敦煌市葡萄园土壤呼吸及其组分变化
2021-04-16朱高峰秦文华陈惠玲
张 扬, 朱高峰, 赵 楠, 秦文华, 陈惠玲
(兰州大学 资源环境学院, 西部环境教育部重点实验室, 甘肃兰州 730000)
土壤呼吸作为全球碳循环和气候变化的重要影响因子之一[1],它是土壤有机碳的主要输出途径和大气CO2的重要来源[2],同时也是陆地生态系统碳循环的第二大通量[3]。土壤呼吸主要包含土壤根呼吸(即自养呼吸)、土壤微生物呼吸(即异养呼吸)、含碳物质的化学氧化和土壤动物呼吸,后两者作用较微弱,通常忽略不计[4-5]。因此,许多研究中土壤呼吸被区分为两大组分:自养呼吸和异养呼吸[6-8]。近年来,大量学者通过野外观测、数据整合分析和模型模拟等方法对土壤呼吸进行了研究,主要集中在土壤呼吸速率变化及其影响因素方面[9-10],内容已涉及森林、草原、农田等多种生态系统[11-13]。其中农田受人类生产活动扰动最为强烈,其土壤呼吸变化及其对环境因子的响应过程更为复杂。因此,较多学者已开展各类农田土壤呼吸研究,且多集中在小麦、玉米等[14-15]农田中,而针对中国西北干旱区广泛种植的葡萄园关注仍相对较少,中国西北干旱区生态环境脆弱,绿洲农田具有独特的温度和水分条件,因此深入探究其土壤呼吸与土壤温湿度的关系显得尤为重要。
本文以敦煌南湖绿洲葡萄园为研究对象,于2019年5月设计试验,6—12月连续观测其土壤呼吸和环境因子变化,对观测期间葡萄园的土壤呼吸及其组分变化进行分析,探讨土壤呼吸与土壤温度、土壤含水率的关系,旨在明晰该区特色经济作物(葡萄)的土壤呼吸变化的内在机制,研究结果可为西北干旱区的土壤碳排放的估算以及特色农业发展提供一定的参考。
1 研究区概况
研究区位于甘肃省敦煌市西南方向约70 km的南湖绿洲,东经94°06′—94°09′,北纬39°51°—39°55′之间,海拔约1 100~1 297 m[16-17]。该区光照充足,多年平均日照时数约为3 115~3 247 h[18]。同时,党河水库为其提供了稳定的灌溉水源,且土壤类型主要为沙壤土。上述光、热、水和土壤等资源,为葡萄的广泛种植提供了适宜的条件。
该区葡萄品种主要是无核白葡萄,熟制为一年一熟。本研究选取一个典型的无核白葡萄样地(52 m×43 m),其土壤有机碳、全氮含量分别为13.3,0.9 g/kg,容重为1.22 g/cm3,pH值约为8.3[19];葡萄种植行向为东西向,架口向南,行间距约3 m,株距1 m。
