
表1 五大连池火山森林群落的环境特征和数量特征Table 1 Characteristics of the quantitative and environment of forest communities in Wudalianchi Volcanoes
2.2 森林群落物种多样性特征
2.2.1 北坡森林群落物种多样性特征 从图1可见:4座火山北坡森林群落的各层次物种多样性指数有差异。Margalef指数和Shannon-Wiener指数从大到小依次为乔木层、草本层、灌木层,乔木层最大值分别为东焦得布山和尾山,最小值都为小孤山;灌木层最大值都为东焦得布山,最小值分别为南格拉球山和尾山;草本层最大值都为尾山,最小值都为南格拉球山。Simpson指数和Pielou指数从大到小依次为乔木层、灌木层、草本层,乔木层最大值都为尾山,最小值分别为小孤山和东焦得布山;灌木层最大值都为东焦得布山,最小值都为尾山;草本层最大值分别为尾山和东焦得布山,最小值都为南格拉球山。可见,4座火山北坡森林群落物种多样性主要受乔木层的影响;乔木层尾山的多样性指数、优势度指数和均匀度指数都最高,而小孤山的丰富度指数、多样性指数和优势度指数都最低;灌木层东焦得布山4种多样性测度指标都最大,而尾山多样性指数、优势度指数和均匀度都最小;草本层尾山物种的丰富度指数、多样性指数和优势度指数都最高,而南格拉球山4种多样性测度指标都最低。方差分析表明:4座火山北坡间,Margalef在乔木层是极显著差异(P<0.01),在草本层是显著差异(P<0.05);Shannon-Wiener指数在乔木层是显著差异外(P<0.05),其余群落内各层次的各种多样性指数均无显著差异(表2)。
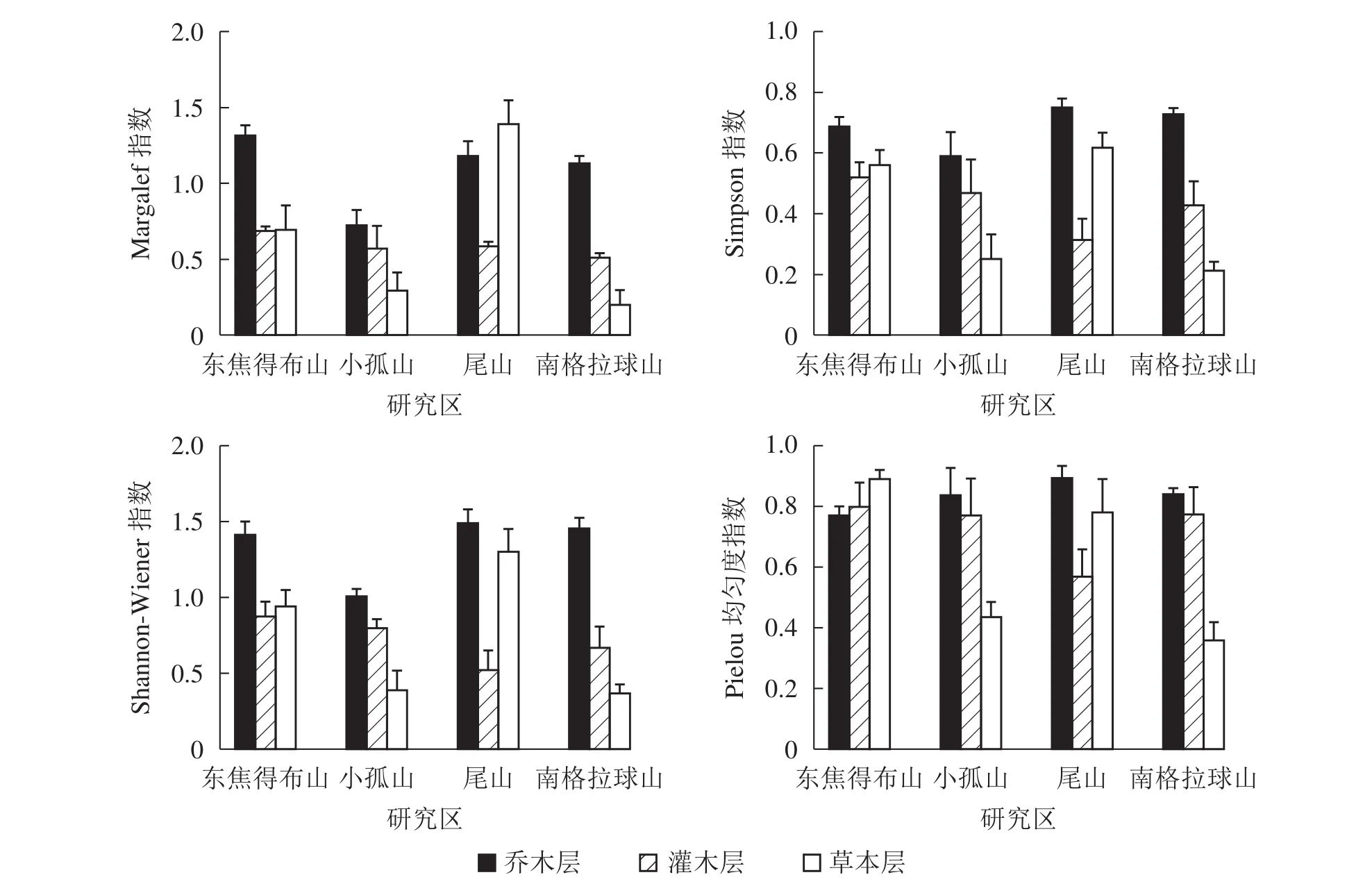
图1 五大连池火山北坡森林群落物种多样性特征Figure 1 Diversity index of forest communities in north slope of Wudalianchi Volcanoes
