Chromosomal mapping of 5S and 18S-5.8S-25S rRNA genes in Saccharina japonica (Phaeophyceae) as visualized by dual-color fluorescence in situ hybridization*
2021-04-14YuLIUPengfeiLIUYanhuiBIZhigangZHOU
Yu LIU , Pengfei LIU, Yanhui BI , Zhigang ZHOU
1 Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources Conferred by Ministry of Education, Shanghai Ocean University, Shanghai 201306, China
2 National Demonstration Center for the Experimental Teaching of Fisheries Science, Shanghai Ocean University, Shanghai 201306, China
3 International Research Center for Marine Biosciences Conferred by Ministry of Science and Technology, Shanghai Ocean University, Shanghai 201306, China
Abstract It has been reported that there was a linkage of 5S rRNA gene to 18S-5.8S-25S rRNA gene in a few of species in Ochrophyta. In regard to the usual two positions oflinked 5S rDNA to the 3' end of 25S rDNA, two pairs of primers were designed for amplification to verify this linkage of two genes in a kelp cultivar of Saccharina japonica, one of species in Ochrophyta. This result supplemented the previous report that 5S rDNA was unlinked to 25S rDNA in this kelp. In order to simultaneously visualize this unlinkage of two genes, dual-color fluorescence in situ hybridization (FISH) technique was applied to the cytogenetics of S. japonica. Dual-color FISH images showed that two and four hybridization signals were present in the kelp gametophyte and sporophyte, respectively, metaphase nuclei hybridized simultaneously with the labeled probes of 18S rDNA and 5S rDNA. Both haploid and diploid karyotypes in decreasing length of chromosomes showed that 18S-5.8S-25S rDNA was localized at the interstitial region of Chromosome 23,whereas 5S rDNA resided at the sub-telomeric region of Chromosome 27. These karyotypes suggested that the kelp nuclear genome had only one locus of each rRNA gene, and their loci on diff erent chromosomes indicated the physical unlinkage of 5S rDNA to 18S-5.8S-25S rDNA in this kelp. Therefore, dual-color FISH seems to be a powerful technique for the discrimination and pairing of chromosomes featured in both small size and nearly identical shape in S. japonica.
Keyword: 5S rDNA; 18S-5.8S-25S rDNA; chromosome; fluorescence in situ hybridization (FISH); kelp;linkage; locus
1 INTRODUCTION
Ribosomal RNAs (rRNAs), constituting vital components of ribosomes, are coded by rRNA genes(rDNAs). Nuclear rDNAs are present in thousands of copies and organized in tandem arrays at one or more chromosomal loci (Garcia et al., 2012), and they are categorized into two diff erent families in nearly all of eukaryotes. The major rRNA family contains 18S,5.8S, and 25S/26S/28S rRNA genes that encode 45S pre-ribosomal RNA as synthesized by RNA polymerase I (Paule and White, 2000). The minor non-nucleolus forming 5S rRNA family contains a highly conserved 120-bp long coding region and a variable intergenic spacer region (Long and Dawid,1980; Volkov et al., 2007), and it is transcribed by RNA polymerase III (Paule and White, 2000). In many lower eukaryotes and most of higher eukaryotes examined, 5S rRNA genes are found usually at a diff erent locus from that of 45S rRNA genes on their nuclear genomes (Volkov et al., 2007; Garcia et al.,2017; Sochorová et al., 2018). However, physical linkage of 5S rRNA and the major rRNA family has been reported in the brown seaweed Scytosiphon lomentaria (Kawai et al., 1995) and other related taxa in Ochrophyta (or Chromophyta or Chromista or Stramenopiles) except the examined Synura(Synurophyceae) and two diatoms ( Ditylum and Nitzschia, Bacillariophyceae) (Kawai et al., 1997).This linkage has even been found in the bryophytes Marchantia and Funaria (Sone et al., 1999), the gymnosperm Ginkgo (Galián et al., 2012) and the angiosperms Artemisia (Garcia et al., 2009) and the subfamily Aurantioideae (Barros e Silva et al., 2013)in Plantae.