葡萄的生长季在4—9月间,其中4月底出土,5月初萌芽,5月底开花,6—7月中旬结果,7月底至8月成熟,9月采摘,10月初埋土冬藏。为保证葡萄树的正常生长,分别在出土期、萌芽期、开花期、两次结果高峰期和埋土前期通过沟渠进行漫灌,每次灌水量约为145 mm[16],其中8月底果实糖分积累需水量减少。同时每次灌水前伴随着施肥过程,4月底施加约375 kg/hm2氮肥(以尿素为主),5—8各月灌前则追施复合肥约375 kg/hm2。
2 材料与方法
2.1 试验设计
2019年5月,选择自动气象站附近的两行葡萄树,在距离葡萄主干约0.3~0.9 m处取6个0.6 m×0.6 m的样方。其中,2个不做任何处理以测定土壤呼吸,另外2个用去根埋网法处理,还有2个用挖沟隔板法处理,这两种方法的原理是排除葡萄根系呼吸,即测定的是异养呼吸。去根埋网法具体处理如下:①将2个样方深挖1 m,土体分层放置并记录深度:0—20,20—40,40—60,60—80,80—100 cm;②在样方四周,放置0.15 mm的尼龙网阻隔根系;③筛选出土内细根,把土按原位置回填。而挖沟隔板法则在2个样方的四周挖壕沟,放置薄板以阻隔根系。上述6个样方处理后将PVC环插入样方内的土壤中,出露约2 cm。为减少土壤扰动和残留根系的影响,于6月待土壤结构稳定后使用LI-8 100 A及其配套的LI-8 150多路器连接6个土壤呼吸测量气室开始连续同步的测定土壤呼吸。
2.2 土壤呼吸及环境因子测定
土壤呼吸速率采用定点连续测定,测定时间为2019年6月21日至12月9日,每日进行12次测定,测定间隔为2 h。由于仪器损坏和灌溉原因导致9月只有2 d土壤呼吸数据,其他月份个别日数据缺失。本研究以9月30日为界分为两段,6—9月属于生长季,而10—12月属于非生长季。其中,异养呼吸为两种处理方法(去根埋网法和挖沟隔板法)的平均值,自养呼吸则由土壤呼吸和上述异养呼吸的差值得到。
气象数据〔空气温度(Ta)、降水量〕和5 cm处土壤温湿度(Ts5,θ)的测定时间为4月24日至12月9日,间隔为30 min。其中Ta和降水量分别采用温度传感器(HMP60,Vaisala,Finland)和翻斗式雨量筒(TE525,Texas Electronics,USA)观测,而土壤体积含水率和土壤温度则采用埋在地下5 cm深度处的土壤温湿度传感器(5TE,Decagon,USA)长期监测样地的土壤温湿度变化。
研究还结合了各个土壤呼吸测量气室上连接的土壤温度探头测定的5 cm土壤温度来分析小时尺度上的土壤呼吸对土壤温度的响应,其优势是可以获得土壤呼吸测量样方处的土壤温度;但由于其较容易损坏,因此在日尺度上则采用了样地中间的小型气象站测得的土壤温湿度数据。
