米槁天然林群落组成调查及分析
2021-04-17刘济明李丽霞
刘 欢,刘济明,李丽霞
(贵州大学 林学院,贵州 贵阳 550025)
植物群落物种多样性是衡量群落组成结构、物种丰富度、分布均匀程度的重要指标[1-2]。植物种间联结是种群在空间配置上的相互关联,能反映物种间的相互作用、物种与环境的耦合性及种群的空间分布格局[3],理论上有助于了解植物种间关系、正确认识植物群落结构特征、分析种群在水平格局上的动态变化[4-5]。因此,研究植物群落物种多样性及种间关联特征可以揭示群落的结构组成特征及其空间分布规律,同时为物种的生物学特性、种间相互关系和生境因子等因素综合作用下的种群个体水平空间配置和分布情况做出定量描述,为维持群落生态系统稳定性、资源合理利用、生物多样性保护提供理论依据[6-7]。
米槁(CinnamomummigaoH.W.Li)系樟科(Lauraceae)樟属常绿乔木,高可达20 m,主要分布于贵州、云南、广西三省交界的南北盘江、红水河流域,在贵州、广西两省交界区狭小区域,垂直分布范围为海拔300~1000 m[8]。其干燥成熟或近成熟果实为著名苗药大果木姜子[9]。作为重要的药用植物,其应用前景广阔、经济价值高且具有较高的生态效益,但由于近年来对米槁资源的不合理利用及其生境的不断恶化,导致野生资源面临枯竭,加之米槁人工繁殖较困难等原因,使其已处于濒危的状态,《中国生物多样性红色物种名录·高等植物卷》已将其列为近危种(Near threatened species,NT)[10]。前人对米槁的研究多集中于挥发油成分、光合生理及药物提取等方面[11-13]。除本课题组前期对米槁的种群特征进行过研究外,鲜有人对其群落结构、种间关系进行过深入探讨[14]。基于此,本研究通过对米槁天然林群落的物种多样性和种间联结进行了测定和分析,以期为米槁群落植物资源的保护和利用、生物多样性的维持和群落稳定性的研究提供科学依据。
1 研究地概况