In the brown seaweed Saccharina japonica, one species of the ochrophytes, Yotsukura et al. (2002)indicated that its 5S rDNA could be distinguished from the 18S-5.8S-25S rDNA (i.e. 45S rDNA) after polymerase chain reaction (PCR) amplification analysis. By using mono-color fluorescence in situ hybridization (FISH) technique, Liu et al. (2017)observed that 5S rRNA gene located on Chromosome 27 and 18S-5.8S-25S rRNA gene resided on Chromosome 23 of this kelp as illustrated by independent FISH images. This finding suggested that 5S rDNA could be unlinked to 45S rDNA in S. japonica, which was in agreement with the results as examined in most of higher plants (Garcia et al.,2012, 2017). In order to concurrently visualize this unlinkage of two genes in this kelp, dual-color FISH technique was employed as suggested by Liu et al.(2017) in the present study. As a result, independent location of the 5S rDNA and 18S-5.8S-25S rDNA on diff erent chromosomes in both gametophyte and sporophyte nuclei of S. japonica clearly illustrates the unlinked relationship between these two rRNA genes.
2 MATERIAL AND METHOD
2.1 Algal material
Saccharina japonica C E Lane, C Mayes, L D Druehl & G W Saunders 2006 (= Laminaria japonica Areschoug 1851) was selected as the plant material in the present study. The gametophytes germinated from its zoospores were isolated under a microscope and cultured under the vegetative growth conditions of 30 μ mol·photons/(m2·s) and 17 °C with a photoperiod 12 h꞉12 h (light꞉dark) (Zhou and Wu, 1998). PES medium (Starr and Zeikus, 1993) was used and replaced once every two weeks.Juvenile sporophytes (about 1 cm in size),developed from the isolated male and female gametophytes of this kelp, were presented by Oriental Ocean Sci-tec Co., Ltd., Yantai, Shandong Province,China.
2.2 DNA isolation
Genomic DNA was extracted from freshly harvested gametophytes according to the modified cetyltrimethyl ammonium bromide (CTAB) method of Hu and Zhou (2001). The extracted DNA was dissolved in Tris-EDTA buff er which was composed of 10 mmol/L Tris-HCl and 1 mmol/L EDTA at pH 8.0.
2.3 PCR
To ascertain whether 5S rDNA is linked to the 18S-5.8S-25S rDNA unit, two pairs of primers were designed based the coding sequences of 25S rDNA(GenBank Accession No. KX827269) and 5S rDNA(Yotsukura et al., 2002). They were 25S-F(5'-TACTGGGGGAATCCTGGTTAGTTT-3') and 5S-RF (5'-CGTGGTCCCCCACCGTAGTACTAGCCT-3'), and 25S-F and 5S-R (5'-AGGCTAGTACTACGGTGGGGGACCACG-3'). Amplification was conducted in a 25- μ L reaction volume containing the template DNA (50 ng), Mg2+(2.5 mmol/L), Taq DNA polymerase (1 U), dNTPs (100 μ mol/L each),forward and reverse primers (0.2 μ mol/L each), and 1× reaction buff er. PCR was performed in a gradient Mastercycler (Eppendorf, Hamburg, Germany) and programmed as described by Hu and Zhou (2001)except for the use of 30 cycles of the reaction and annealing at 62.8 °C for 30 s. The amplified products were resolved on a 1.0% low-melting-point agarose gel for DNA recovery. The target product was purified using a DNA gel extraction kit (Aidlab, Beijing,China) and ligated into a pMD19-T vector (TaKaRa,Dalian, China). The constructed vector was subsequently transformed into Escherichia coli DH5 α competent cells (Tiangen, Beijing, China), and several positive clones were screened and sent to Sangon Biotech (Shanghai, China) for sequencing.
2.4 Labeling of 18S and 5S rDNA probes
Prior to FISH, the unlabeled 18S and 5S rDNA probes were generated by PCR with the gametophyte genomic DNA as a template. PCR was carried out using the pair of primers 18S-F (5'-ACCTGGTTGATCCTGCCAGT-3')/18S-R(5'-TCACCTACGGAAACCTTGT-3') and 5Ss-F (5'-TATACATGCGCGCCAACTTG-3')/5Ss-R(5'-CCCCGTGGGTACGAGTAACA-3') as used by Liu et al. (2017). PCR was programmed as described by Liu et al. (2017) and the amplified products were recovered, purified, and ligated into plastid pMD19-T as described above. The 18S rDNA probe was labeled with Texas Red-5-dCTP(Invitrogen, Carlsbad, CA, USA), whereas the 5S rDNA probe with Alex Flour Green-5-dUTP(PerkinElmer, Boston, MA, USA), by nick translation(Rigby et al., 1977) using an ADVANCETMNick Translation Kit (Sigma-Aldrich, St. Louis, MO, USA)under the guidance of the manufacturer’s instructions.Twenty μ L reaction contains independent plastid DNA (200 ng/ μ L) 10 μ L, 10×Nick Translation buff er 2 μ L, labeled dNTP (1 mmol/L) 0.5 μ L, non-labeled dNTP (2 mmol/L each mixed) 2 μ L, DNA polymerase I (10 U/ μ L) 4 μ L, DNase I (100 U/mL) 0.4 μ L. The labeled probes were checked by electrophoresis and stored at -20 °C.