2.3 数据分析
(1) 5 cm土壤温度与土壤呼吸、异养呼吸的关系采用指数模型拟合[20],式(1)所示:
(1)
式中:R为呼吸速率〔μmol/(m2·s)〕;Ts5为5 cm处土壤温度(℃);α,β分别为拟合参数。
(2) 土壤呼吸的温度敏感性计算如式(2)所示[20]:
Q10=e10β
(2)
式中:Q10为土壤呼吸的温度敏感性;β为式(1)中拟合参数。
(3) 为了分析观测期内不同土壤含水率对于土壤呼吸及其组分的影响,我们选取时间跨度小且较连续的时间段,尽可能的减小其他环境因子和植物生理要素对于土壤呼吸的影响,筛选了生长季的3个阶段(6月21日至7月6日、7月18日至8月7日和8月8日至8月29日)和非生长季的1个阶段(10月25日至12月9日),依次对应微弱降水后、年最大降水后、大水漫灌后和无灌溉无降水之后。生长季3种水分条件下,对土壤呼吸及其组分与土壤含水率的关系采用二次函数进行描述[21],见式(3);而非生长季采用指数模型拟合,见式(4):
R=αθ2+bθ+c
(3)
R=ea+bθ
(4)
式中:R为呼吸速率〔μmol/(m2·s)〕;θ为5 cm处土壤含水率(%);a,b,c分别为拟合系数。
上述数据处理与结果绘图均采用Origin 2017软件进行处理。
3 结果与分析
3.1 环境因子变化
温度和水分是影响土壤呼吸的主要因素,观测期内Ta和Ts5的逐日变化见图1a。在生长季的6—9月份,平均Ta高于Ts5,分别为20.08,19.58 ℃,在8月Ta和Ts5均达到最大,分别为22.36,20.39 ℃;而非生长季的10—12月份,Ta和Ts5逐渐下降,同时由于土壤和空气对热的传导性不同,导致Ts5的降低速率低于Ta,因而Ta值低于Ts5值。
θ的变化主要受灌溉的强烈影响(图1b),其6次大的突变分别是由6次灌溉事件所导致。而第198 d(7月17日)发生1次小突变,主要是因为该天降水量达到9.14 mm,为该区年内最大日降水。另外每次灌溉或降水后,θ的降低速率随时间逐渐变小。非生长季的10—12月,θ在灌溉后降低速率先快速,然后缓慢(298~330 d),再快速(第330 d后)。先快后慢主要是因为初期θ较高,下渗快,蒸散发强,θ快速下降,之后葡萄植株埋至地下,随着辐射减弱,水分蒸发变弱,同时根系吸水减少,仅少量吸水维持生命活动。而在第330 d又快速降低,主要与冻后聚熵有关,即此时Ts5始终接近0 ℃但位于0 ℃以上,所以5 cm处土壤没有冻结,而Ta已经低于0 ℃,因此表层土壤可能产生冻结现象,水汽压降低,而冻土以下土层(5 cm土层)的水汽压较高,于是5 cm处水汽不断向冻层集聚、冻结,使得冻层不断加厚,表层含水量有所增加,而θ则迅速降低。