4座火山北坡间,群落内各层次的物种多样性指数呈不同变化。本研究用变异系数定量表示群落物种多样性指数空间变化程度的差异(表2)。草本层的各种多样性指数变化最大,乔木层的Simpson指数和Pielou指数变化最小。因此,乔木层在物种多样性指数的空间变化上比灌木层和草本层更稳定,草本层表现出最大的空间差异。
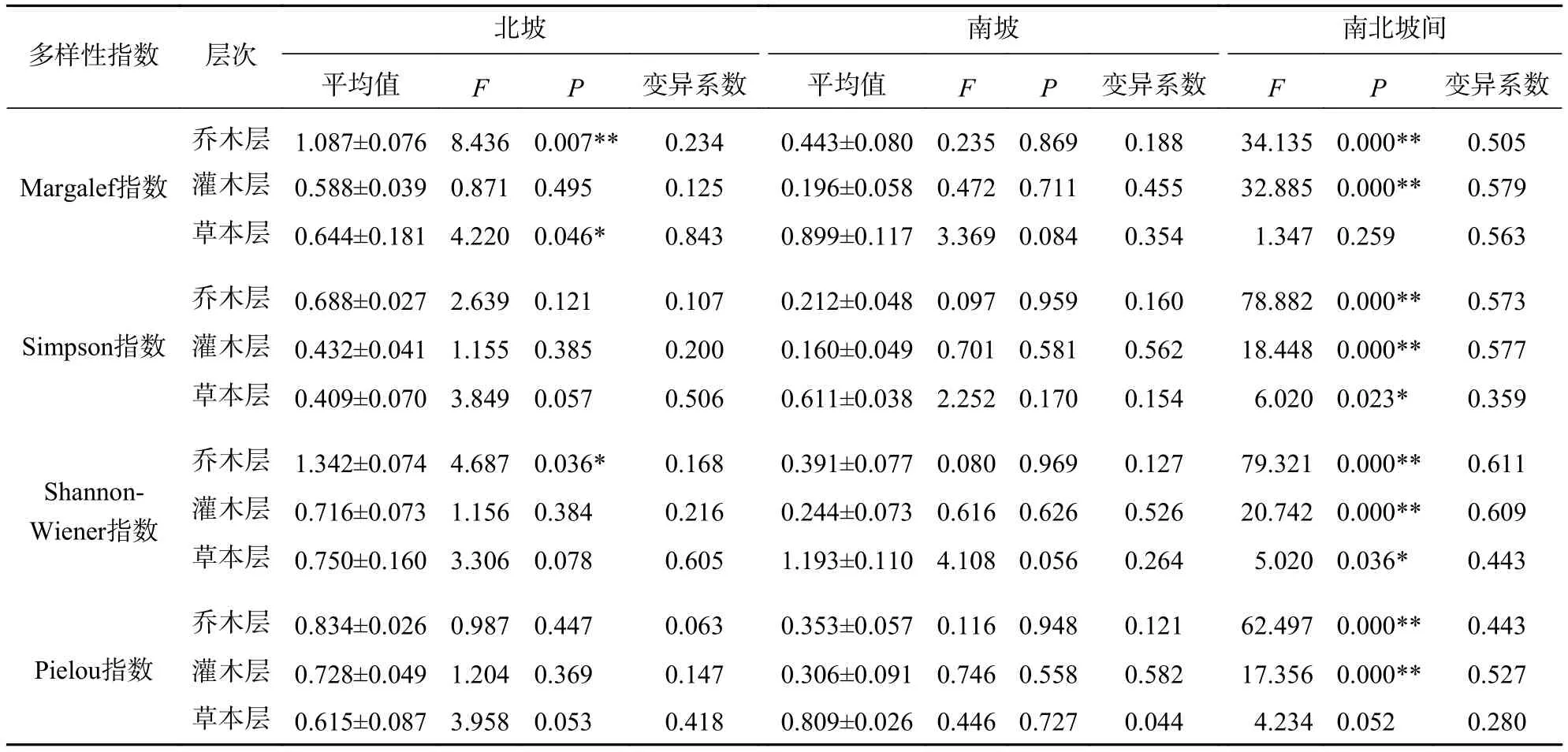
表2 五大连池火山森林群落多样性指数的方差分析和变异系数Table 2 One-way ANOVA and variation coefficient of the diversity index of forest communities in Wudalianchi Volcanoes
对4座火山北坡森林群落内各层次物种多样性指数进行相关分析(表3)表明:乔木层与草本层的各指数均呈正相关性,而乔木层与灌木层、灌木层与草本层之间仅丰富度指数呈正相关性,其他各指数间均呈负相关。说明4座火山北坡森林群落物种多样性主要受乔木层和草本层的影响。
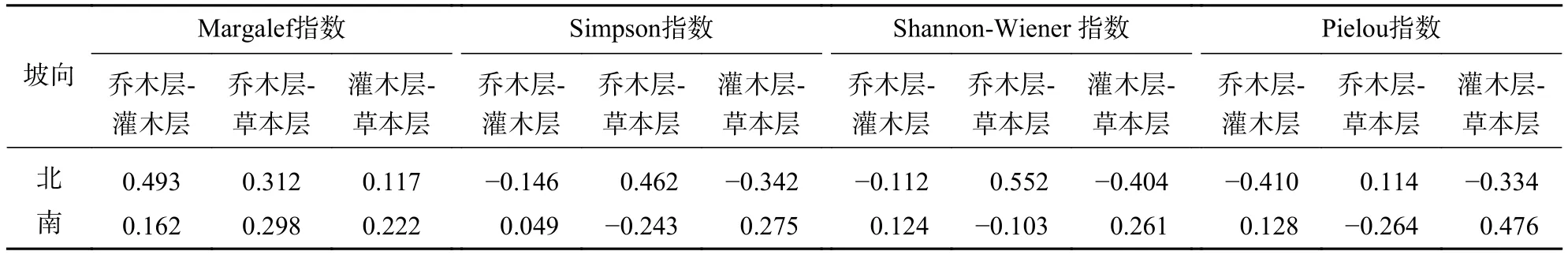
表3 五大连池火山森林群落层次间多样性指数的相关系数Table 3 Correlation coefficients of the diversity index between forest community layers in Wudalianchi Volcanoes
2.2.2 南坡森林群落物种多样性特征 从图2可见:4座火山南坡森林群落中的各层次物种多样性有差异。Margalef指数、Simpson指数、Shannon-Wiener指数和Pielou指数从大到小依次为草本层、乔木层、灌木层。乔木层Margalef指数、Simpson指数和Shannon-Wiener指数最大值都是东焦得布山,最小值都是南格拉球山;乔木层Pielou指数最大值为小孤山,最小值为东焦得布山。