研究地位于贵州省西南地区的罗甸县、贞丰县、册亨县、望谟县、荔波县。作为贵州省米槁野生资源的核心分布区,该区域非常适合于米槁的生长和繁殖,气候类型为典型的亚热带湿润季风气候[15],冬季低温少雨、夏季高温多雨,干湿季分明,年平均温度在16~20℃,雨量充沛,年均降雨量在1100~1335 mm,相对湿度78%~80%,无霜期长[14]。区域内地形起伏大,地貌复杂,以山地高原为主,成土母质多为砂页岩风化物,主要的土壤类型为红壤、黄红壤、黄壤、黄棕壤[16]。地带性植被为常绿阔叶林,植物类型丰富[17],米槁群落垂直结构分层明显,枫香(LiquidambarformosanaHance)、杉木[Cunninghamialanceolata(Lamb.)Hook]、青檀(PteroceltistatarinowiiMaxim)、油桐[Verniciafordii(Hemsl.)Airy Shaw]等构成乔木层主要物种;灌木层物种分布较乔木层均匀,主要物种有杜茎山[Maesajaponica(Thunb.)Moritzi]、地桃花(UrenalobataL.)、菝葜(SmilaxchinaL.)、胡颓子(ElaeagnuspungensThunb)、水麻(BoehmeriapendulifloraWedd);草本层以禾本科植物和蕨类植物为主,大多为一年生草本,代表植物为荩草[Arthraxonhispidus(Thunb.)Makino]、暗鳞鳞毛蕨[Dryopterisatrata(Wall.ex Kunze)Ching],林下还分布着乔木幼苗、幼树。
2 研究方法
2.1 样地设置与调查
在对贵州省米槁野生资源分布情况进行实地调查的基础上,选择米槁分布较集中,群落结构完整,地形平坦,且人为干扰较少的天然林进行野外群落调查[14]。采用常规的群落调查方法[18],选择8个具有代表性的样点,设置9个不连续的样地,根据地形起伏的差异和地势条件将样地设置为不同的规格,分别为20 m×50 m(6个)、20 m×30 m(1个)、20 m×20 m(1个)、30 m×30 m(1个),总面积为7900 m2,记录样地的海拔、坡度、坡向、郁闭度等生境指标。将每一个样地看作一个大样方,在9个样地里共调查79个10 m×10 m的乔木小样方、对样方内的乔木树种进行测量其高度、胸径、冠幅等,在每个乔木小样方内划分出一个5 m×5 m的灌木样方和1 m×1 m的草本样方,记录其植物名称、数量高度等指标。