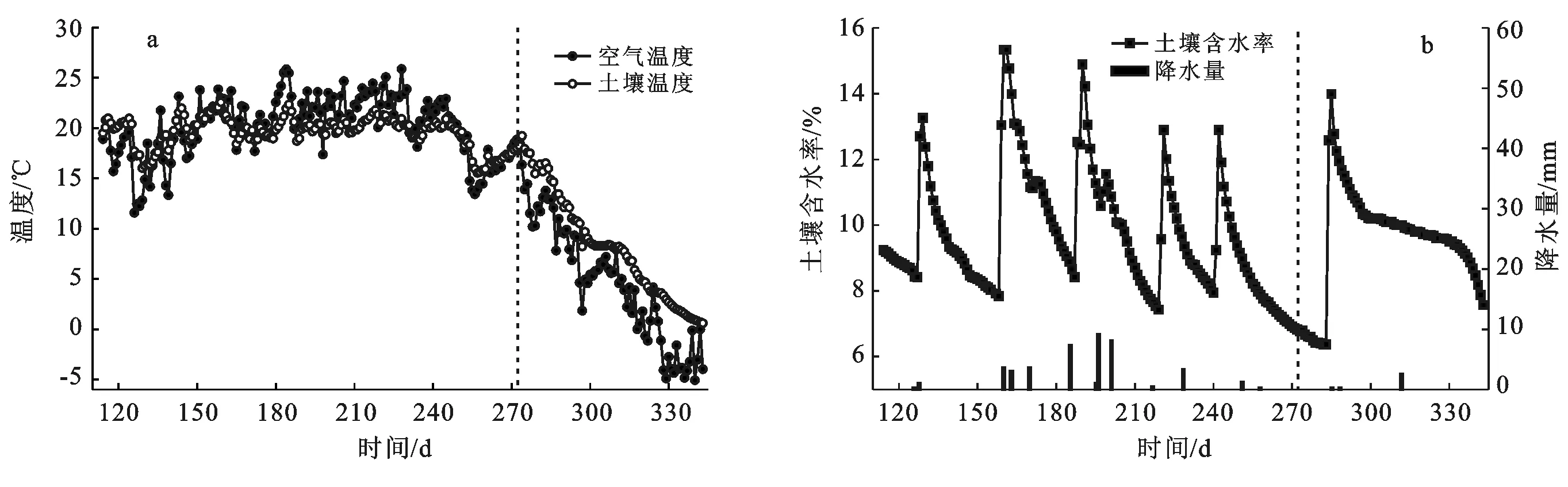
图1 环境因子的逐日变化(生长季和非生长季以虚线为界)
3.2 土壤呼吸及其组分贡献率变化
(1) 土壤呼吸速率变化。观测期内,葡萄园的土壤呼吸速率逐日变化见图2a,7月3日达到最大,为6.31 μmol/(m2·s),此时处于生长旺盛的结果期。在生长季的6—9月,土壤呼吸速率波动变化明显,其中7月和8月土壤呼吸速率各有1次突降,突降日期对应着农田灌水时间,表明每次农田灌溉积水后θ突增,此时会短暂抑制土壤呼吸,之后随着θ逐渐减小,土壤呼吸速率逐渐恢复。而非生长季的10—12月,土壤呼吸逐渐减小,这主要与土壤温度的逐渐降低密切相关。
(2) 异养和自养呼吸的贡献率变化。异养和自养呼吸的贡献率变化如图2b所示,在生长季的6—9月,异养和自养组分的贡献率存在明显波动,但异养组分对于土壤呼吸的贡献率普遍高于自养组分,且二者的平均贡献率分别为64.8%和35.2%。对于整个生长季,按照二者对土壤呼吸的贡献率大小可分为3个阶段:第一阶段是第172—187 d,该阶段异养和自养呼吸同步增长,因此二者的贡献率稳定,平均贡献率分别为53.6%和46.4%。第二阶段是第197—219 d,年最大降水使得土壤呼吸、异养和自养呼吸三者均受到抑制,随着土壤含水率减少,一方面,自养呼吸未能恢复到原水平,主要是该区多次灌溉使得土壤板结,葡萄根系多分布在60 cm乃至更深,植物根系呼吸不够顺畅所致;另一方面异养呼吸量却能基本恢复到第一阶段的水平,主要与该区长期大水漫灌形成的地下少氧环境使得该区微生物以兼性厌氧和厌氧等为主。综上两方面,导致自养呼吸贡献率极低。第三阶段为221—273 d,因第220 d进行了灌水,导致第221 d的异养呼吸值减小,自养呼吸值增大,因此自养呼吸贡献比突增,之后随着含水率减小,自养呼吸贡献比逐渐减小。可见,土壤含水率的变化会直接调节着土壤呼吸及其异养和自养组分的变化,进而影响着各组分的贡献率。
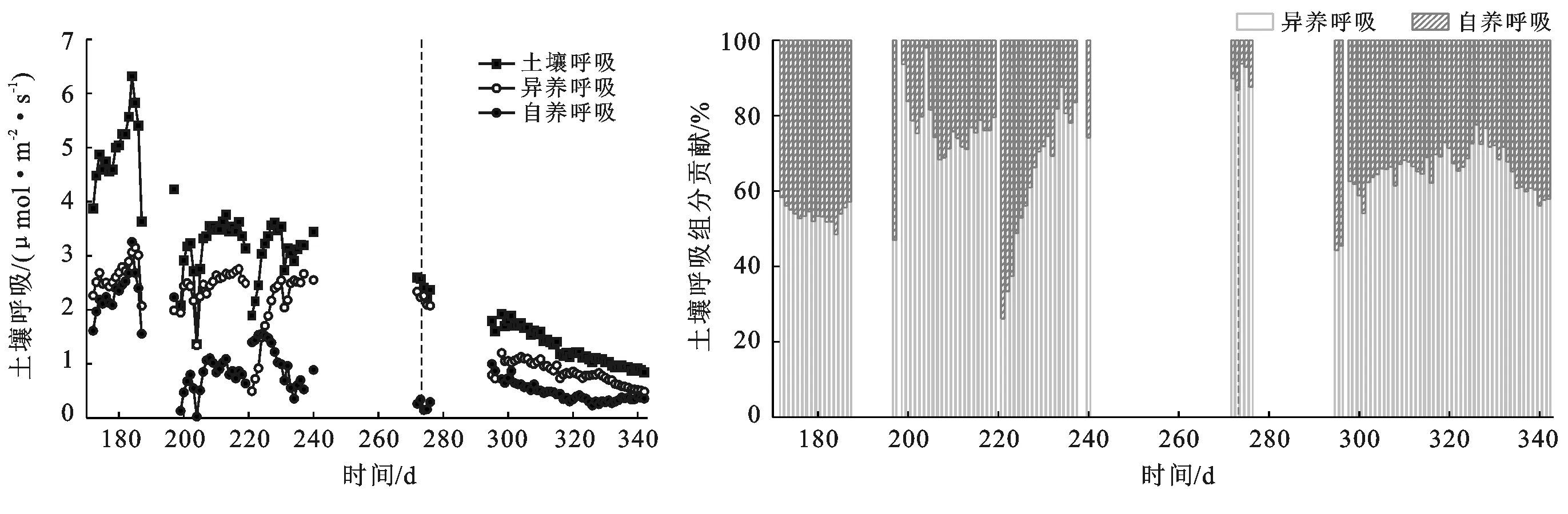
图2 土壤呼吸速率及其组分贡献率变化(生长季和非生长季以虚线为界)