灌木层4种多样性指数最大值都为南格拉球山,最小值都为尾山;草本层Margalef指数、Simpson指数和Shannon-Wiener指数最大值都是东焦得布山,草本层Pielou指数最大值为小孤山,草本层4种多样性指数最小值都为南格拉球山。可见,4座火山南坡森林群落物种多样性主要受草本层和乔木层的影响。乔木层东焦得布山的丰富度指数、多样性指数和优势度指数都最大,而南格拉球山4种多样性指数都最小;灌木层南格拉球4种多样性指数都最大,而尾山4种指数都最小;草本层东焦得布山的丰富度指数、多样性指数和优势度指数都最大,而南格拉球山4种多样性指数都最小。
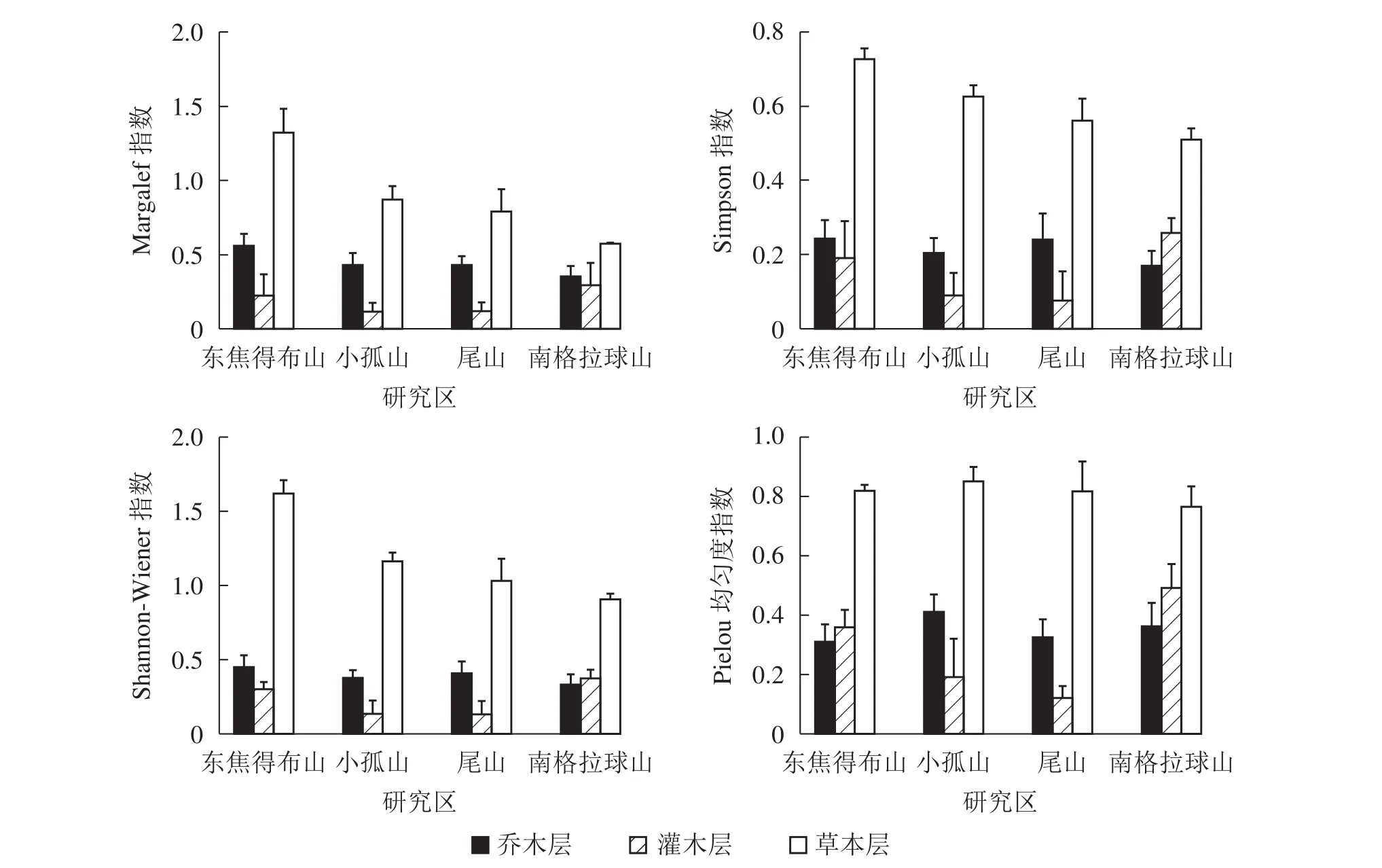
图2 五大连池火山南坡森林群落物种多样性特征Figure 2 Diversity index of forest communities in south slope of Wudalianchi Volcanoes
4座火山南坡间,群落内各层次的各种物种多样性指数均无显著差异(表2)。4座火山南坡间,灌木层的各种多样性指数变化最大,乔木层的Simpson指数和Shannon-Wiener指数总体上变化最小,草本层的均匀度指数变化最小。因此,乔木层和草本层在物种多样性指数的空间变化上表现出比灌木层更稳定,灌木层表现出最大的空间差异。
对4座火山南坡森林群落内各层次物种多样性指数进行相关分析(表3)表明:乔木层与灌木层、灌木层与草本层各指数均呈正相关,而乔木层与草本层之间仅丰富度指数呈正相关,其他指数均呈负相关。说明4座火山南坡森林群落物种丰富度主要受草本层的影响。
2.2.3 南北坡向间森林群落物种多样性特征比较 由表2可知:4种多样性指数都是乔木层和灌木层北坡高于南坡,草本层北坡低于南坡,说明北坡乔木层和灌木层的物种多样性指数高于南坡,而草本层低于南坡。同时,南北坡向间仅草本层的Margalef指数没有显著差异,其余物种多样性指数均呈极显著(P<0.01)或显著差异(P<0.05)。北坡乔木层和灌木层的Simpson指数和Pielou指数的变异系数都低于南坡,而北坡草本层物种多样性指数的变异系数均高于南坡。同时,南北坡向间物种多样性指数的变异系数都较大。