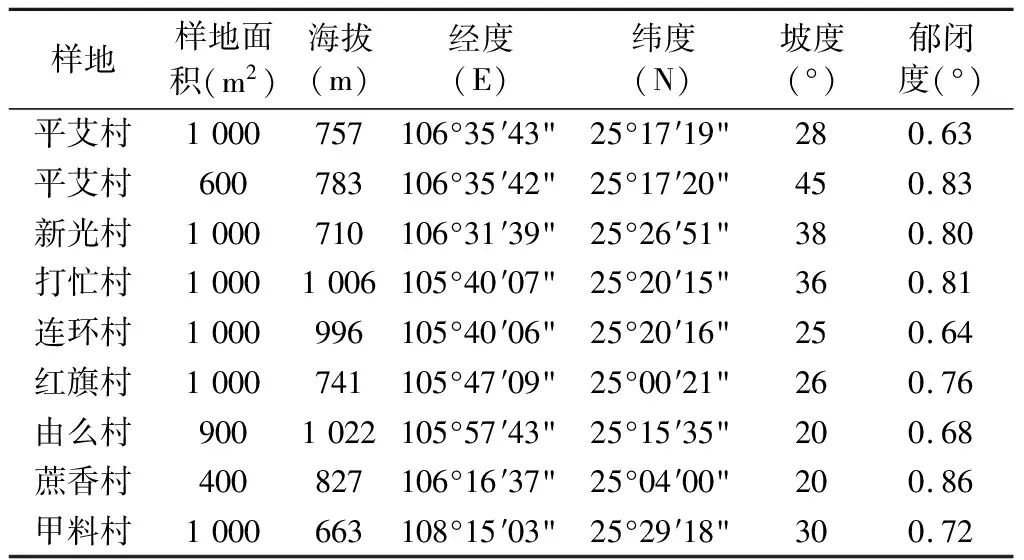
表1 样地基本信息
2.2 多样性计算
根据野外调查数据,将米槁群落组成结构进行分类:乔木层、灌木层(包括胸径≤5 cm、高度≤ 3 m的木本植物和灌木)、草本层[19]。以物种的重要值为测度指标,采用α多样性指数综合评估群落物种多样性水平,包括Margalef丰富度指数,Shannon-Wiener多样性指数、Simposon优势度指数和Pielou均匀度指数。
(1)重要值(Important value, IV)[20]:
乔木层:IV=(相对密度+相对频度+相对显度著)/3
灌木层:IV=(相对频度+相对密度)/2
草本层:IV=(相对密度+相对频度+相对盖度)/3
(2)多样性指数[21-23]:
Margalef丰富度指数(E):
E=(S-1)/lnN
Shannon-Wiener多样性指数(H):
Simposon优势度指数(D):
Pielou均匀度指数(J):
J=H/lnS
式中S为样地内出现的物种数,ni为第i个物种的重要值,Pi为第i个物种的相对重要值,N为群落中所有植物的重要值之和。
2.3 种间联结计算
选取米槁天然林群落中重要值排名靠前的18种主要木本植物进行种间联结性分析。根据原始数据矩阵,将物种间所有种对的定性数据列入2×2列联表,计算a、b、c、d的值[24]。由于取样的非连续性,用Yates修正过的χ2统计量对种间联结性进行定性研究。
式中:N为取样总数,a为2个物种都存在的样方数,b、c分别为2个物种独自存在的样方数,d为2个物种都不存在的样方数,ad>bc,表示种间正联结,ad
用联结系数AC和种间共同出现率PC来表征种间联结强度[26]:
式中AC值越趋近1,表明种间的正联结性越强,而AC值越趋于-1,表明种间的负联结性越强,当AC值为0时,种间完全独立;同理,PC值越趋近于100%,则种对正联结越强[27]。
3 结果与分析