和生长季的6—9月类似,在非生长季的10—12月,异养组分贡献率也高于自养,二者平均贡献率分别为67.2%和32.8%。但非生长季的10—12月二者贡献率变化基本稳定,主要与此阶段无灌溉和少降水有关,土壤含水率稳定减小,因此土壤呼吸及其各组分均呈一致减少趋势,所以异养和自养呼吸的贡献率也基本保持一致。
3.3 土壤温度对土壤呼吸和异养呼吸的影响
3.3.1 小时尺度上土壤温度对土壤呼吸、异养呼吸的影响 已有研究指出,自养呼吸部分主要受到光合产物的调节[22],且本研究区异养组分较自养组分占比高,因此我们主要探讨了土壤温度对于土壤总呼吸和异养组分的影响。生长季的6—9月和非生长季的10—12月,小时尺度上土壤温度和土壤呼吸、异养呼吸的关系均呈明显回环关系(图3)。结合土壤呼吸和土壤温度的月平均日变化(图4)来看,土壤呼吸与土壤温度的平均日变化均近似正弦函数曲线,但二者的日变化之间具有一定的时间滞后效应,因此出现了日尺度上的顺时针环状关系。其中,不论是生长季的6—9月还是非生长季的10—12月,土壤温度均从早上8:00开始增加;生长季由于日照时间长土壤温度在18:00达到最大,而土壤呼吸则于16:00达到最大;非生长季土壤温度在16:00达到最大,其土壤呼吸在12:00达到最大。

图3 土壤呼吸及其异养组分与土壤温度的关系
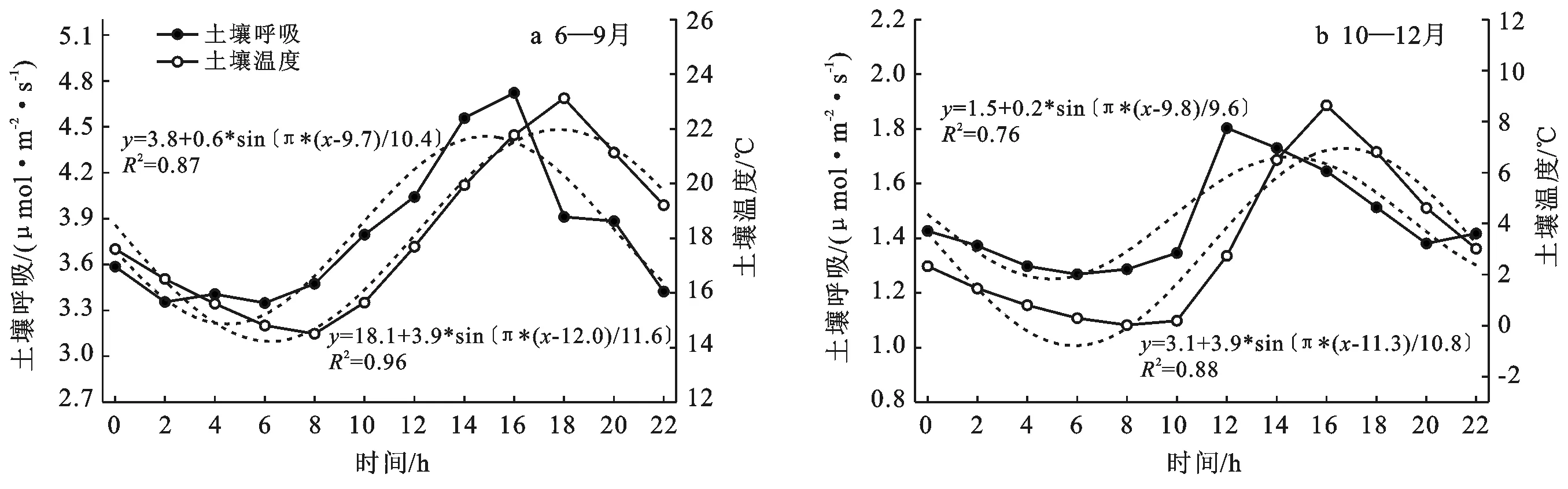
图4 土壤呼吸与土壤温度的平均日变化
3.3.2 日尺度上土壤温度对土壤呼吸、异养呼吸的影响 在日尺度上,土壤温度与土壤呼吸、异养呼吸的关系见图5。随着土壤温度的升高,土壤呼吸、异养呼吸也在增强,其中非生长季10—12月表现为明显的指数增长,且异养呼吸的土壤温度敏感性最高,Q10值为2.07。然而,生长季的6—9月几乎不存在指数关系(R2极低),通过拟合生长季3种不同水分条件下的土壤呼吸与土壤温度的关系进行验证,发现主要是因为灌溉和较大降雨引起土壤含水率波动进而干扰了生长季的土壤呼吸对于土壤温度的指数响应。在微弱降水后,土壤呼吸和异养呼吸的指数模型拟合较好,较大降水时指数模型拟合较差,而大水漫灌后则几乎不存在指数关系,甚至表现为负相关。
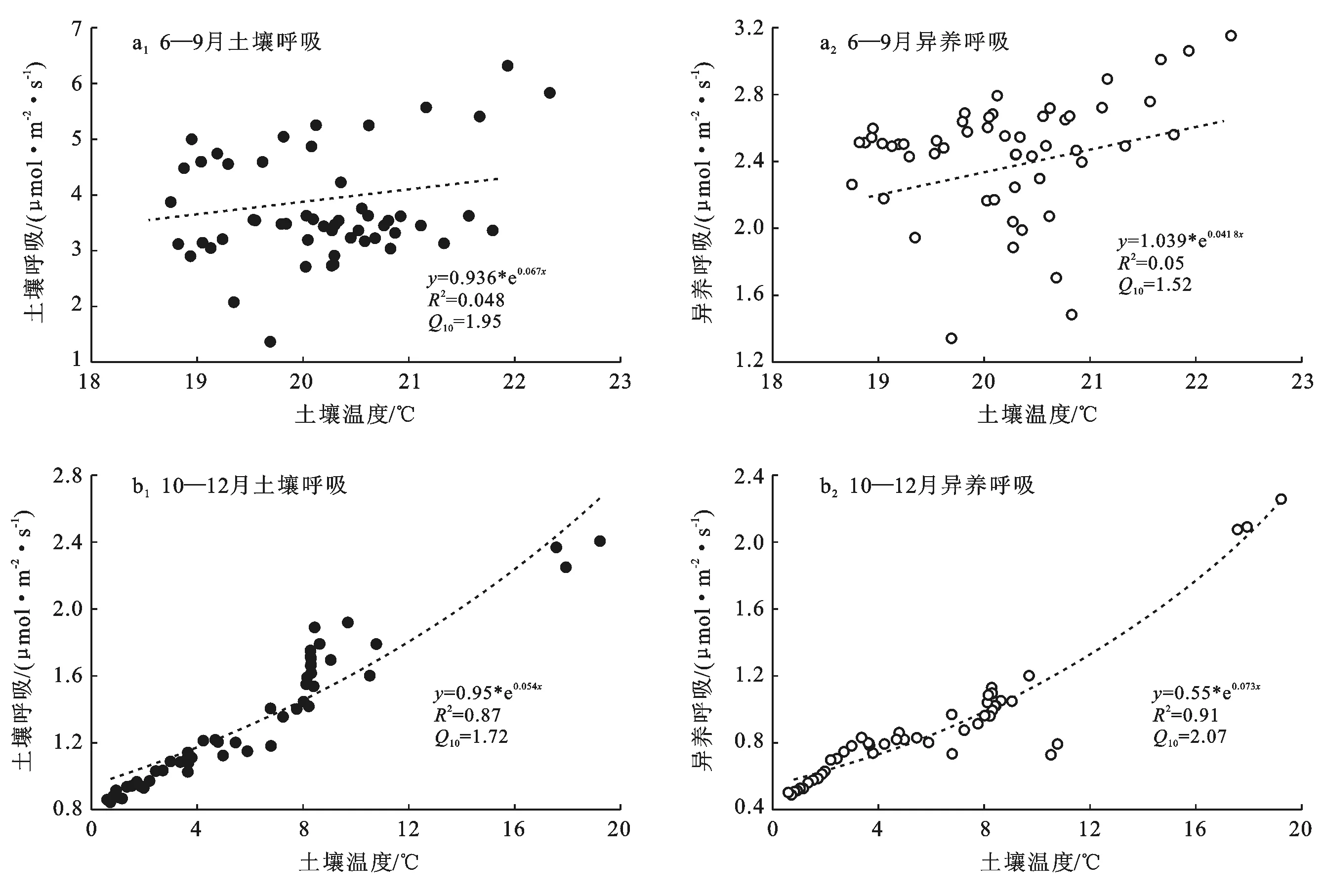
图5 土壤温度对土壤呼吸及其异养组分的影响
3.4 土壤含水率对土壤呼吸、异养呼吸和自养呼吸的影响