2.5 Chromosome preparation
The freshly harvested gametophytes and sporophyte tissues of S. japonica were independently treated with 0.02% colchicine for 8- 10 h at room temperature and then washed 3 times in sterilized seawater. The samples were subsequently fixed in a freshly prepared Carnoy’s fixative solution (100%ethanol꞉acetic acid, 3꞉1, v/v) for 24 h (Schweizer,1976). After enzymatic digestion with mixture of cellulose, pectinase, cellulose, macerozyme R-10,and extracted abalone alginate lyase as described by Liu et al. (2012), the digested samples were dropped on slides for chromosome preparation. The slides were screened under a phase contrast light microscope to detect whether the chromosomes were at mitotic metaphase. The well-prepared slides were subjected to FISH detection.
2.6 Dual-color FISH
The FISH experiment was conducted as described by Liu et al. (2012) with slight modifications. The chromosomes on slides were ultraviolet (UV) light cross-linked twice in a UV cross-linker BLX-E254(Vilbert Lourmat, France) at 254 nm for 15 s each time, followed by addition of 8 μ L of (0.9 mol/L NaCl, 20 mmol/L Tris-HCl, pH 7.2, 0.01% SDS)containing 5 ng/ μ L of the labeled two probes. The slides were heated to 100 °C in a water bath for 5 min and then transferred to a 55 °C air oven for 12 h for hybridization to occur. Post-hybridization washes of the slides were carried out in a slightly more stringent solution of 2×SSC (0.3 mol/L NaCl, 0.03 mol/L sodium citrate, pH 7) at 42 °C for 10 min to remove unbound, nonspecifically bound, or weakly hybridized probes.
The chromosomes were counterstained with 15 μ L 4',6-diamidino-2-phenylindole (DAPI, 2 μ g/mL) in a VectaShield mounting medium (Vector Laboratories,Burlingame, CA, USA) for 5 min incubation(Schweizer, 1976). The slides were stored in the dark before microscopic analysis.
2.7 Fluorescence microscopy and image acquisition
Images were taken with a Leica DM4000 epifluorescent microscope equipped with diff erent filter set and a digital camera Orca ER (Wetzlar,Germany). Three kinds of fluorescent signals, Alex Flour (green), Texas Red (red) and DAPI (blue), were detected at the emission wavelength of 520 nm,612 nm, and 488 nm, respectively. All images were processed using Adobe Photoshop 7.0 (http://www.adobe.com/products/photoshop.html) for color contrast and brightness uniformity. The karyotypic patterns of the gametophytes and sporophytes were edited using this software based on the chromosome size and distribution of 5S and 18S rDNA on chromosomes.
3 RESULT AND DISCUSSION
In most eukaryotes, the 5S rRNA genes are physically separated from the 18S-5.8S-25S rRNA genes, as reviewed by Garcia et al. (2017) and Sochorová et al. (2018) based on the updated information in the Plant and Animal, respectively,rDNA database (http://www.plantrdnadatabase.com/and http://www.animalrdnadatabase.com/). However,several eukaryotic species of plants including several brown macroalgae have been reported to possess 5S rDNA linked to the 3' end of 25S rDNA (Kawai et al.,1995; 1997; Sone et al., 1999; Garcia et al., 2009;Galián et al., 2012; Barros e Silva et al., 2013; Garcia and Kovařík, 2013). As proposed by Drouin and de Sá(1995), the 5S rRNA genes could be transcribed on the same or opposite strand as compared to the genes to which they were linked (Supplementary Fig.S1a).Accordingly, we designed two pairs of primers (25SF and 5S-RF, 25S-F and 5S-R) for the amplification using PCR to investigate a kelp cultivar of S. japonica for the linkage of 5S rDNA to 25S rDNA. The electrophoresis profile (Supplementary Fig.S1b) of these amplified products illustrated that several fragments were obtained. Upon cloning and sequencing, it was found that they were the results of nonspecific amplification. This result suggested that the kelp 5S rDNA could not be localized downstream from the 3' end of 25S rDNA regardless of the transcription orientation. This supplemented the previous results also by using PCR but only concerning one direction of transcription for the design of primers in this kelp (Yotsukura et al., 2002).