2.3 森林群落稳定性特征
2.3.1 北坡森林群落稳定性特征 从图3可见:4座火山北坡森林群落的年龄结构均为稳定型,群落表现稳定增长状态。小孤山Ⅱ~Ⅵ龄级(40~80 a)的乔木株数占个体总数的69.74%,且无Ⅰ龄级(0~20 a)个体,处于成熟树阶段,群落的稳定性较差,其余3座山Ⅱ~Ⅲ龄级(20~60 a)的乔木株数分别占总数的73.53%(东焦得布山)、56.56%(尾山)和75.90%(南格拉球山),处于中龄树阶段,群落的稳定性较好。
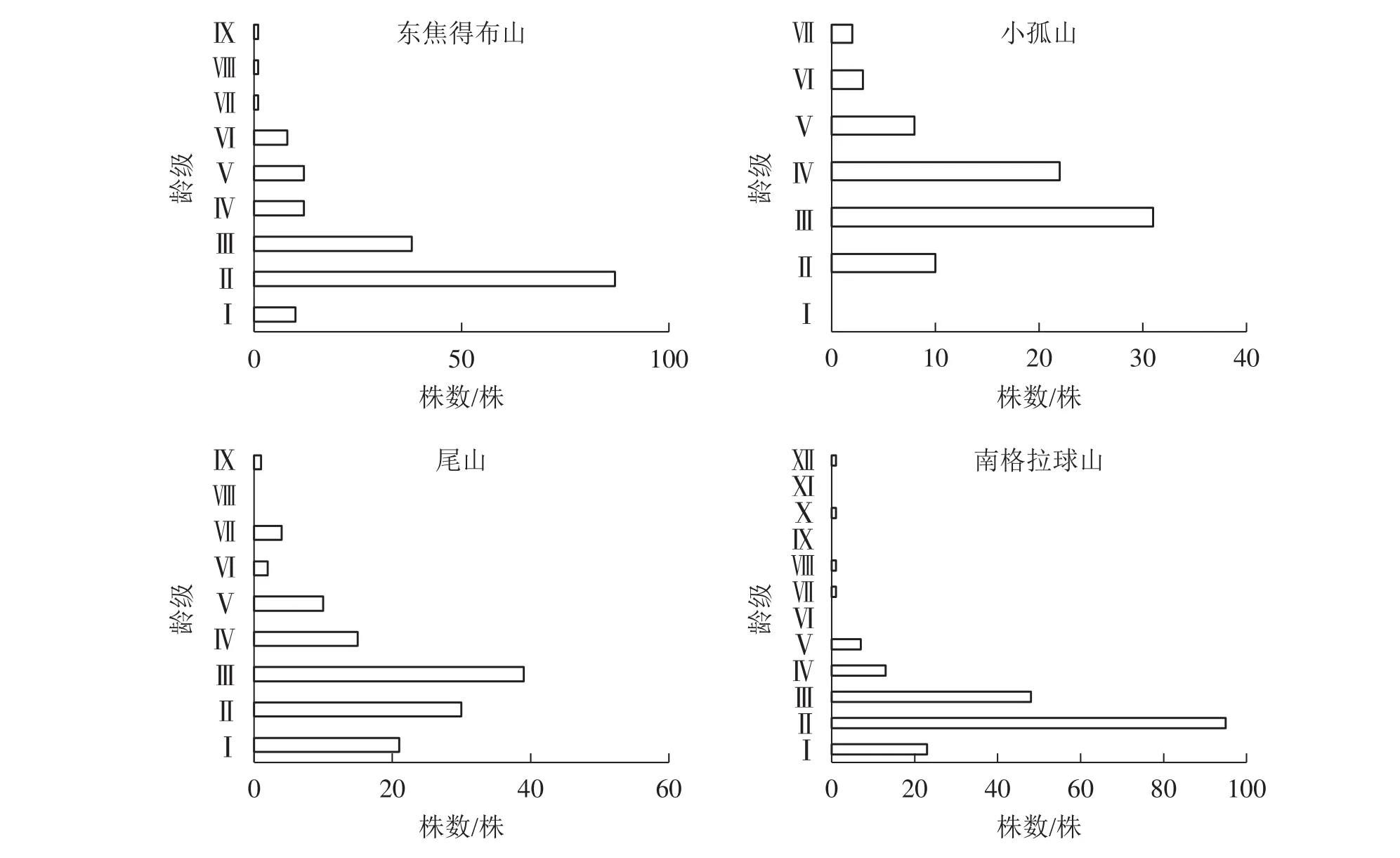
图3 五大连池火山北坡森林群落的年龄结构Figure 3 Age structure of forest communities in north slope of Wudalianchi Volcanoes
由表4可知:在北坡各山体的乔木层中,紫椴的重要值相对较高,其次为色木槭、山槐和黑桦,表明紫椴在北坡各山体的群落中重要性较大。为了更好地分析北坡群落的稳定情况,进一步对北坡乔木层中重要值最大的紫椴的年龄结构进行分析。
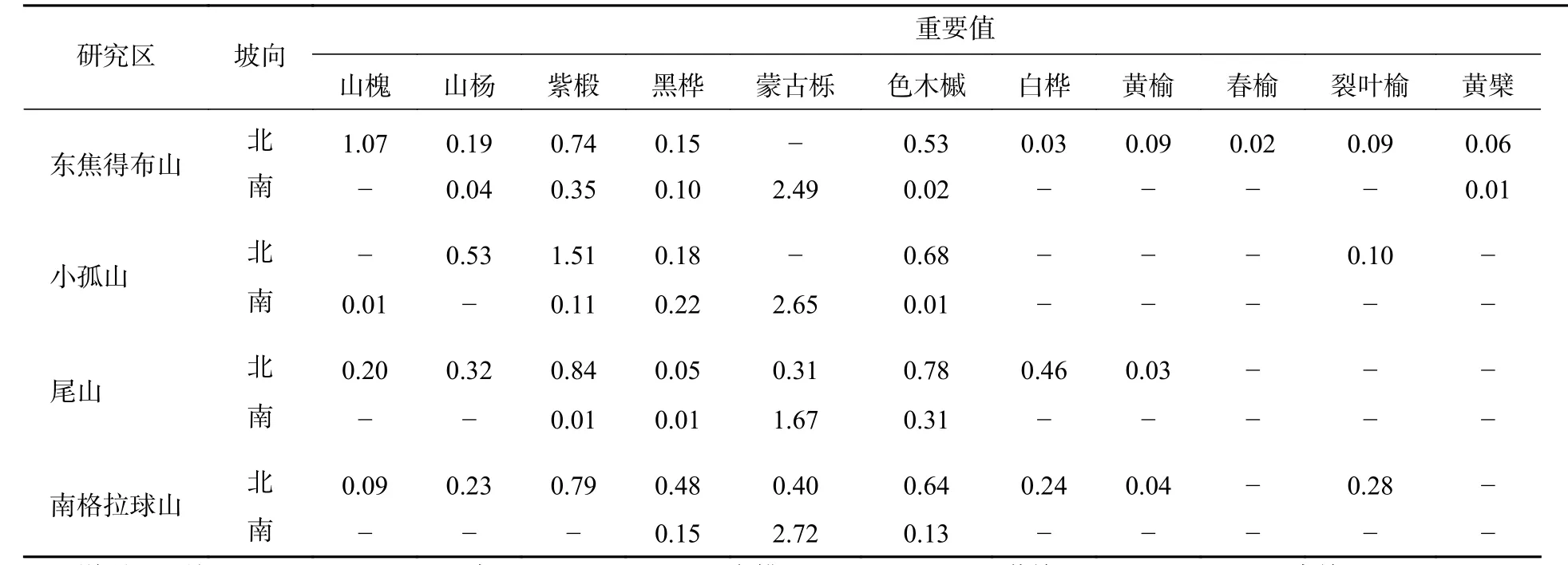
表4 五大连池火山森林群落乔木树种的重要值Table 4 Tree species with importance value of forest communities in Wudalianchi Volcanoes
由图4可知:4座火山的紫椴年龄结构均呈稳定型,Ⅱ~Ⅲ龄级个体数分别占总数的80.49%(东焦得布山)、56.76%(小孤山)、78.79%(尾山)和62.22%(南格拉球山),都处于中龄树阶段,群落的稳定性都较好,为稳定增长种群。群落的发展变化是以各个体的变化以及增减来实现的,年龄结构正是变化的依据,说明4座火山北坡森林群落处于稳定增长型状态。