3.1 群落物种组成
物种多样性可以综合全面地反映森林群落的发展阶段和稳定程度,体现出物种与其周围环境间的相互关系。因此,进行群落物种多样性的研究可以为植物资源的合理保护与科学利用提供参考[28]。在9个样地中,共调查到139种维管束植物(除去灌木层中与乔木层中重复的物种)隶属66科113个属。其中木本植物有92种,占总数的66.19%,草本植物有47种,占总数的33.81%。从植物种类来看,5种及以上的科有8个,这8个科包含了群落中39.57%的物种,而且群落优势种大多来自于这些科,如樟科的枫香、米槁,禾本科的荩草。2~4种之间的科有20个,占总数的30.30%,其它科的植物大多以两属两种的形式分布在群落中,有38科的植物以单属单种的形式出现,说明群落中植物种类的多样性高,分布不均匀。
从群落组成结构的角度来探讨物种多样性对划分群落功能种群具有重要作用。米槁群落的垂直分层结构明显,主要划分为乔木层、灌木层和草本层。其中乔木层有39个物种,占总数的24.84%,以枫香、米槁、杉木、油桐为主;灌木层有71个物种,占总数的45.22%,以杜茎山、地桃花、油茶、菝葜为主;草本层有47个物种,占总数的29.94%,主要为荩草、暗鳞鳞毛蕨、铁芒萁、凤尾蕨。
3.2 重要值分析
对米槁天然林群落不同层次排名前10的物种进行重要值分析,结果表明,乔木层与草本层的重要值之和都达到了5.000以上。在乔木层中,枫香(1.604)和米槁(1.261)的重要值最大,远远超过其它种,在群落中的分布范围广泛,为群落中的优势种。灌木层中,杜茎山(0.723)的重要值最大,其它种主要集中在0.200~0.500之间,胡颓子(0.147)的重要值最小,在群落中零星分布。草本层中,荩草、暗鳞鳞毛蕨、凤尾蕨、铁芒箕、贯众和紫茎泽兰的重要值都在0.500以上,卷柏、苍耳、五叶地锦的重要值偏低,在群落中分布稀疏。

表3 米槁群落主要物种重要值分析
3.3 多样性分析