(1) 生长季土壤含水率对土壤呼吸及组分的影响。生长季的6—9月,3种不同初始水分条件下的土壤呼吸、异养和自养呼吸三者与土壤含水率均表现为较好的二次函数关系(图6)。其中在微弱降水、年最大降水和灌溉后的条件下,土壤呼吸均会受到立即抑制且灌溉后的抑制作用更强,其土壤呼吸和异养呼吸速率初期甚至达到1.90,0.50 μmol/(m2·s)。
在土壤呼吸、异养呼吸速率受到抑制之后,由于该区较高的辐射和强烈的蒸散发,在生长季期间,土壤含水率在微弱降水、年最大降水和灌溉之后均逐渐减少,土壤呼吸则逐渐增加且增加速率先快后慢;当土壤呼吸达到最大值时对应最适土壤含水率,不同水分条件下各组分的最适含水率存在差异(图6和表1),其中各组分的最适含水率均表现为:灌溉后>年最大降水后>微弱降水后;另外不同水分条件下的最适含水率均表现为:自养呼吸>土壤呼吸>异养呼吸。

图6 不同初始水分条件下土壤呼吸及其组分与土壤含水率的关系

表1 生长季不同水分条件下对应的土壤呼吸异养呼吸和自养呼吸的最适土壤含水率 %
土壤含水率达到最适值后,土壤呼吸又会受到干旱的抑制而降低,但由于葡萄园每月一次大水漫灌,所以生长季6—9月期间土壤含水率基本维持在8%以上,能够保证葡萄树的正常生长。
(2) 非生长季土壤含水率对土壤呼吸及组分的影响。土壤含水率在非生长季初期约为10%,大于生长季的最适含水量,随着其含水率的减少,土壤呼吸及其组分没有和生长季的变化趋势一样,而是呈指数减少趋势,减少速率先快后慢,初期减少速率较快主要是因为含水率下降的同时土壤温度也在持续降低,且葡萄埋土冬藏造成呼吸均迅速降到低值,然后处于稳定状态所致。
3.5 土壤温度和土壤含水率对土壤呼吸及其组分的综合影响
由于土壤呼吸会受到土壤温湿度的综合调控,因此本文尝试探讨在生长季和非生长季土壤温度、土壤含水率对土壤呼吸的共同影响(表2),结果表明,双因子模型可以解释生长季6—9月和非生长季10—12月土壤呼吸变化的32%和91%。

表2 土壤温度和土壤含水率对土壤呼吸组分的响应模型
同时自养呼吸和异养呼吸的生长季6—9月双因子模型拟合优度较差,而非生长季10—12月则表现良好。在非生长季的10—12月,土壤呼吸的双因子模型拟合关系较好,在三维空间上呈显著正相关变化,即土壤呼吸随着土壤温度和土壤含水率的增大而增大;而在生长季的6—9月,我们尝试了多种双因子模型,拟合优度均较低,最终确定的模型也仅能够解释土壤呼吸变化的32%,这主要与生长季土壤呼吸影响因素(生理因子和环境变量)的复杂性有关,因此仅考虑温湿度因子不能很好地反映土壤呼吸的变化。
4 讨 论