To simultaneously visualize this unlinked relationship of two genes, dual-color FISH technique was employed as suggested by Liu et al. (2017). With the plastid pMD19-T containing kelp 18S rDNA or 5S rDNA fragments (Supplementary Fig.S1) as templates, both of 18S rDNA and 5S rDNA probes were labeled by nick translation (Rigby et al., 1977).The labeled probes appeared smear with a peak intensity of about 300 bp as resolved by electrophoresis (Supplementary Fig.S2), suggesting that they were suitable for the requirements of next experiment (Zhang and Friebe, 2009). After attempting to perform independently with single labeled probes as advised by Jowett (1999), the ratio of 1꞉1.5 (18S rDNA꞉5S rDNA, v/v) in the mixed hybridization buff er was determined. More than 100 metaphase nuclei of the S. japonica gametophytes were examined, and 67.9% (93/137) of them showed the presence of two distinguishable fluorescent signals on diff erent chromosomes as illustrated by the merged image (Fig.1d). The arrayed chromosomes(Fig.1f) by their decreasing length indicated that 18S rDNA (red signal) was situated on Chromosome 23 while 5S rDNA (green signal) was on Chromosome 27. This result was in agreement with the observation by Liu et al. (2017) using digoxigenin-labeled 5S rDNA or 18S rDNA probes. Apparently, the simultaneous hybridization for two targets with dualcolor labeled probes on the same chromosome slide in the present study made the visualization more convenient and intuitive than that with mono-color probes separately. Independent location of 5S rDNA and 18S-5.8S-25S rDNA on diff erent chromosomes(Fig.1) as well as PCR amplification analysis(Supplementary Fig.S1) led to the conclusion that the 5S rRNA gene was unlinked to the 18S-5.8S-25S rRNA one in the gametophyte genomes of S.japonica.
As we know, the microscopic gametophytes are haploid whereas the macroscopic sporophytes are diploid in S. japonica (Bi and Zhou, 2014). The occurrence of two hybridization signals in the kelp gametophyte genomes thus means that four signals are expected to be present in the sporophyte ones.This was supported by the dual-color FISH image of hybridization on a sporophyte metaphase nucleus(Fig.2a) which possessed four distinguishable fluorescent signals on diff erent chromosomes. Upon chromosome pairing and arranging in the decreasing length, it was found that 18S rDNA and 5S rDNA were located on the chromosomes as same as their order in a gametophyte nuclei (Fig.2c vs. Fig.1f). So the chromosomes which rDNA resided on are easily distinguished from the others and paired for homologous chromosomes in a diploid nucleus, when the dual-color FISH technique is employed in the kelp cytogenetics.
From the exemplified FISH images (Figs.1 & 2)with dual-color labeled probes, kelp was inferred to have a single locus of both 5S and 18S-5.8S-25S rRNA genes. The latter one, usually associated with nucleolus organizer region (Volkov et al., 2007),implied that the kelp nucleus could have one nucleolus which was examined in detail under an electron microscope by Bouck (1965) and Dai and Fang(1979). The rDNA locus number in the kelp was ofinterest coincident with the statistical result that 51.38% of diploid karyotypes had a single 5S rDNA locus and 35.16% had a single 18S-5.8S-25S rDNA locus from the publications on the rDNA of higher plants (Garcia et al., 2017). Garcia et al. (2017) also stated that 76.21% of plants with a single 18S-5.8S-25S rDNA locus had that locus at a terminal location.In contrast to this, the position of kelp 18S-5.8S-25S rDNA locus was interstitial while the site of 5S one was sub-terminal as shown in Figs.1 & 2. This distribution and position of rDNA in the kelp genome as illustrated by dual-color FISH was in agreement with that by mono-color FISH (Liu et al., 2017),indicating the practicability and reproducibility of FISH technique employed in S. japonica.
4 DATA AVAILABILITY STATEMENT
All data generated and/or analyzed during this study are included in this published article and its supplementary information files.