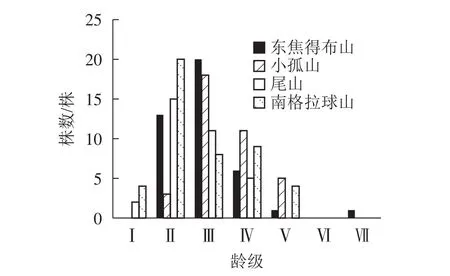
图4 五大连池火山北坡紫椴的年龄结构Figure 4 Age structure of T. amurensis in north slope of Wudalianchi Volcanoes
2.3.2 南坡森林群落稳定性特征 由图5可知:4座火山南坡森林群落的年龄结构均为稳定型,群落处于稳定状态。小孤山Ⅱ~Ⅵ龄级的乔木株数占总数的85.62%,处于成熟树阶段,群落的稳定性较差,其余3座山Ⅱ~Ⅲ龄级的乔木株数分别占总数的80.18%(东焦得布山)、59.16%(尾山)和80.41%(南格拉球山),都处于中龄树阶段,群落的稳定性较好。
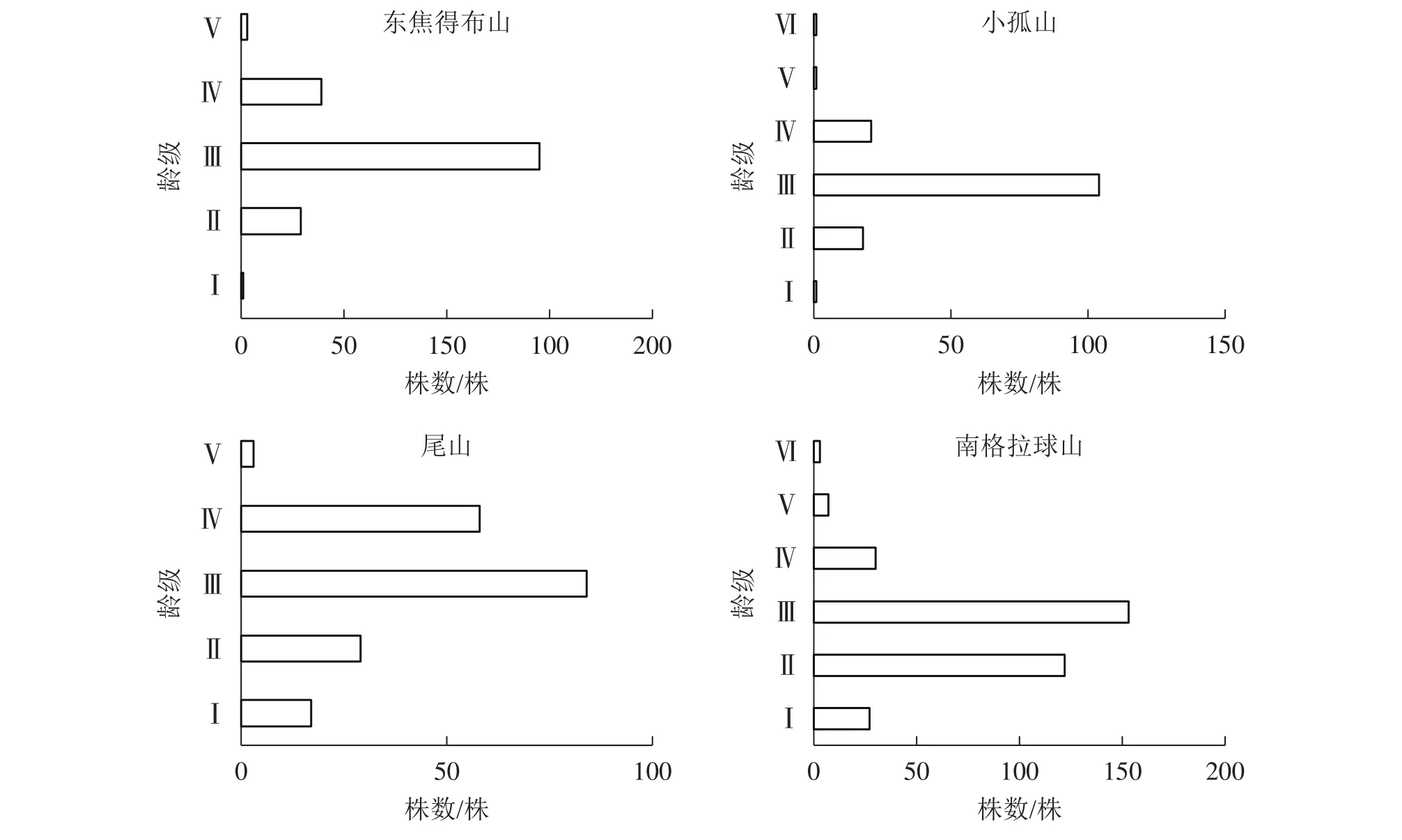
图5 五大连池火山南坡森林群落的年龄结构Figure 5 Age structure of forest communities in south slope of Wudalianchi Volcanoes
由表4可知:在南坡各山体的乔木层中蒙古栎的重要值较高,其次为紫椴和黑桦,表明蒙古栎在南坡各山体的群落中重要性较大。为了更好地分析南坡群落的稳定情况,进一步对南坡乔木层中重要值最大的蒙古栎的年龄结构进行分析。从图6可见:4座火山蒙古栎的年龄结构均呈稳定型,Ⅲ~Ⅵ龄级个体数分别占总株数的94.44%(东焦得布山)、86.82%(小孤山)、98.52%(尾山)和58.53%(南格拉球山),均处于成熟树阶段,群落的稳定性都较好,为稳定型种群,说明4座火山南坡森林群落处于稳定状态。
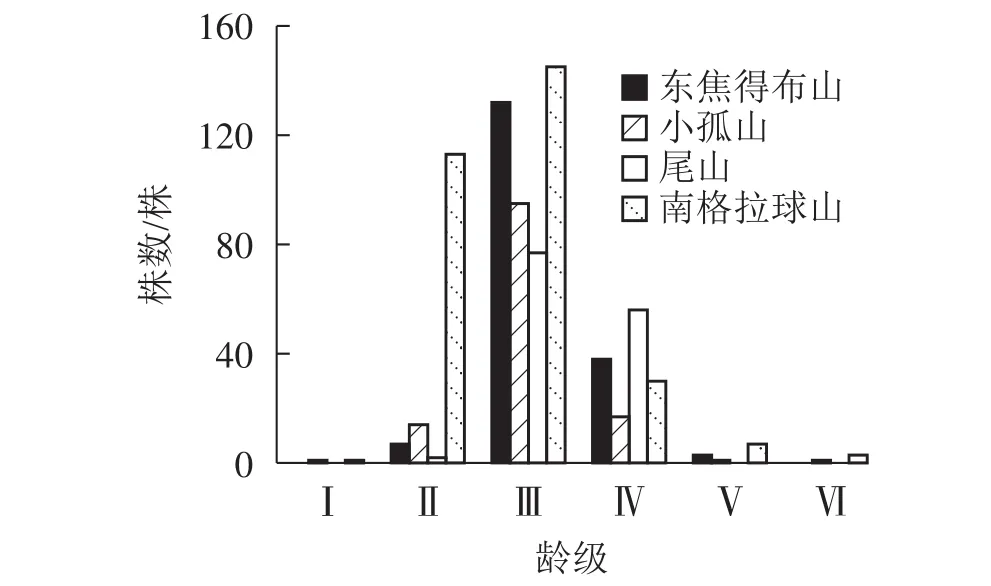
图6 五大连池火山南坡蒙古栎的年龄结构Figure 6 Age structure of Q. mongolica in north slope of Wudalianchi Volcanoes