如表4所示,物种丰富度、优势度和个体分配的均匀程度构成了物种多样性,它能够反映群落结构和生态系统的复杂性。米槁群落内物种多样性表现为灌木层>乔木层>草本层,各样方中灌木层的物种数量多、分布较乔木层和草本层均匀,各项指标值均最高。草本层的物种多于乔木层,但其Simposon-Wiener多样性指数和Pielou均匀度小于乔木层,主要是因为草本层中荩草、暗鳞鳞毛蕨、铁芒萁、凤尾蕨的种群数量多、重要值大,使群落物种分布的均匀程度降低。

表4 米槁群落物种多样性指数
3.4 主要木本植物种间联结性分析
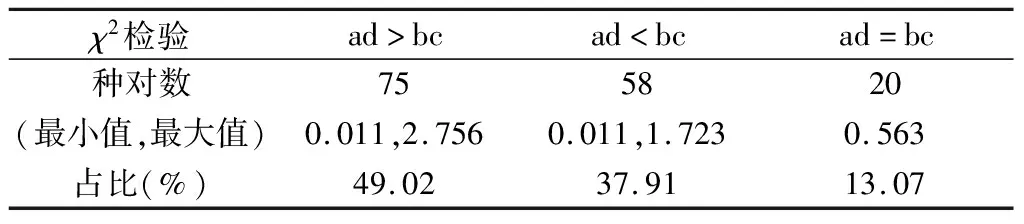
表5 米槁群落主要物种的总体联结性
表6反映了群落内153个种对间的相互关联程度(AC),有75个种对呈正联结,占总对数的49.02%,58个种对呈负相关,占总对数的37.91%,还有20个种对间的AC值为0,种间完全独立。从PC值结果来看,PC值大于60%的仅有5对,占总数的3.27%。说明群落物种间的生态习性和生境要求不一致。
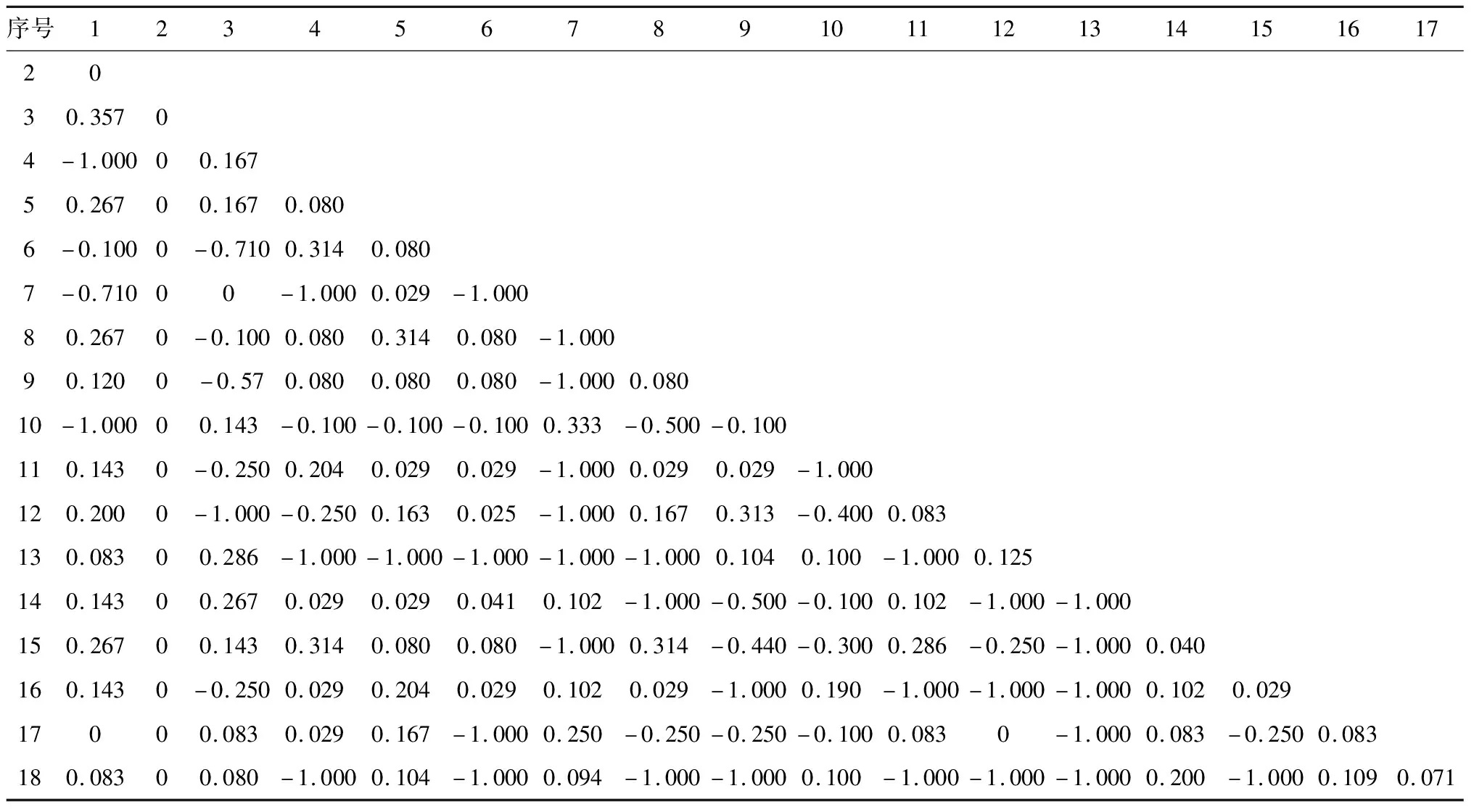
表6 联结系数AC矩阵
4 讨 论
调查共计维管束植物139种,隶属66科113个属。生活型最多的是灌木,以紫金牛科和茶科的物种为主;木本植物中的蔷薇科、樟科占据优势地位,这两个科的物种对调查样地的适应性极强。此外,禾本科、壳斗科、菊科、豆科在群落中的种属都相对较多,与环境有较高的耦合性。对生境的适应性也强。从群落垂直结构来看,米槁天然林群落垂直分层明显,不同层次的物种丰富度指数、多样性指数、优势度指数和均匀度指数总体变化趋势一致,灌木层的物种多样性大于乔木层和草本层。该群落物种多样性格局形成的原因可能是,乔木层和草本层的物种组成相对固定,且局部优势突出,主要优势种在群落中占据统治地位,而灌木层的植物种类除了灌木树种外,还包括一部分乔木树种的幼苗、幼树,使得灌木层物种在群落中分布较均匀,各项综合指标较高[28]。
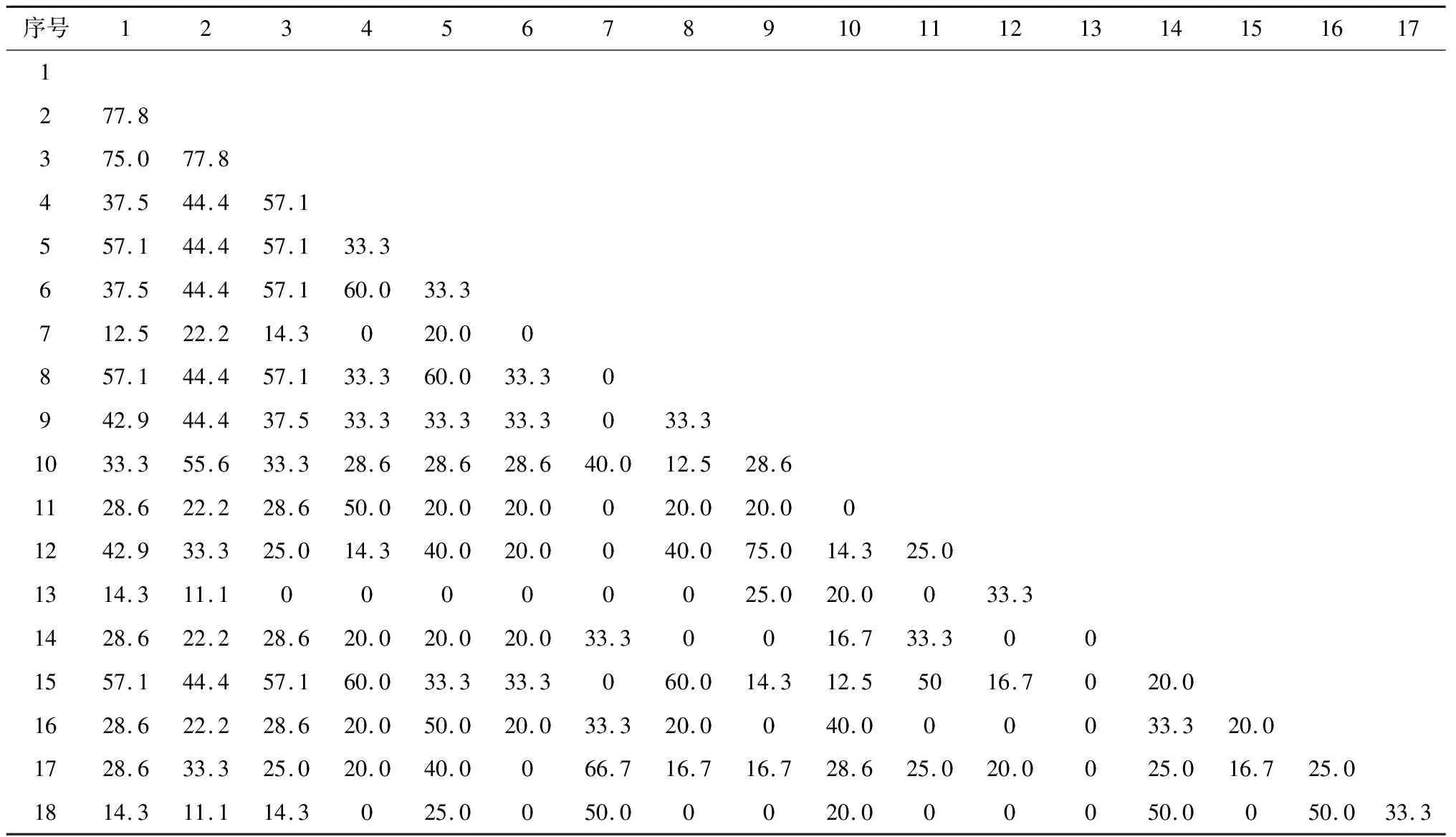
表7 米槁群落主要物种共同出现百分率矩阵