研究发现,土壤呼吸、异养呼吸对于土壤温度的日响应和季节响应特征是不同的。在小时尺度上,土壤呼吸和异养呼吸二者与5 cm土壤温度的关系均表现为明显的回环特征,这与前人研究[22-24]中森林、草地和荒漠等其他生态系统的土壤呼吸和土壤温度的回环关系相似,这种回环关系的产生主要是因为土壤呼吸和土壤温度之间存在时间滞后效应,本研究不仅验证了葡萄园生长季和非生长季土壤呼吸和土壤温度的这种回环关系,同时确定了异养组分与其具有相似性,但是有研究指出回环的变化还会受到土壤含水率等因素的影响,表现出不同的大小和倾角[25],因此后续仍需深入探究葡萄园的回环变化与内在调节机理,开展葡萄园的不同呼吸组分滞后回环变化与模型模拟研究。而在日尺度上,非生长季的10—12月份,土壤呼吸、异养呼吸与土壤温度则均符合指数关系;但生长季的6—9月,灌溉和较大的降雨都会干扰生长季的土壤呼吸对于土壤温度的这种指数响应。本研究显示生长季土壤温度对土壤呼吸的解释能力仅为4%,Q10为1.95,将整个生长季拆分后,3个时段的R2有所提升,Q10分别为2.45,3.67,0.67。针对生长季的土壤呼吸指数关系R2极低和Q10大小的问题,我们对比了其他研究发现了类似的结论[26-28]。例如,卢闯等[28]研究的夏玉米生长季指数模型R2仅为0.03,Q10为1.29,但生长季拆分的3个生育期内则指数关系均表现良好。这表明:土壤呼吸与土壤温度的指数模型在生长季会受到复杂因素的干扰,此时计算的Q10能够反映生长季土壤呼吸对于土壤温度的敏感程度,研究整个生长季和生长季期间的各个时段的Q10,对于理解这种干扰也具有重要意义。
土壤含水率作为影响土壤呼吸的另一重要因素,许多学者已指出二者的二次函数关系,在水分过高或过低时都将导致土壤呼吸速率的降低[21,29],生长季期间葡萄园也存在这种二次函数关系,且在不同水分条件下的土壤呼吸及其组分的最适含水率存在差异,土壤呼吸的最适含水率范围大约为8.1%~9.9%,但在非生长季的10—12月,土壤呼吸与土壤温度却表现为指数关系。生长季的6—9月份,各组分的最适含水率均表现为:灌溉后>年最大降水后>微弱降水后,这主要与灌溉后的呼吸脉冲响应较早有关;另外不同水分条件下的最适含水率均表现为:自养呼吸>土壤呼吸>异养呼吸,土壤呼吸阈值基本介于异养呼吸和自养呼吸之间,表明土壤呼吸是受异养组分和自养组分共同调节的,自养组分在水分的逐渐降低过程中,最先达到最适含水量,主要由于葡萄植株长期处于干旱、高蒸散发的环境下,获得水分后会迅速响应,在含水率较高时立即产生自养呼吸高值。