2.3.3 南北坡向间森林群落物种稳定性特征比较 由图3~6可知:4座火山Ⅱ~Ⅲ龄级(20~60 a)的乔木株数分别占总数的比例均是北坡低于南坡,且北坡的龄级明显多于南坡。同时,北坡重要值最大的树种紫椴的Ⅱ~Ⅲ龄级个体数分别占总数的比例大,而南坡4座火山重要值最大的树种蒙古栎的Ⅲ~Ⅵ龄级个体数分别占总数的比例高达94.44%(东焦得布山)、86.82%(小孤山)、98.52%(尾山)、58.53%(南格拉球山),且蒙古栎的龄级少于紫椴。综上可知,北坡森林群落的稳定性强于南坡。
3 讨论
3.1 森林群落结构与物种多样性的关系
从群落结构的角度来研究生物群落的物种多样性是很有意义的,因为森林群落结构是群落中植物与植物之间、植物与环境之间相互关系的可见标志,也是群落其他特征的基础[25-26]。本研究各山体森林群落的乔木层和灌木层物种数北坡较南坡丰富,北坡物种多样性各指数也高于南坡,同时,森林群落结构较复杂的东焦得布山整体上物种多样性指数高。在各山体间南北坡上森林群落乔、灌、草3层物种丰富度和多样性变异都有差别。乔木层的Simpson指数变化都是最小,北坡上草本层的物种多样性各指数变化在各群落间表现出最大的差异,南坡上灌木层的物种多样性指数变化在各群落间表现出最大的差异,其原因在于物种多样性指数不仅受均匀度指数的影响,还受到物种丰富度的制约。各山体北坡森林群落间乔木层和草本层的物种数相差较大,故使群落间丰富度指数在乔木层和草本层分别是极显著差异和显著差异,Shannon-Wiener指数在乔木层是显著差异,且北坡上各森林群落内乔木层与草本层的物种多样性各指数均呈正相关。可见,北坡上乔木种类数量对草本物种有影响,而灌木层的物种数相差较小,导致物种多样性各指数没有显著差异;南坡森林群落间乔木、灌木、草本层物种数目相差不大,群落间各层次的物种多样性各指数均无显著差异,且南坡上各森林群落内乔木层与灌木层、灌木层与草本层的种物种多样性指数均呈正相关。