本研究以2×2联列表为基础,使用χ2检验、联结系数AC与物种共同出现百分率PC,测定了群落主要物种间的联结性,进而分析群落间的物种组成与分配情况。从测定结果来看样地内乔木树种间的种间联结性并不显著,基本上趋于独立出现,说明乔木层树种在种间关系上比较松散。这可能是因为群落在长期的演替过程中,由于物种对资源的竞争关系,使群落组成结构维持在一个稳定的状态,物种都占据一个有利于自己的生态位,所以表现出较大的独立性。同时,各种对间的独立性强,正负联结种对数差异不大,表明在长期的演化过程中各种群相互竞争并和谐共生于群落中,这与刘春生、刘金福等人的研究结果一致[29-30]。同时,已有多个研究表明,植物群落内呈现正联结的物种对生境具有相近的适应性,两个物种间的正联结强度越高,表明它们之间对生境资源的利用特征越相似,生态位重叠越高;负联结表明两个物种间对资源的利用存在明显的差异或者两者之间存在激烈的竞争关系[28,31-32]。本研究结果也证明了这一点,群落主要物种中枫香和杜茎山、椤木石楠和杉木、菝葜和青檀、地桃花和板栗均表现出较强的正联结,它们的生态位重叠值也较大,对生境的适应性较强;杜茎山和椤木石楠、油茶以及地桃花和菝葜等种对间的负联结偏向性较大,对生境资源的利用相似性低,生态位重叠值小,这可能是由它们对环境要求的差异性或相离的生态位决定的。此外,以共同出现百分率PC来评估特殊种对米槁天然林群落物种多样性的影响时,重要值偏高(如紫茎泽兰,0.517),但在群落中的出现频率小、与其他物种间的联结性弱的物种对群落组成结构的影响较小。
上述研究结果,初步揭示了米槁天然林的群落组成特征和群落物种间的相互关系。米槁作为群落优势种,在群落中占据着绝对优势地位,对资源的利用能力强,但群落内联结程度弱的种对占总对数的比例较大,群落的稳定性差,故以后在对米槁天然林群落的经营和管理中,应该合理控制群落密度,剔除一些负联结性较强的伴生物种,选取一些正联结性较强的树种来缓解种间竞争,使群落达到一个相对稳定的状态,促进物种的共生与发展。
目前在贵州省内的米槁分布范围较窄,现存林分基本为过熟林,小苗和幼树较少,种群的年龄结构属于衰退型,不利于林下幼苗的生长发育,因此,要切实保护这一濒危种群,应加速开展米槁的生理学、生物学、分子遗传学等研究,并通过人工繁育扩大米槁的种群数量。