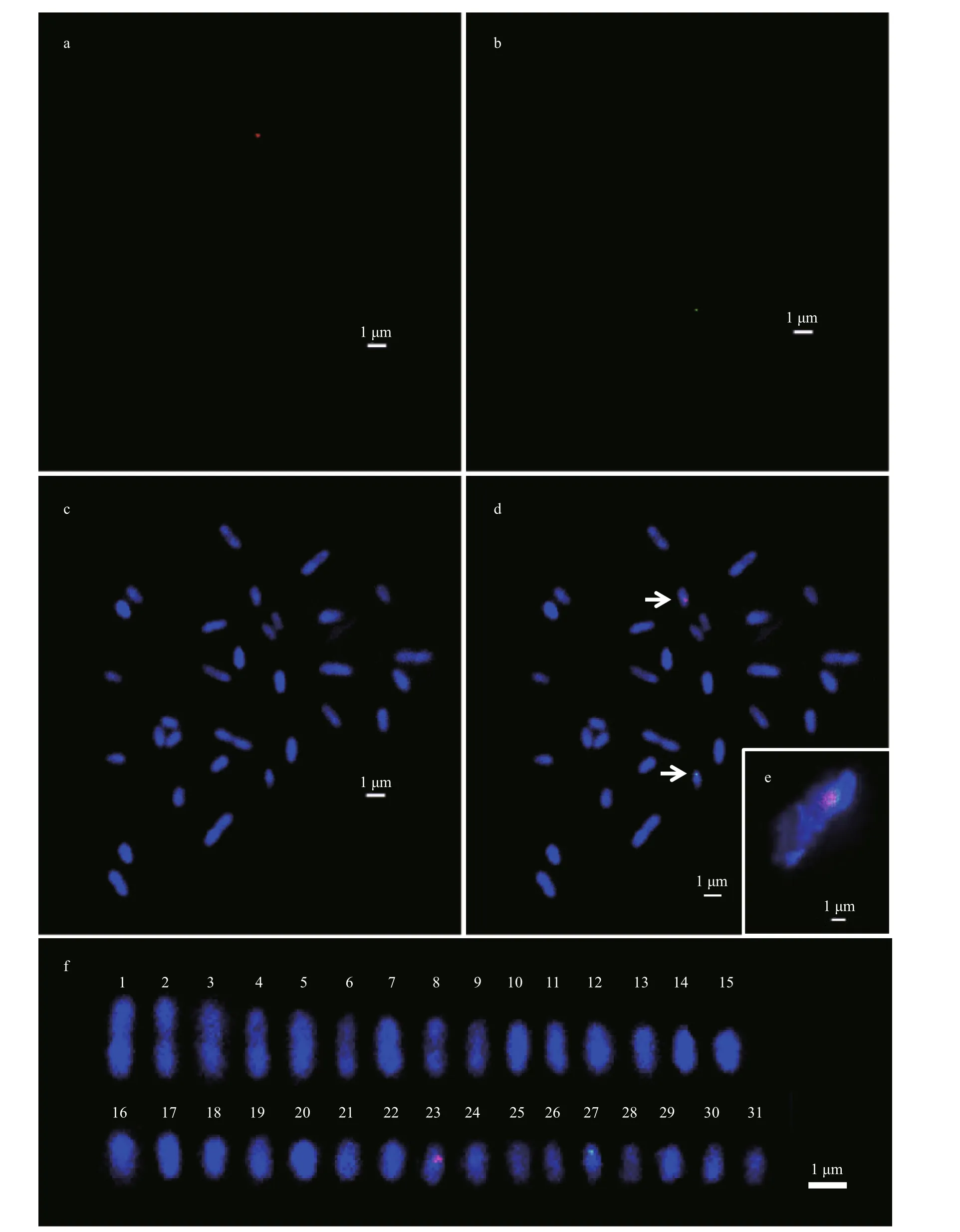
Fig.1 Dual-color fluorescence in situ hybridization of 5S (green) and 18S (red) rDNA probes on the Saccharina japonica gametophyte chromosomes counterstained with DAPI (blue)
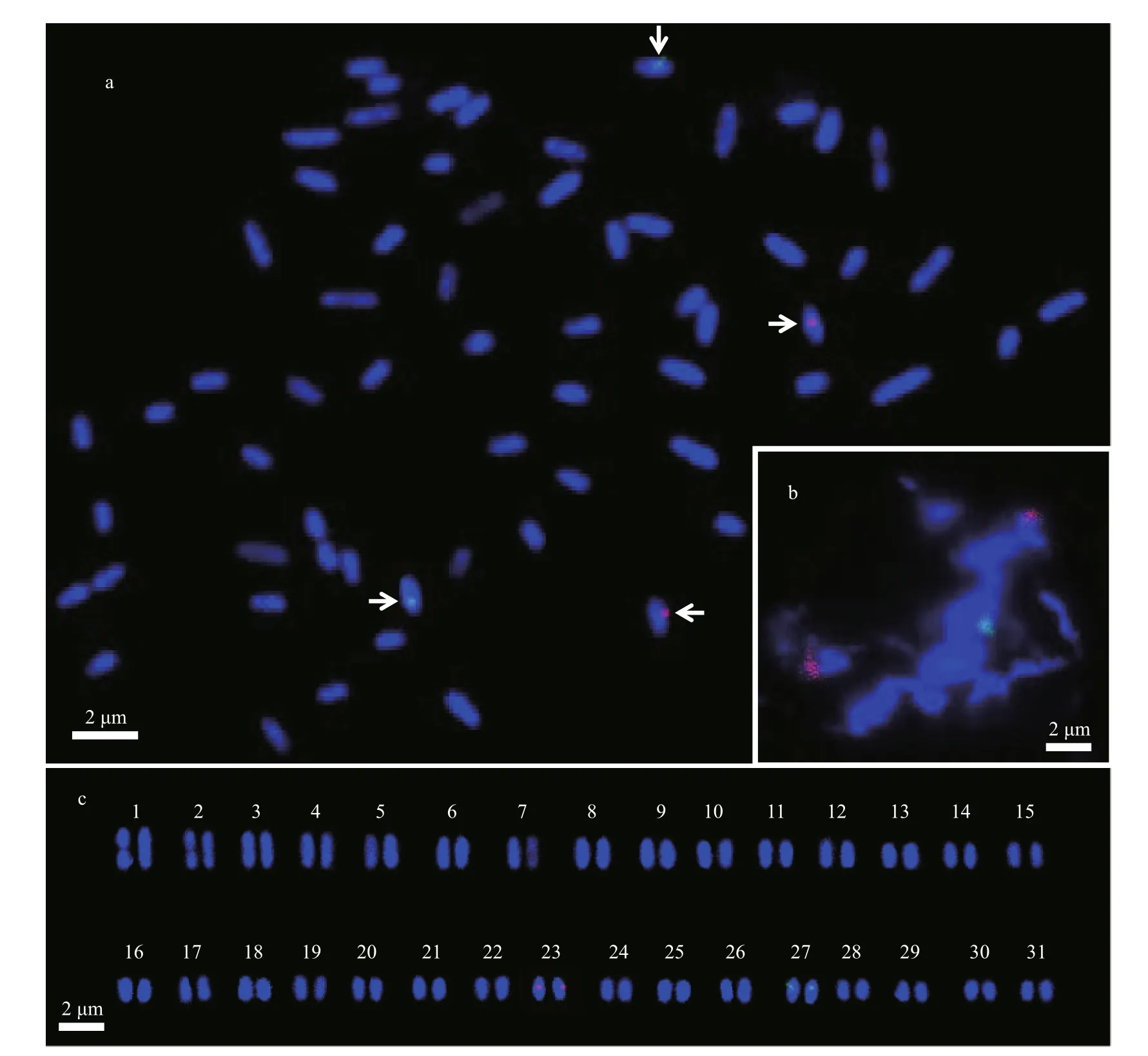
Fig.2 Dual-color fluorescence in situ hybridization of 5S (green) and 18S (red) rDNA probes on the Saccharina japonica sporophyte chromosomes counterstained with DAPI (blue)
杂志排行

Journal of Oceanology and Limnology的其它文章
- Predicting sediment flux from continental shelfislands,southeastern China*
- Laboratory simulation of dissolved oxygen reduction and ammonia nitrogen generation in the decay stage of harmful algae bloom*
- Development of high-resolution chloroplast markers for intraspecific phylogeographic studies of Phaeocystis globosa*
- Effects ofiron and humic acid on competition between Microcystis aeruginosa and Scenedesmus obliquus revealed by HPLC analysis of pigments*
- Effect of river plume on phytoplankton community structure in Zhujiang River estuary*
- Exploring the sublethal genotoxic effects of class II organophosphorus insecticide quinalphos on freshwater fish Cyprinus carpio