3.2 森林群落物种多样性与稳定性的关系
物种多样性和稳定性是植物群落的2个属性,它们之间的相互关系和相互影响已引起了国内外许多生态学者的关注[1,8-9,26]。均匀度是群落物种多样性研究中重要的概念[27]。以均匀度来考虑物种多样性与群落稳定性的关系时,群落的物种均匀度指数越高,群落的物种间相互差异越不显著,说明群落的稳定性越高,从演替动态的角度来看其稳定性就越高[1]。本研究森林群落物种多样性结果表明:乔木种群对群落具有支配作用,决定着群落的发展趋势,能够反映整个群落的物种多样性动态规律。因此,探知乔木层物种多样性与群落稳定性的问题,更有利于认知森林群落物种多样性与其稳定性之间的关系。高贤明等[1]在暖温带若干落叶阔叶林群落物种多样性及其与群落动态的关系研究发现:3个栎属Quercus林均匀度指数均较高,为0.56~0.76,是比较稳定的群落类型。本研究南北坡向各森林群落内乔木层的物种均匀度指数均较高,分别为0.31~0.41和0.77~0.89,是比较稳定的群落。森林群落的稳定程度和发展趋向,是受群落内外诸种生态学因素所决定。但是不管多方面的因素如何影响,影响的原因何等复杂,最终是以群落中各种群的变化来作为承受其结果的表达。因此,在群落的发展过程中,群落结构和相应种群结构变化可从年龄结构反映出来,相对稳定的森林群落应有相对稳定的种群结构,因而有相对稳定的年龄结构。不同稳定程度的森林群落的年龄结构图与种群的年龄结构图相近[22]。本研究南北坡森林群落的年龄结构都是稳定型,重要值高的蒙古栎和紫椴种群的年龄结构也都是稳定型,这说明南北坡各森林群落处于稳定状态。
3.3 坡向对森林群落多样性和稳定性的影响
在局部地区较小的尺度上,物种丰富度、多样性指数和均匀度指数也受到环境因素的影响。因为物理和生物因子的异质性发生在空间的各个尺度上,即使微生境如1株树或1束灌丛就可产生资源的异质性,从而影响其他生命体的分布(包括种类和数量)[28]。