而异养呼吸的最适含水量值则极低,一方面是因为土壤中充满水分会造成土壤氧气减少,部分好氧菌呼吸会显著减弱;另一方面土壤中水分较多,尤其是灌溉后,尽管辐射强、空气温度较高,但土壤温度会因水分的存在而较低,因此微生物活性较弱。在非生长季,土壤呼吸、异养呼吸和自养呼吸与土壤含水率均表现为指数关系,呼吸随着土壤含水率的减少而减小,速率先快后慢。结合非生长季的土壤温度来看,土壤含水率逐渐下降的同时,土壤温度也逐渐降低,因此该阶段微生物活性会迅速下降,异养呼吸减少;葡萄植株刚埋于地下,枝干与根系生理活动迅速减弱,自养呼吸也快速减小,所以初期随着土壤含水率和土壤温度的降低,土壤呼吸降低速率较大。本研究结合异养和自养组分探究了土壤含水率对于土壤呼吸的影响,然而对于各组分与土壤含水率的关系后续仍需结合微生物群落特征验证。
土壤呼吸会受到土壤温湿度的综合调控,本研究采用双因子模型拟合二者的关系,结果表明,仅考虑土壤温湿度的双因子模型可以较好的解释非生长季的土壤呼吸变化,但在生长季土壤温湿度仅能解释土壤呼吸变化的32%,这主要是由于在生长季葡萄的生理因子(物候期、冠层发育(如叶面积指数)、总初级生产力等)和其他环境因子(光合有效辐射等)与土壤呼吸密切相关。后续有待进一步建立生长季土壤呼吸的多因子模型,以便更好地模拟生长季土壤呼吸的变化。另外,研究对比了观测期间的生长季和非生长季土壤温湿度对于土壤呼吸影响存在的显著差异,但由于生长初期仪器维修等特殊情况,未能够获取2019年全序列的生长季和非生长季数据,未来应探究完整的生长季和非生长季期间的这种差异。
5 结 论
(1) 观测期内,葡萄园的土壤呼吸速率在7月3日达到最大值,为6.31 μmol/(m2·s),该区异养组分贡献率较自养组分普遍偏高,平均异养贡献率约为65%。土壤呼吸受到土壤温湿度的综合调控,生长季和非生长季的双因子模型可以解释土壤呼吸变化的32%和91%。
(2) 在小时尺度上,土壤呼吸及其异养组分与土壤温度由于日变化的时间滞后效应而均呈回环关系。而在日尺度上,非生长季的10—12月,土壤呼吸、异养呼吸随着土壤温度的增加呈指数增加趋势,且异养组分的温度敏感性更高,Q10值达到2.07;但在生长季的6—9月份,灌溉和较大降雨会引起土壤含水率波动进而干扰上述指数响应。
(3) 生长季的6—9月份,土壤呼吸及其组分与土壤含水率表现为二次函数关系,土壤呼吸的最适含水率范围约为8.1%~9.9%;而非生长季10—12月则呈指数关系,二者响应差异主要是由于非生长季的10—12月土壤含水率下降的同时土壤温度持续降低且葡萄被埋土冬藏,造成异养、自养呼吸均迅速降到低值,然后处于稳定状态所致。