坡向影响了非生物资源分配,对地表接收的太阳辐射量能够产生较大的影响,进而使不同坡向的光、热、水、土等自然因素呈现较大的差异,营造局部小气候,从而使不同坡向的群落结构和群落物种多样性等产生相应的变化。在五大连池老期火山,南坡与北坡植物群落上层的主要生态因子光照和与之相关的水分和温度等生态因子存在一定的差异,耐干旱、瘠薄、喜光惯生长于阳坡的乔木和灌木种类少,耐阴湿惯生于北坡的乔木和灌木种类多,物种多样性增加,但同时北坡灌木种类多且盖度大导致草本植物可获得生长机会减少,致使北坡草本种类少且盖度小,物种多样性较南坡低。综上表明:北坡与南坡群落上层的光照、水分与温度等生态因子的差异导致北坡的乔木层和灌木层的物种多样性各指数均大于南坡,而草本层的物种多样性低于南坡,南北坡向间物种多样性各指数差异显著且变异系数都较大。稳定性与多样性具有更为复杂的关系,植物种的多样性并不能完全代表群落的稳定性,但却是群落稳定性的必要条件[29]。闫东锋等[30]在宝天曼栎属天然林物种多样性与稳定性研究中,通过群落物种多样性与稳定性相关机制的讨论,认为在森林生态系统中,物种多样性高可以导致较强稳定性,两者具有显著的正相关关系,并且发现最稳定的群落及不稳定的群落乔木层多样性指数的最大值分别为1.99和0.46。李凤英等[31]在凉水国家级自然保护区森林群落结构及物种多样性分析研究中发现:红松Pinus koraiensis-白桦Betula platyphylla森林群落乔木层多样性指数为2.08。本研究森林群落乔木层多样性最高值在北坡,为1.49,同时,北坡森林群落的年龄结构也较稳定,重要值显著高的紫椴种群的年龄结构也较稳定。综上所述,五大连池火山北坡森林群落多样性指数较高,且森林群落稳定性更好。
五大连池4座老期火山森林群落结构北坡较南坡丰富,北坡的乔木层和灌木层的物种多样性指数均大于南坡,而草本层的物种多样性低于南坡,南北坡向间物种多样性指数差异显著且变异系数都较大。北坡森林群落多样性指数也较南坡高,且森林群落稳定状态更好。同时,山体间森林群落结构较复杂的东焦得布山整体上物种多样性指数也较高。
